
Weed Species from Tea Gardens as a Source of Novel Aluminum Hyperaccumulators
 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Results
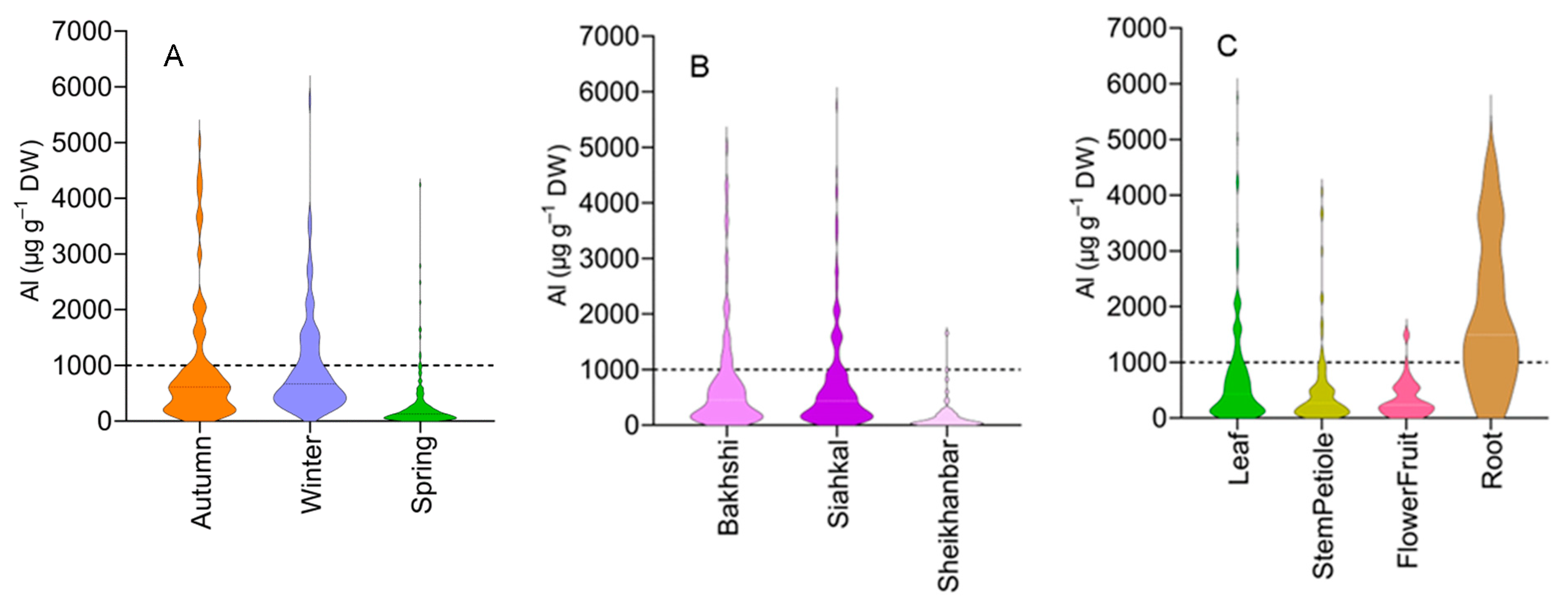
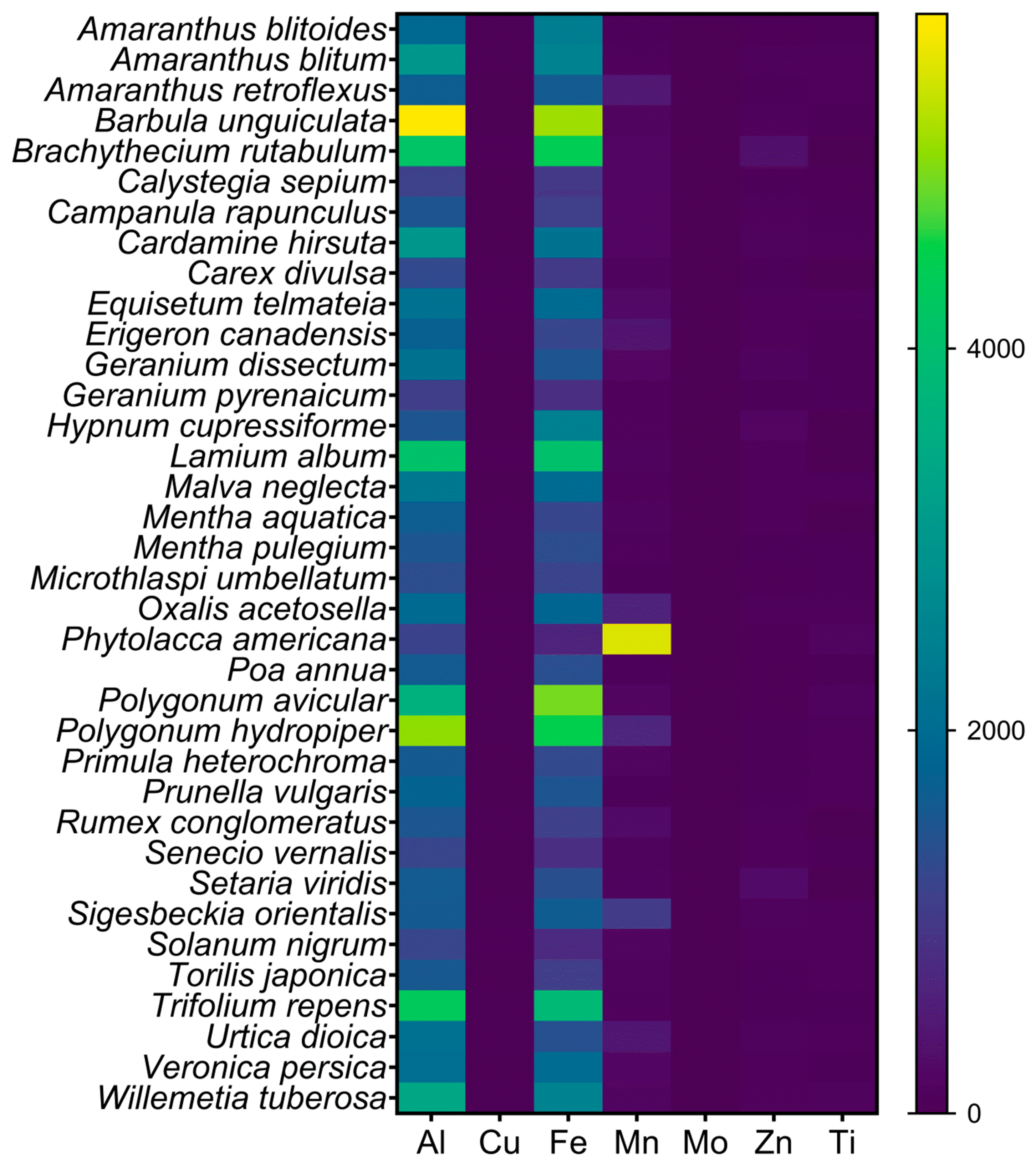
2.1. Al Concentration in the Specimens Collected from Acid Soils
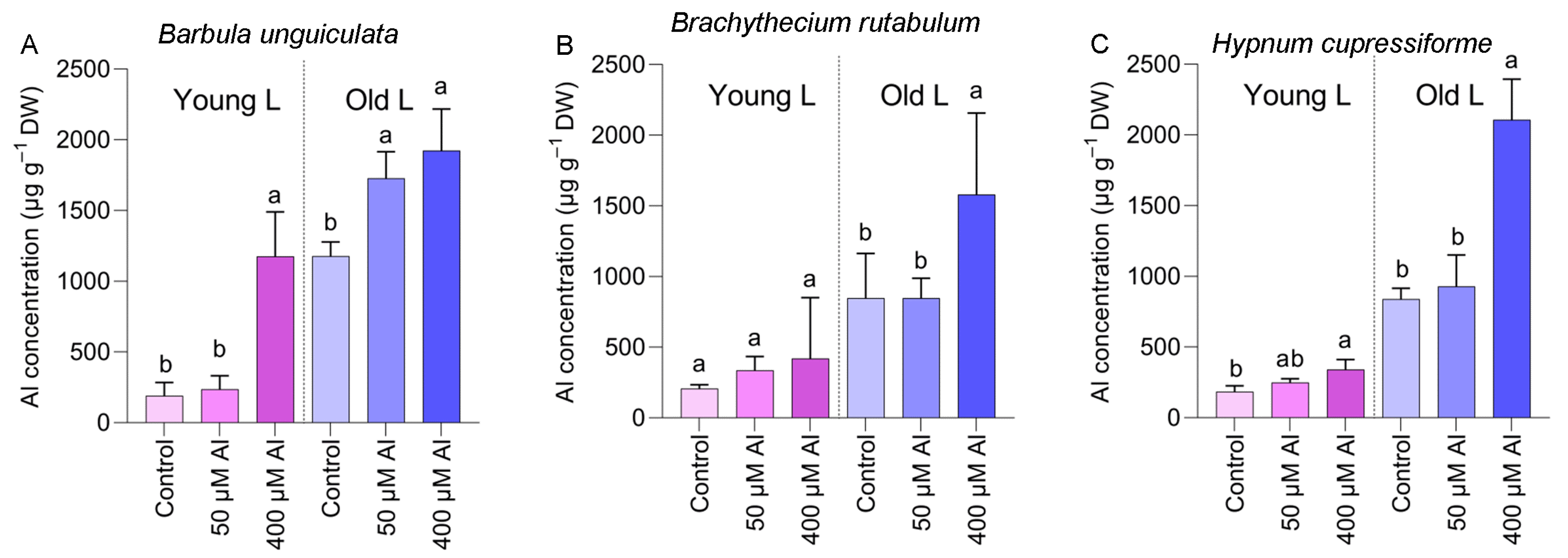
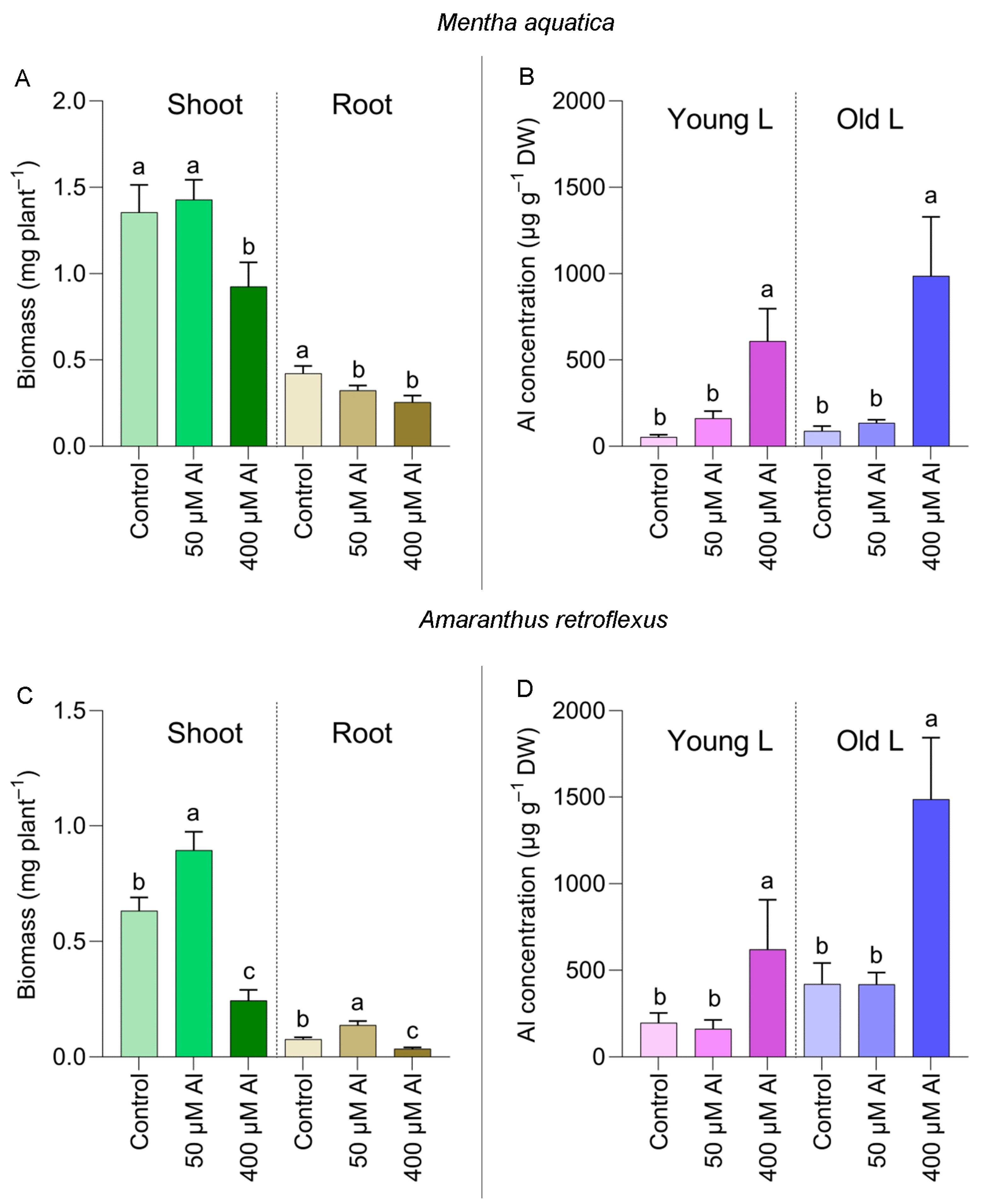
2.2. Accumulation of Al in Plants under Laboratory Conditions
3. Discussion
3.1. Al Accumulation in Species Not Previously Recorded as Al Accumulators
3.2. Accumulation of Al in Different Seasons and Locations
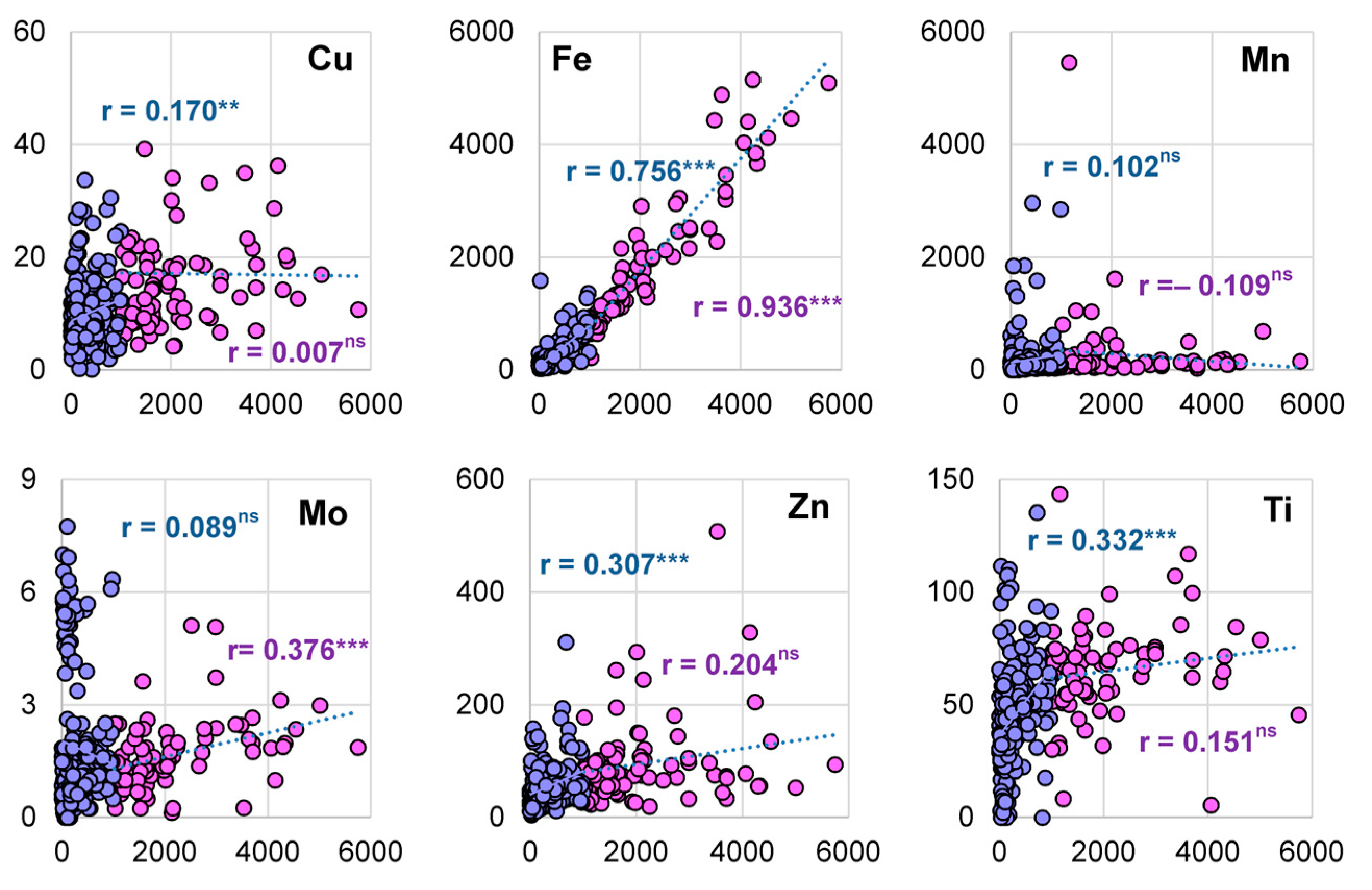
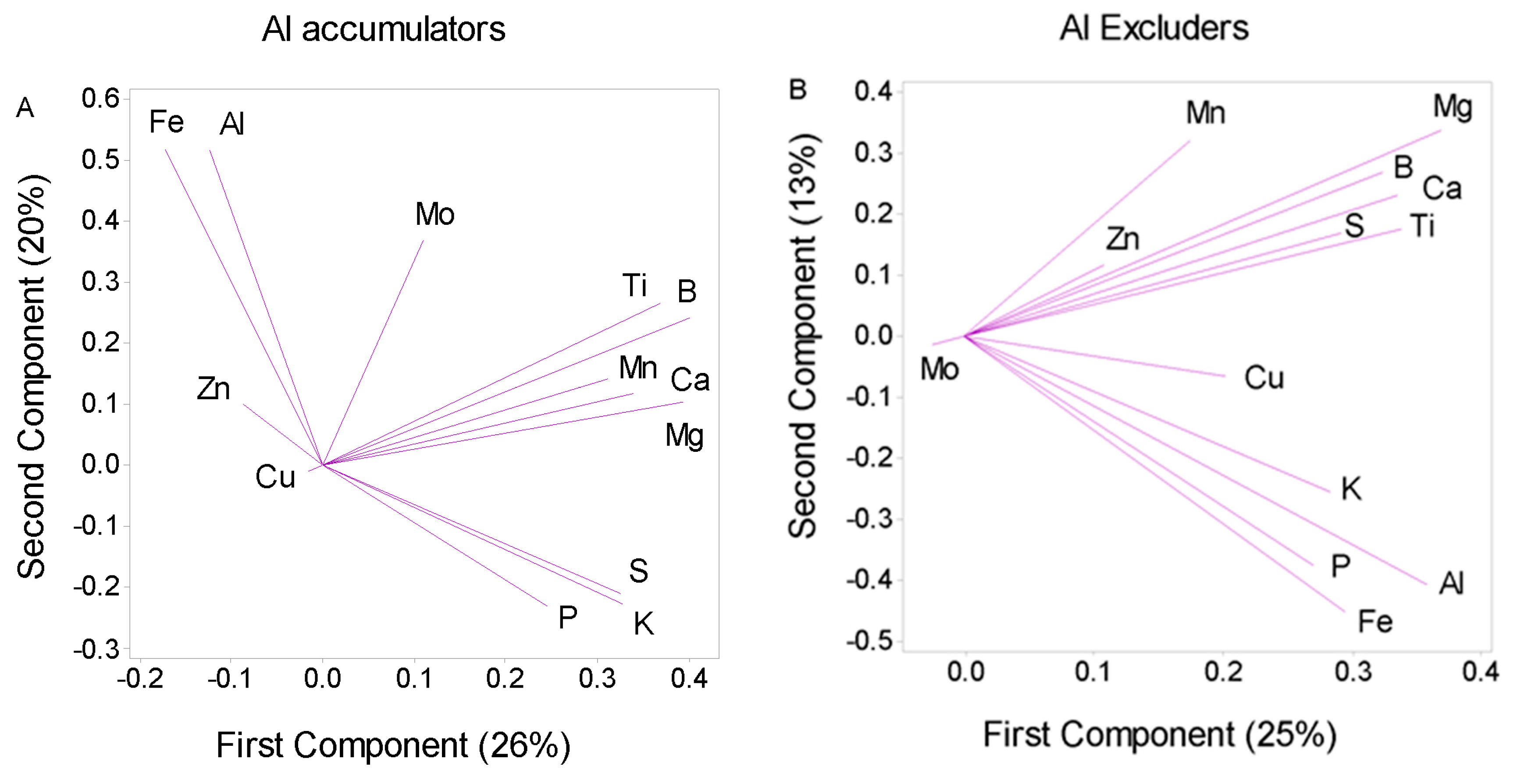
3.3. Accumulation of Al and Concentration of Micronutrients
3.4. Accumulation of Al from an Ecological and Evolutionary Point of View
4. Materials and Methods
4.1. Study Area
4.2. Collection of Soil Samples
4.3. Collection of Plants and Determination of Species
4.4. Preparation of Samples and Elemental Analyses
4.5. Plants Cultivation and Treatment under Growth Chamber Conditions
4.6. Data Analysis of Field Samples and Design of the Lab Experiments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Von Uexküll, H.R.; Mutert, E. Global extent, development and economic impact of acid soils. Plant Soil 1995, 171, 1–5. [Google Scholar] [CrossRef]
- Hajiboland, R.; Panda, C.K.; Lastochkina, O.; Gavassil, M.A.; Habermann, G.; Pereira, J.F. Aluminum toxicity in plants: Present and future. J. Plant Growth Regul. 2022. [Google Scholar] [CrossRef]
- Haynes, R.J. Soil acidification induced by leguminous crops. Grass Forage Sci. 1983, 38, 1–11. [Google Scholar] [CrossRef]
- Wei, Y.; Han, R.; Xie, Y.; Jiang, C.; Yu, Y. Recent advances in understanding mechanisms of plant tolerance and response to aluminum toxicity. Sustainability 2021, 13, 1782. [Google Scholar] [CrossRef]
- Kochian, L.V.; Pineros, M.A.; Hoekenga, O.A. The physiology, genetics and molecular biology of plant aluminum resistance and toxicity. Plant Soil 2005, 274, 175–195. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Tolrà, R.; Hajiboland, R.; Arroyave, C.; Barceló, J. Mechanisms of hyper-resistance and hyper-tolerance to aluminum in plants. In Aluminum Stress Adaptation in Plants; Panda, S., Baluška, F., Eds.; Springer: Chem, Switzerland, 2015; pp. 81–98. ISBN 978-3-319-36331-8. [Google Scholar]
- Poschenrieder, C.; Gunsé, B.; Corrales, I.; Barceló, J. A glance into aluminum toxicity and resistance in plants. Sci. Total Environ. 2008, 400, 356–368. [Google Scholar] [CrossRef]
- Liu, J.; Piñeros, M.A.; Kochian, L.V. The role of aluminum sensing and signaling in plant aluminum resistance. J. Integr. Plant Biol. 2014, 56, 221–230. [Google Scholar] [CrossRef]
- Jansen, S.; Broadley, M.R.; Robbrecht, E.; Smets, E. Aluminum hyperaccumulation in angiosperms: A review of its phylogenetic significance. Bot. Rev. 2002, 68, 235–269. [Google Scholar] [CrossRef]
- Brunner, I.; Sperisen, C. Aluminum exclusion and aluminum tolerance in woody plants. Front. Plant Sci. 2013, 4, 172. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, M.; Mehltreter, K.; Sundue, M.; Testo, W.; Watanabe, T.; Jansen, S. The evolution of aluminum accumulation in ferns and lycophytes. Amer. J. Bot. 2017, 104, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Shen, R.; Ma, J.F.; Kyo, M.; Iwashita, T. Compartmentation of aluminium in leaves of an Al-accumulator Fagopyrum escuentum Moench. Planta 2001, 215, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Branquinho, C.; Serrano, H.C.; Pinto, M.J.; Martins-Louçao, M.A. Revisiting the plant hyperaccumulation criteria to rare plants and earth abundant elements. Environ. Pollut. 2007, 146, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Agegnehu, G.; Amede, T.; Erkossa, T.; Yirga, C.; Henry, C.; Tyler, R.; Nosworthy, M.G.; Beyene, S.; Sileshi, G.W. Extent and management of acid soils for sustainable crop production system in the tropical agroecosystems: A review. Acta Agric. Scand. Section B—Soil Plant Sci. 2021, 71, 852–869. [Google Scholar] [CrossRef]
- Liu, C.A.; Liang, M.Y.; Nie, Y.; Tang, J.W.; Siddique, K.H.M. The conversion of tropical forest to rubber plantations accelerates soil acidification ad changes the distribution of soil metal ions in topsoil layers. Sci. Total Environ. 2019, 696, 13082. [Google Scholar] [CrossRef]
- Dviwedi, R.S. Geospatial Technologies for Land Degradation Assessment and Management; CRC Press: Boca Raton, FL, USA, 2018; ISBN 9780367570897. [Google Scholar]
- Yan, P.; Wu, L.; Wang, D.; Fu, J.; Shen, C.; Li, X.; Zhang, L.; Zhang, L.; Fan, L.; Wenyan, H. Soil acidification in Chinese tea plantations. Sci. Total Environ. 2020, 715, 136963. [Google Scholar] [CrossRef]
- Jahed, R.R.; Hosseini, S.M.; Kooch, Y. The effect of natural and planted forest stands on soil fertility in the Hyrcanian region, Iran. Biodiversitas J. Biol. Divers. 2014, 15, 206–214. [Google Scholar] [CrossRef]
- Box, E.O. Modeling ecological structure and function. In Macroclimate and Plant Forms. Tasks for Vegetation Science, Vol 1; Springer: Dordrecht, The Netherlands, 1981; pp. 5–9. ISBN 978-94-009-8682-4. [Google Scholar]
- Ghorbanalizadeh, A.; Akhani, H. Plant diversity of Hyrcanian relict forests: An annotated checklist, chorology and threat categories of endemic and near endemic vascular plant species. Plant Divers. 2022, 44, 39–69. [Google Scholar] [CrossRef]
- Akhani, H.; Djamali, M.; Ghorbanalizadeh, A.; Ramezani, E. Plant biodiversity of Hyrcanian relict forests, N Iran: An overview of the flora, vegetation, palaeoecology and conservation. Pak. J. Bot. 2010, 42, 231–258. [Google Scholar]
- Jansen, S.; Watanabe, T.; Smets, E. Aluminium accumulation in leaves of 127 species in Melastomataceae, with comments on the order Myrtales. Ann. Bot. 2002, 90, 53–64. [Google Scholar] [CrossRef] [Green Version]
- Römheld, V.; Nikolic, M. Iron. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; Taylor Francis Group: Boca Raton, FL, USA, 2007; pp. 329–350. ISBN 0-8247-5904-4. [Google Scholar]
- Hajiboland, R.; Barceló, J.; Poschenrieder, C.; Tolrà, R. Amelioration of iron toxicity: A mechanism for aluminum-induced growth stimulation in tea plants. J Inorg. Biochem. 2013, 128, 183–187. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Function of nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 191–248. ISBN 978-0-12-384905-2. [Google Scholar]
- Nguyen, N.T.; Hiep, N.D.; Fujita, K. Iron enhances aluminum-induced leaf necrosis and plant growth inhibition in Eucalyptus camaldulensis. Plant Soil 2005, 277, 139–152. [Google Scholar] [CrossRef]
- Humphries, J.M.; Stangoulis, J.C.R.; Graham, R.D. Manganese. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; Taylor Francis Group: Boca Raton, FL, USA, 2007; pp. 351–374. ISBN 0-8247-5904-4. [Google Scholar]
- Hamlin, R.L. Molybdenum. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; Taylor Francis Group: Boca Raton, FL, USA, 2007; pp. 375–394. ISBN 0-8247-5904-4. [Google Scholar]
- Vázquez, M.D.; Barceló, J.; Poschenrieder, C.H.; Madico, J.; Hatton, P.; Baker, A.J.; Cope, G.H. Localization of zinc and cadmium in Thlaspi caerulescens (Brassicaceae), a metallophyte that can hyperaccumulate both metals. J. Plant Physiol. 1992, 140, 350–355. [Google Scholar] [CrossRef]
- Cosio, C.; Martinoia, E.; Keller, C. Hyperaccumulation of cadmium and zinc in Thlaspi caerulescens and Arabidopsis halleri at the leaf cellular level. Plant Physiol. 2004, 134, 716–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broadley, M.R.; Willey, N.J.; Wilkins, J.C.; Baker, A.J.; Mead, A.; White, P.J. Phylogenetic variation in heavy metal accumulation in angiosperms. New Phytol. 2001, 152, 9–27. [Google Scholar] [CrossRef] [PubMed]
- Serrano, H.C.; Cotrim, H.; Pinto, M.J.; Martins-Loução, M.A.; Branquinho, C. Metal hyperaccumulation patterns within Plantago phylogeny (Plantaginaceae). Plant Soil 2017, 411, 227–241. [Google Scholar] [CrossRef]
- van der Ent, A.; Baker, A.J.; Reeves, R.D.; Pollard, A.J.; Schat, H. Commentary: Toward a more physiologically and evolutionarily relevant definition of metal hyperaccumulation in plants. Front. Plant Sci. 2015, 6, 554. [Google Scholar] [CrossRef] [Green Version]
- Goolsby, E.W.; Mason, C.M. Response: Commentary: Toward a more physiologically and evolutionarily relevant definition of metal hyperaccumulation in plants. Front. Plant Sci. 2016, 6, 1252. [Google Scholar] [CrossRef] [Green Version]
- van der Ent, A.; Baker, A.J.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Cronquist, A. Chemistry in Plant Taxonomy: An Assessment of Where We Stand; Academic Press: London, UK, 1980. [Google Scholar]
- Goolsby, E.W.; Mason, C.M. Toward a more physiologically and evolutionarily relevant definition of metal hyperaccumulation in plants. Front. Plant Sci. 2015, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Reeves, R.D.; Baker, A.J.M. Metal-accumulating plants. In Phytoremediation of Toxic Metals; Raskin, I., Ensley, B.D., Eds.; John Wiley: New York, NY, USA, 2000; pp. 193–229. ISBN 10: 0-47119-254-6. [Google Scholar]
- Djamali, M.; Akhani, H.; Khoshravesh, R.; Andrieu-Ponel, V.; Ponel, P.; Brewer, S. Application of the global bioclimatic classification to Iran: Implications for understanding the modern vegetation and biogeography. Ecol. Mediterr. 2011, 37, 91–114. [Google Scholar] [CrossRef]
- Khormali, F.; Ayoubi, S.; Kananro-Foomani, F.; Fatemi, A. Tea yield and soil properties as affected by slope position and aspect in Lahijan area, Iran. Int. J. Plant Prod. 2007, 1, 99–111. [Google Scholar]
- Rechinger, K.H. (Ed.) Flora Iranica; Naturhistorisches Museum Wien: Wien, Austria, 1963–2010. [Google Scholar]
- Assadi, M.; Massoumi, A.A.; Khatamsaz, M.; Mozaffrian, V. (Eds.) Flora of Iran; Research Institute of Forests and Rangelands Publication: Tehran, Iran, 1988–2011. (In Persian) [Google Scholar]
- Cook, L.L.; McGonigle, T.P.; Inouye, R.S. Titanium as an indicator of residual soil on arid-land plants. J. Environ. Qual. 2009, 38, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Reeves, R.D.; Baker, A.J.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Del Río-Celestino, M.; Font, R.; Moreno-Rojas, R.; De Haro-Bailón, A. Uptake of lead and zinc by wild plants growing on contaminated soils. Ind. Crops Prod. 2006, 24, 230–237. [Google Scholar] [CrossRef]
- Watanabe, T.; Murata, Y.; Osaki, M. Amaranthus tricolor has the potential for phytoremediation of cadmium-contaminated soils. Commun. Soil Sci. Plant Anal. 2009, 40, 3158–3169. [Google Scholar] [CrossRef]
- Printarakul, N.; Meeinkuirt, W. The bryophyte community as bioindicator of heavy metals in a waterfall outflow. Sci. Rep. 2022, 12, 1–2. [Google Scholar]
- Sun, Z.; Chen, J.; Wang, X.; Lv, C. Heavy metal accumulation in native plants at a metallurgy waste site in rural areas of Northern China. Ecol. Eng. 2016, 86, 60–68. [Google Scholar] [CrossRef]
- Yuan, L.; Zhu, Y.; Lin, Z.Q.; Banuelos, G.; Li, W.; Yin, X. A novel selenocystine-accumulating plant in selenium-mine drainage area in Enshi, China. PLoS ONE 2013, 8, e65615. [Google Scholar] [CrossRef]
- Both, E.B.; Stonehouse, G.C.; Lima, L.W.; Fakra, S.C.; Aguirre, B.; Wangeline, A.L.; Xiang, J.; Yin, H.; Jókai, Z.; Soós, Á.; et al. Selenium tolerance, accumulation, localization and speciation in a Cardamine hyperaccumulator and a non-hyperaccumulator. Sci. Total Environ. 2020, 703, 135041. [Google Scholar] [CrossRef]
- Boojar, M.M.; Tavakkoli, Z. New molybdenum-hyperaccumulator among plant species growing on molybdenum mine-a biochemical study on tolerance mechanism against metal toxicity. J. Plant Nutr. 2011, 34, 1532–1557. [Google Scholar] [CrossRef]
- van Der Ent, A.; Vinya, R.; Erskine, P.D.; Malaisse, F.; Przybyłowicz, W.J.; Barnabas, A.D.; Harris, H.H.; Mesjasz-Przybyłowicz, J. Elemental distribution and chemical speciation of copper and cobalt in three metallophytes from the copper–cobalt belt in Northern Zambia. Metallomics 2020, 12, 682–701. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, M.; Li, T.; Xu, X.; Deng, L. A newly found cadmium accumulator—Malva sinensis Cavan. J. Hazard. Mater. 2010, 173, 705–759. [Google Scholar] [CrossRef] [PubMed]
- Shengguo, X.U.; Yingxu, C.H.; Qi, L.I.; Shengyou, X.; Yuanpeng, W. Phytolacca acinosa Roxb. (Phytolaccaceae): A new manganese hyperaccumulator plant from Southern China. Acta Ecol. Sin. 2003, 23, 935–937. [Google Scholar]
- Min, Y.; Meizhen, T.; Aoyama, I. Accumulation and uptake of manganese in a hyperaccumulator Phytolacca americana. Miner. Eng. 2007, 20, 188–190. [Google Scholar] [CrossRef]
- Liu, P.; Tang, X.; Gong, C.; Xu, G. Manganese tolerance and accumulation in six Mn hyperaccumulators or accumulators. Plant Soil 2010, 335, 385–395. [Google Scholar] [CrossRef]
- Liu, C.; Liu, W.S.; van der Ent, A.; Morel, J.L.; Zheng, H.X.; Wang, G.B.; Tang, Y.T.; Qiu, R.L. Simultaneous hyperaccumulation of rare earth elements, manganese and aluminum in Phytolacca americana in response to soil properties. Chemosphere 2021, 282, 131096. [Google Scholar] [CrossRef]
- Deng, H.; Li, M.S.; Chen, Y.X. Accumulating characteristics of manganese by Polygonum pubescens Blume. Acta Ecol. Sin. 2009, 29, 5450–5454. [Google Scholar]
- Vondráčková, S.; Száková, J.; Drábek, O.; Tejnecký, V.; Hejcman, M.; Müllerová, V.; Tlustoš, P. Aluminium uptake and translocation in Al hyperaccumulator Rumex obtusifolius is affected by low-molecular-weight organic acids content and soil pH. PLoS ONE 2015, 10, e0123351. [Google Scholar] [CrossRef]
- Tolrà, R.P.; Poschenrieder, C.; Luppi, B.; Barceló, J. Aluminium-induced changes in the profiles of both organic acids and phenolic substances underlie Al tolerance in Rumex acetosa. L. Environ. Exp. Bot. 2005, 54, 231–238. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J.; Borhidi, A.; Berazain, R. Nickel hyperaccumulation in the serpentine flora of Cuba. Ann. Bot. 1999, 83, 29–38. [Google Scholar] [CrossRef]
- Wei, S.H.; Zhou, Q.X.; Wang, X. Cadmium-hyperaccumulator Solanum nigrum L. and its accumulating characteristics. Huan Jing Ke Xue 2005, 26, 167–171. (In Chinese) [Google Scholar] [PubMed]
- Bech, J.; Roca, N.; Tume, P.; Ramos-Miras, J.; Gil, C.; Boluda, R. Screening for new accumulator plants in potential hazards elements polluted soil surrounding Peruvian mine tailings. Catena 2016, 136, 66–73. [Google Scholar] [CrossRef]
- Hadi, F.; Ahmad, A.; Ali, N. Cadmium (Cd) removal from saline water by Veronica anagallis and Epilobium laxum plants in hydroponic system. Agric. Sci. 2014, 5, 935. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H. (Eds.) Methods of Soil Analysis; Part 3: Chemical Methods; John Wiley & Sons: Hoboken, NJ, USA, 2020; ISBN 978-0-891-18825-4. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Family | Al Concentration (µg g−1 DW) |
|---|---|---|
| Amaranthus blitoides S. Watson | Amaranthaceae | 1930–2992 |
| Amaranthus blitum L. | Amaranthaceae | 1578–2672 |
| Amaranthus retroflexus L. | Amaranthaceae | 1108–1655 |
| Barbula unguiculata Hedw. | Pottiaceae | 2132–5752 |
| Brachythecium rutabulum (Hedw.) Schimp. | Brachytheciaceae | 1180–1292 |
| Calystegia sepium (L.) R. Br. | Convolvulaceae | 1120–1139 |
| Campanula rapunculus L. subsp. lambertiana (A.DC.) Rech. f. | Campanulaceae | 1209–1495 |
| Cardamine hirsuta L. | Brassicaceae | 1570–2981 |
| Carex divulsa Stokes | Cyperaceae | 1010–1295 |
| Equisetum telmateia Ehrh. | Equisetaceae | 1183–2114 |
| Erigeron canadensis L. | Asteraceae | 1023–2028 |
| Geranium dissectum L. | Geraniaceae | 2072–2153 |
| Geranium pyrenaicum Burm. f. | Geraniaceae | 1005–1101 |
| Hypnum cupressiforme Hedw. | Hypnaceae | 1102–1501 |
| Lamium album L. | Lamiaceae | 2151–4061 |
| Malva neglecta Wallr. | Malvaceae | 1635–2237 |
| Mentha aquatica L. | Lamiaceae | 1654–5336 |
| Mentha pulegium L. | Lamiaceae | 1034–1511 |
| Microthlaspi umbellatum F. K. Mey. | Brassicaceae | 1011–1342 |
| Oxalis acetosella L. | Oxalidaceae | 1420–1954 |
| Phytolacca americana L. | Phytolaccaceae | 1098–1156 |
| Poa annua L. | Poaceae | 1615–3700 |
| Polygonum aviculare L. | Polygonaceae | 1578–3627 |
| Polygonum hydropiper L. | Polygonaceae | 1030–5008 |
| Primula heterochroma Stapf | Primulaceae | 1250–1581 |
| Prunella vulgaris L. | Lamiaceae | 1023–1782 |
| Rumex conglomeratus Murray | Polygonaceae | 1311–1522 |
| Senecio vernalis Waldst. & Kit. | Asteraceae | 1159–1209 |
| Setaria viridis (L.) P. Beauv. | Poaceae | 1240–1618 |
| Sigesbeckia orientalis L. | Asteraceae | 1086–1601 |
| Solanum nigrum L. | Solanaceae | 1143–1208 |
| Torilis japonica (Huott.) DC. | Apiaceae | 1076–1412 |
| Trifolium repens L. | Fabaceae | 3707–4299 |
| Urtica dioica L. | Urticaceae | 1007–2095 |
| Veronica persica Poir. | Plantaginaceae | 1194–2044 |
| Willemetia tuberosa Fisch. & C. A. Mey. Ex DC. | Asteraceae | 1456–3376 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hajiboland, R.; Moradi, A.; Kahneh, E.; Poschenrieder, C.; Nazari, F.; Pavlovic, J.; Tolra, R.; Salehi-Lisar, S.-Y.; Nikolic, M. Weed Species from Tea Gardens as a Source of Novel Aluminum Hyperaccumulators. Plants 2023, 12, 2129. https://doi.org/10.3390/plants12112129
Hajiboland R, Moradi A, Kahneh E, Poschenrieder C, Nazari F, Pavlovic J, Tolra R, Salehi-Lisar S-Y, Nikolic M. Weed Species from Tea Gardens as a Source of Novel Aluminum Hyperaccumulators. Plants. 2023; 12(11):2129. https://doi.org/10.3390/plants12112129
Chicago/Turabian StyleHajiboland, Roghieh, Aiuob Moradi, Ehsan Kahneh, Charlotte Poschenrieder, Fatemeh Nazari, Jelena Pavlovic, Roser Tolra, Seyed-Yahya Salehi-Lisar, and Miroslav Nikolic. 2023. "Weed Species from Tea Gardens as a Source of Novel Aluminum Hyperaccumulators" Plants 12, no. 11: 2129. https://doi.org/10.3390/plants12112129