
Hormonal Balance, Photosynthesis, and Redox Reactions in the Leaves of Caragana korshinskii Kom. under Water Deficit

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation and Experimental Design
2.2. Gas Exchange Measurements
2.3. Measurements of Leaf Chlorophyll Fluorescence
2.4. Determination of O2•−, H2O2, MDA, and the Activities of Antioxidant Enzymes
2.5. Determination of Phytohormones
2.6. Statistical Analysis
3. Results
3.1. Growth Performance
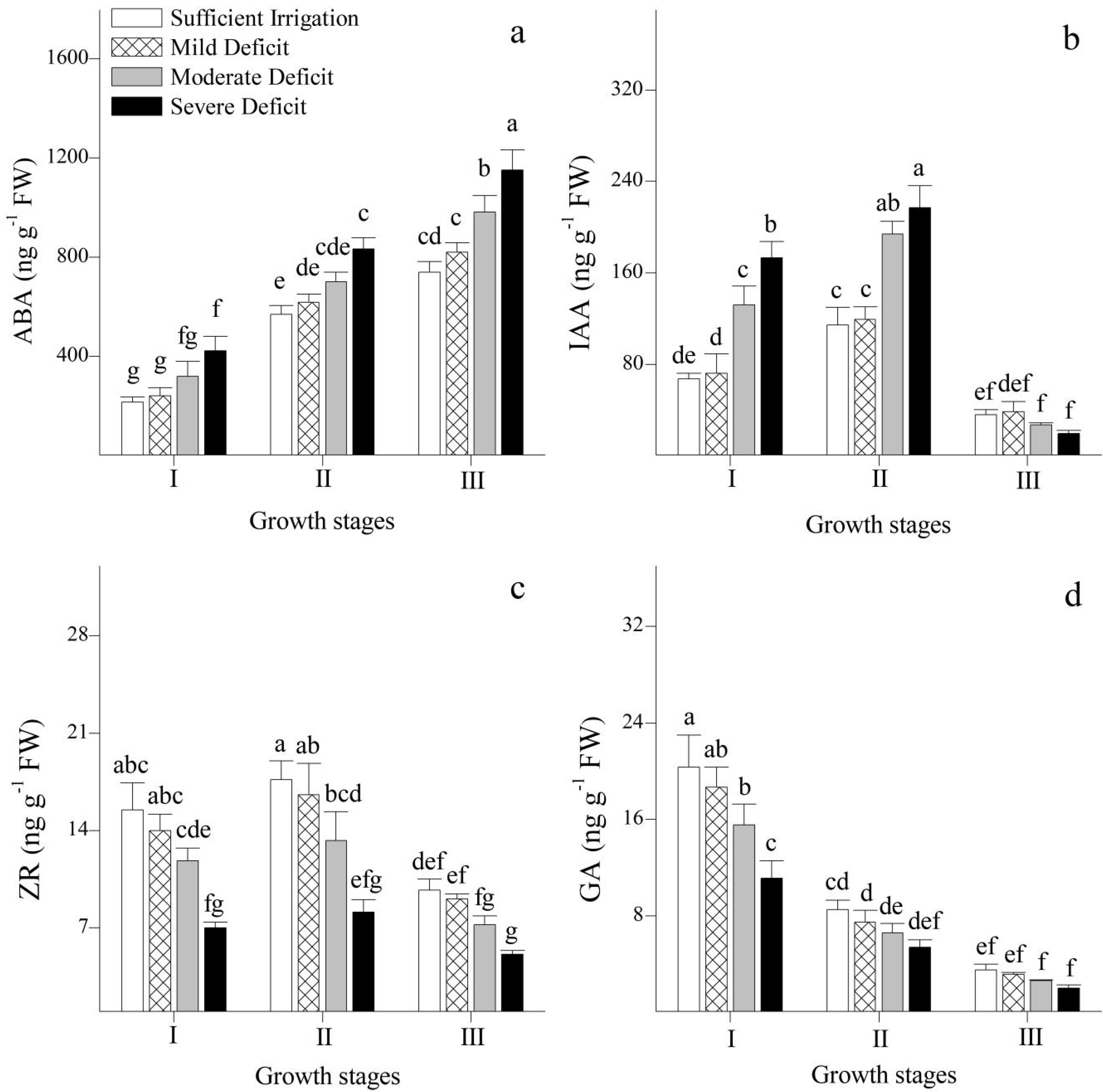
3.2. Endogenous Hormones and Their Ratios in Leaves
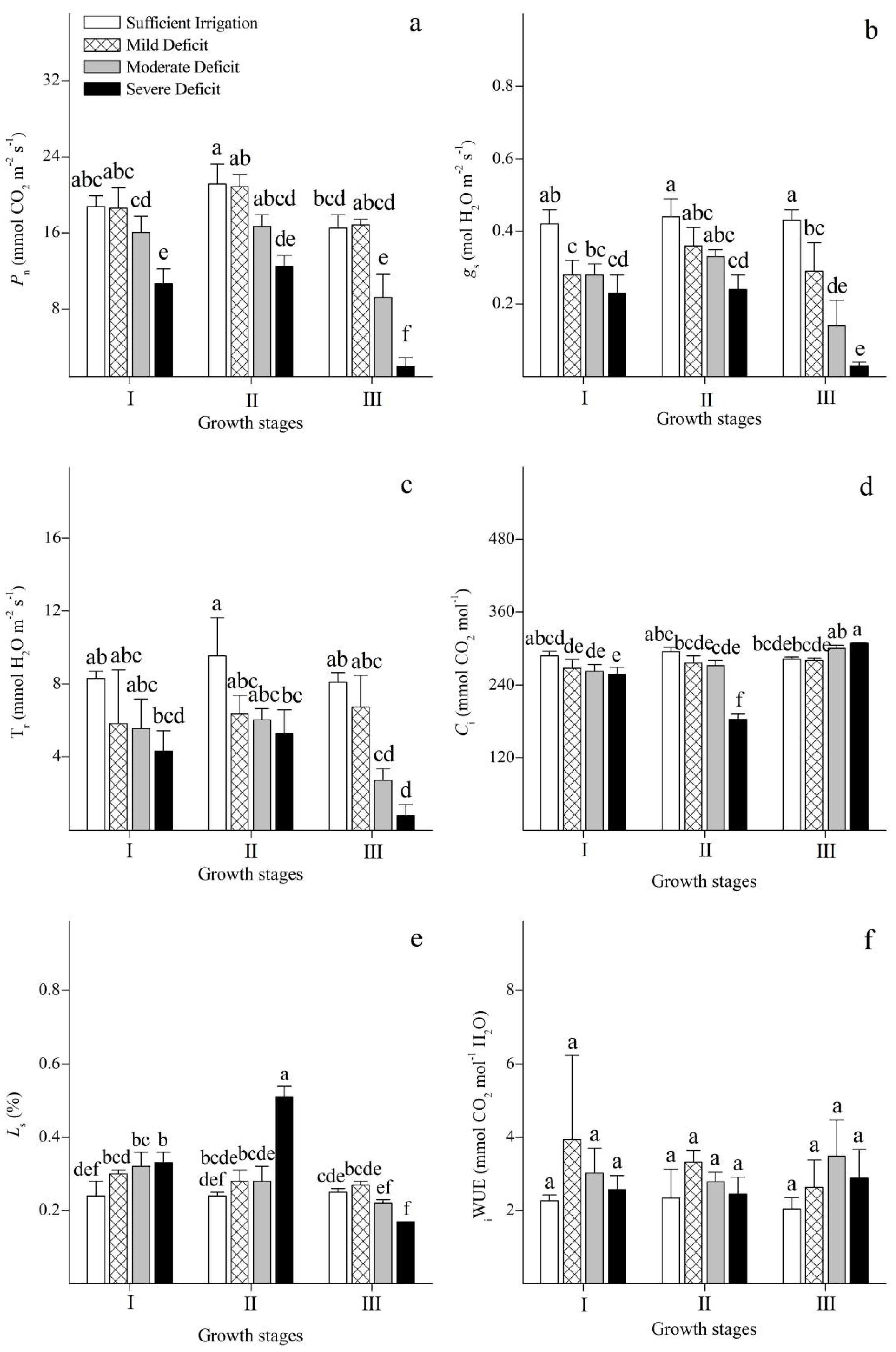
3.3. Photosynthetic Gas Exchange
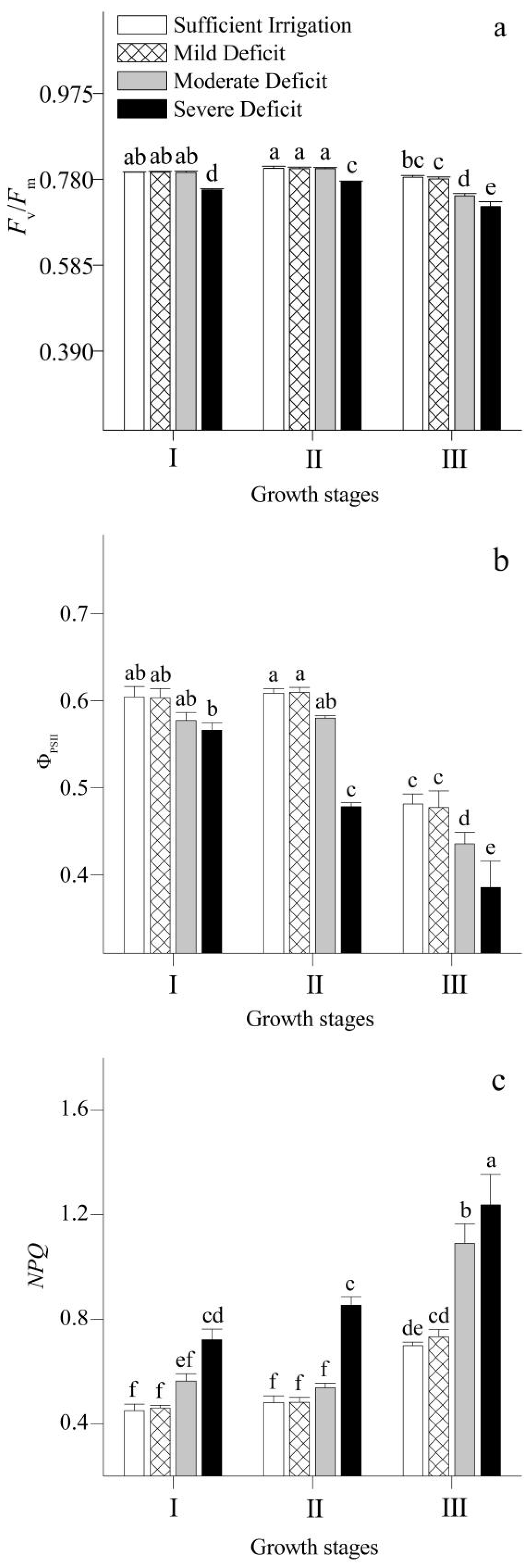
3.4. Chlorophyll Fluorescence
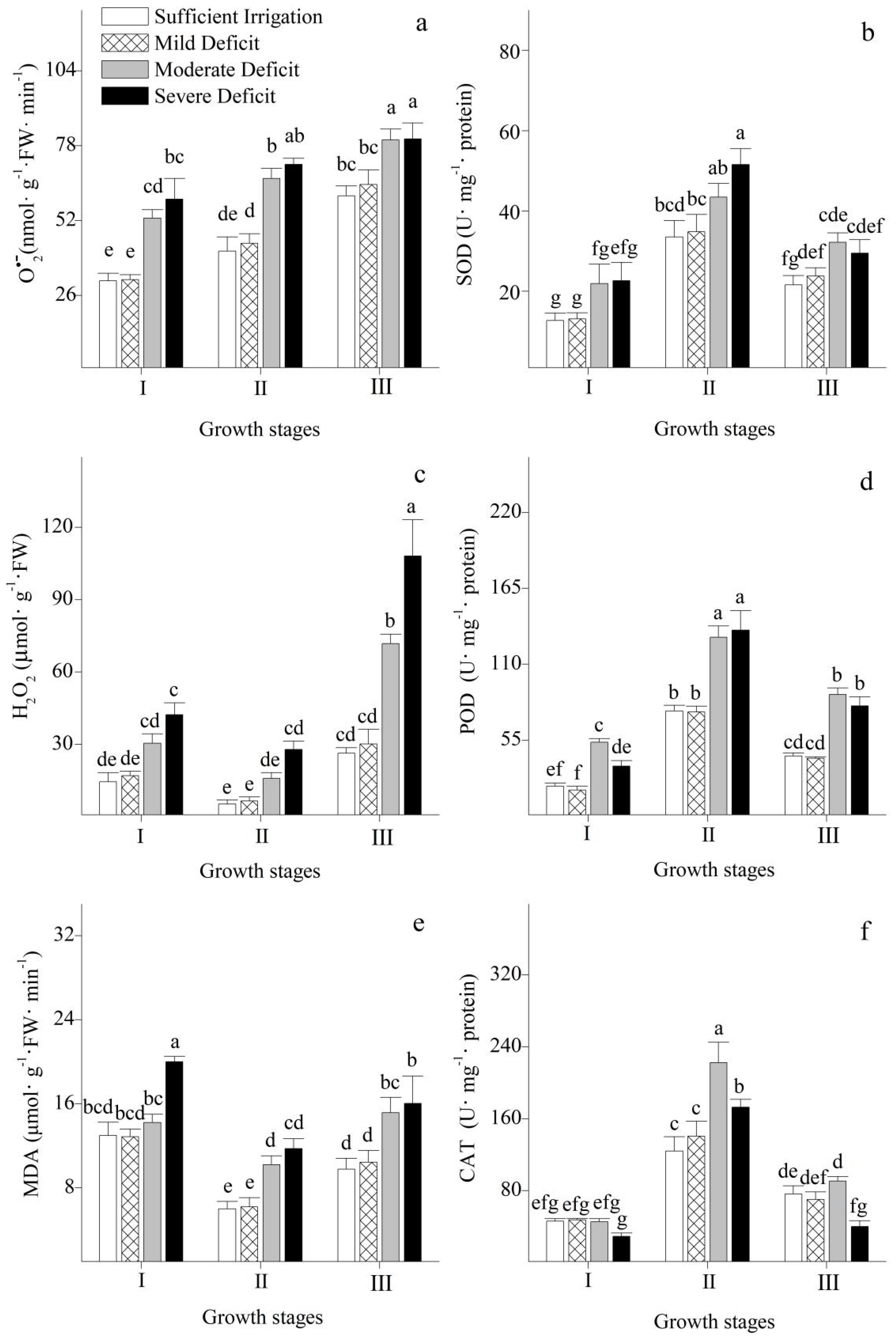
3.5. Oxidative Damage and ROS-Scavenging Systems
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fang, X.W.; Turner, N.C.; Li, F.M.; Li, W.J.; Guo, X.S. Caragana korshinskii seedlings maintain positive photosynthesis during short-term, severe drought stress. Photosynthetica 2011, 49, 603–609. [Google Scholar] [CrossRef]
- Yan, H.; Hu, X.; Li, F. Leaf photosynthesis, chlorophyll fluorescence, ion content and free amino acids in Caragana korshinskii Kom exposed to NaCl stress. Acta Physiol. Plant. 2012, 34, 2285–2295. [Google Scholar] [CrossRef]
- Llanes, A.S.; Andrade, A.M.; Alemano, S.G.; Luna, M.V. Alterations of endogenous hormonal levels in plants under drought and salinity. Am. J. Plant Sci. 2016, 7, 1357. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Cheng, Z.; Meng, H.; Tang, X. The garlic allelochemical diallyl disulfide affects tomato root growth by influencing cell division, phytohormone balance and expansin gene expression. Front. Plant. Sci. 2016, 7, 1199. [Google Scholar] [CrossRef] [PubMed]
- Davies, W.J.; Kudoyarova, G.; Hartung, W. Long-distance ABA signaling and its relation to other signaling pathways in the detection of soil drying and the mediation of the plant’s response to drought. J. Plant Growth Regul. 2005, 24, 285–295. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; He, Q.; Zhou, H. Stomatal limitations to photosynthesis and their critical Water conditions in different growth stages of maize under water stress. Agric. Water Manag. 2020, 241, 106330. [Google Scholar] [CrossRef]
- Khatri, K.; Rathore, M.S. Photosystem photochemistry, prompt and delayed fluorescence, photosynthetic responses and electron flow in tobacco under drought and salt stress. Photosynthetica 2019, 57, 61–74. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Ramakrishnan, M.; Sidhu, G.P.S.; Bali, A.S.; Handa, N.; Kapoor, D.; Yadav, P.; Khanna, K.; et al. Photosynthetic response of plants under different abiotic stresses: A review. J. Plant Growth Regul. 2020, 39, 509–531. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Cifre, J.; Escalona, J.M.; Galmés, J.; Guléas, J.; Lefi, E.K.; Martínez-Cañellas, S.F.; Moreno, M.T.; Ribas-Carbó, M.; et al. Understanding down-ragulation of photosynthesis under water stress: Future prospects and searching for physiological tools for irrigation management. Annl. Appl. Biol. 2004, 144, 273–283. [Google Scholar] [CrossRef]
- Yuan, X.T.; Li, F.P.; Gu, H.H. Effects of high temperature on photosynthetic capacity in the leaves of creepers. Scienceasia 2020, 46, 436–443. [Google Scholar]
- Hu, W.H.; Wu, Y.; Zeng, J.Z.; He, L.; Zeng, Q.M. Chill-induced inhibition of photosynthesis was alleviated by 24-epibrassinolide pretreatment in cucumber during chilling and subsequent recovery. Photosynthetica 2010, 48, 537–544. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Yin, H.; Chen, Q.; Yi, M. Effects of short-term heat stress on oxidative damage and responses of antioxidant system in Lilium longiflorum. Plant Growth Regul. 2008, 54, 45–54. [Google Scholar] [CrossRef]
- Wu, X.H.; Wang, W.; Xie, X.L.; Yin, C.M.; Xie, K.J. Photosynthetic and yield responses of rice (Oryza sativa L.) to different water management strategies in subtropical China. Photosynthetica 2018, 56, 1031–1038. [Google Scholar]
- Zhou, H.; Beynon-Davies, R.; Carslaw, N.; Dodd, I.C.; Ashworth, K. Yield, resource use efficiency or flavour: Trade-offs of varying blue-to-red lighting ratio in urban plant factories. Sci. Hortic. 2022, 295, 110802. [Google Scholar] [CrossRef]
- Rajendran, D.K.; Park, E.; Nagendran, R.; Hung, N.B.; Cho, B.K.; Kim, K.H.; Lee, Y.H. Visual analysis for detection and quantification of Pseudomonas cichorii disease severity in tomato plants. Plant Pathol. J. 2016, 32, 300–310. [Google Scholar] [CrossRef]
- He, J.; Qin, J.; Long, L.; Ma, Y.; Li, H.; Li, K.; Jiang, X.; Liu, T.; Polle, A.; Liang, Z.; et al. Net cadmium flux and accumulation reveal tissue-specific oxidative stress and detoxification in Populus × canescens. Physiol. Plant. 2011, 143, 50–63. [Google Scholar] [CrossRef]
- Yan, H.; Filardo, F.; Hu, X.; Zhao, X.; Fu, D.H. Cadmium stress alters the redox reaction and hormone balance in oilseed rape (Brassica napus L.) leaves. Environ. Sci. Pollut. Res. 2016, 23, 3758–3769. [Google Scholar]
- Luo, Z.B.; Calfapietra, C.; Scarascia-Mugnozza, G.; Liberloo, M.; Polle, A. Carbon-based secondary metabolites and internal nitrogen pools in Populus nigra under free air CO2 enrichment (FACE) and nitrogen fertilisation. Plant. Soil 2008, 30, 45–57. [Google Scholar] [CrossRef]
- He, Z. A Laboratory Guide to Chemical Control Technology on Field Crop; Beijing Agricultural University Press: Beijing, China, 1993; pp. 60–68. [Google Scholar]
- Bray, E.A. Abscisic acid regulation of gene expression during water-deficit stress in the era of the Arabidopsis genome. Plant Cell Environ. 2002, 25, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Cui, N.; Zhou, H. Hormonal regulation and redox reaction of maize under partial root drying irrigation. Acta Physiol. Plant. 2019, 41, 116. [Google Scholar] [CrossRef]
- Yan, H.; Wu, L.; Filardo, F.; Yang, X.; Zhao, X.; Fu, D. Chemical and hydraulic signals regulate stomatal behavior and photosynthetic activity in maize during progressive drought. Acta Physiol. Plant. 2017, 39, 125. [Google Scholar] [CrossRef]
- Pustovoitova, T.N.; Zhdanova, N.E.; Zholkevich, V.N. Epibrassinolide Increases Plant Drought Resistance. Dokl. Biochem. Biophys. 2001, 376, 36–38. [Google Scholar] [CrossRef]
- Pustovoitova, T.N.; Zhdanova, N.E.; Zholkevich, V.N. Changes in the levels of IAA and ABA in cucumber leaves under progressive soil drought. Russ. J. Plant. Physl. 2004, 51, 513–517. [Google Scholar] [CrossRef]
- Ghanem, M.E.; Albacete, A.; Martínez-Andújar, C.; Acosta, M.; Romero-Aranda, R.; Dodd, I.C.; Lutts, S.; Pérez-Alfocea, F. Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). J. Exp. Bot. 2008, 59, 3039–3050. [Google Scholar] [CrossRef]
- Huang, H.J.; Yang, Z.Q.; Zhang, M.Y.; Li, Y.X.; Zhang, J.H.; Hou, M.Y. Effects of water stress on growth, photosynthesis, root activity and endogenous hormones of Cucumis sativus. Int. J. Agric. Biol. 2018, 20, 2579–2589. [Google Scholar]
- Liang, C.; Liu, H. Response of hormone in rice seedlings to irrigation contaminated with cyanobacterial extract containing microcystins. Chemosphere 2020, 256, 127157. [Google Scholar] [CrossRef]
- Lee, T.Y.; Woo, S.Y.; Kwak, M.J.; Inkyin, K.; Lee, K.E.; Jang, J.H.; Kim, I.R. Photosynthesis and chlorophyll fluorescence responses of Populus sibirica to water deficit in a desertification area in Mongolia. Photosynthetica 2016, 54, 317–320. [Google Scholar] [CrossRef]
- Wilkinson, S.; Davies, W.J. ABA-based chemical signalling: The co-ordination of responses to stress in plants. Plant Cell Environ. 2002, 25, 195–210. [Google Scholar] [CrossRef]
- Sarabi, B.; Fresneau, C.; Ghaderi, N.; Bolandnazar, S.; Streb, P.; Badeck, F.W.; Citerne, S.; Tangama, M.; David, A.; Ghashghaie, J. Stomatal and non-stomatal limitations are responsible in down-regulation of photosynthesis in melon plants grown under the saline condition: Application of carbon isotope discrimination as a reliable proxy. Plant. Physiol. Biochem. 2019, 141, 1–19. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Q.; Ma, K.; Chen, L. Temperature-dependent gas exchange and stomatal/non-stomatal limitation to CO2 assimilation of Quercus liaotungensis under midday high irradiance. Photosynthetica 2001, 39, 383–388. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Kannan, N.D.; Kulandaivelu, G. Drought induced changes in physiological, biochemical and phytochemical properties of Withania somnifera. Dun. J. Med. Plants Res. 2011, 5, 3929–3935. [Google Scholar]
- Elstner, E.F. Oxygen activation and oxygen toxicity. Annu. Rev. Plant. Biol. 1982, 33, 73–96. [Google Scholar] [CrossRef]
- Ort, D.R.; Baker, N.R. A photoprotective role for O2 as an alternative electron sink in photosynthesis? Curr. Opin. Plant. Biol. 2002, 5, 193–198. [Google Scholar] [CrossRef]
- Karuppanapandian, T.; Moon, J.C.; Kim, C.; Manoharan, K.; Kim, W. Reactive oxygen species in plants: Their generation, signal transduction, and scavenging mechanisms. Aust. J. Crop Sci. 2011, 5, 709–725. [Google Scholar]
- Jiang, M.; Zhang, J. Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, H.; Liu, X.; Ding, H.; Dai, Z.; Niu, X.; Zhao, L. Hormonal Balance, Photosynthesis, and Redox Reactions in the Leaves of Caragana korshinskii Kom. under Water Deficit. Plants 2023, 12, 2076. https://doi.org/10.3390/plants12112076
Yan H, Liu X, Ding H, Dai Z, Niu X, Zhao L. Hormonal Balance, Photosynthesis, and Redox Reactions in the Leaves of Caragana korshinskii Kom. under Water Deficit. Plants. 2023; 12(11):2076. https://doi.org/10.3390/plants12112076
Chicago/Turabian StyleYan, Hui, Xiaoli Liu, Hao Ding, Zhiguang Dai, Xiaoli Niu, and Long Zhao. 2023. "Hormonal Balance, Photosynthesis, and Redox Reactions in the Leaves of Caragana korshinskii Kom. under Water Deficit" Plants 12, no. 11: 2076. https://doi.org/10.3390/plants12112076