


Gintonin Isolated from Ginseng Inhibits the Epithelial—Mesenchymal Transition Induced by TGF-β in A549 Lung Cancer Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
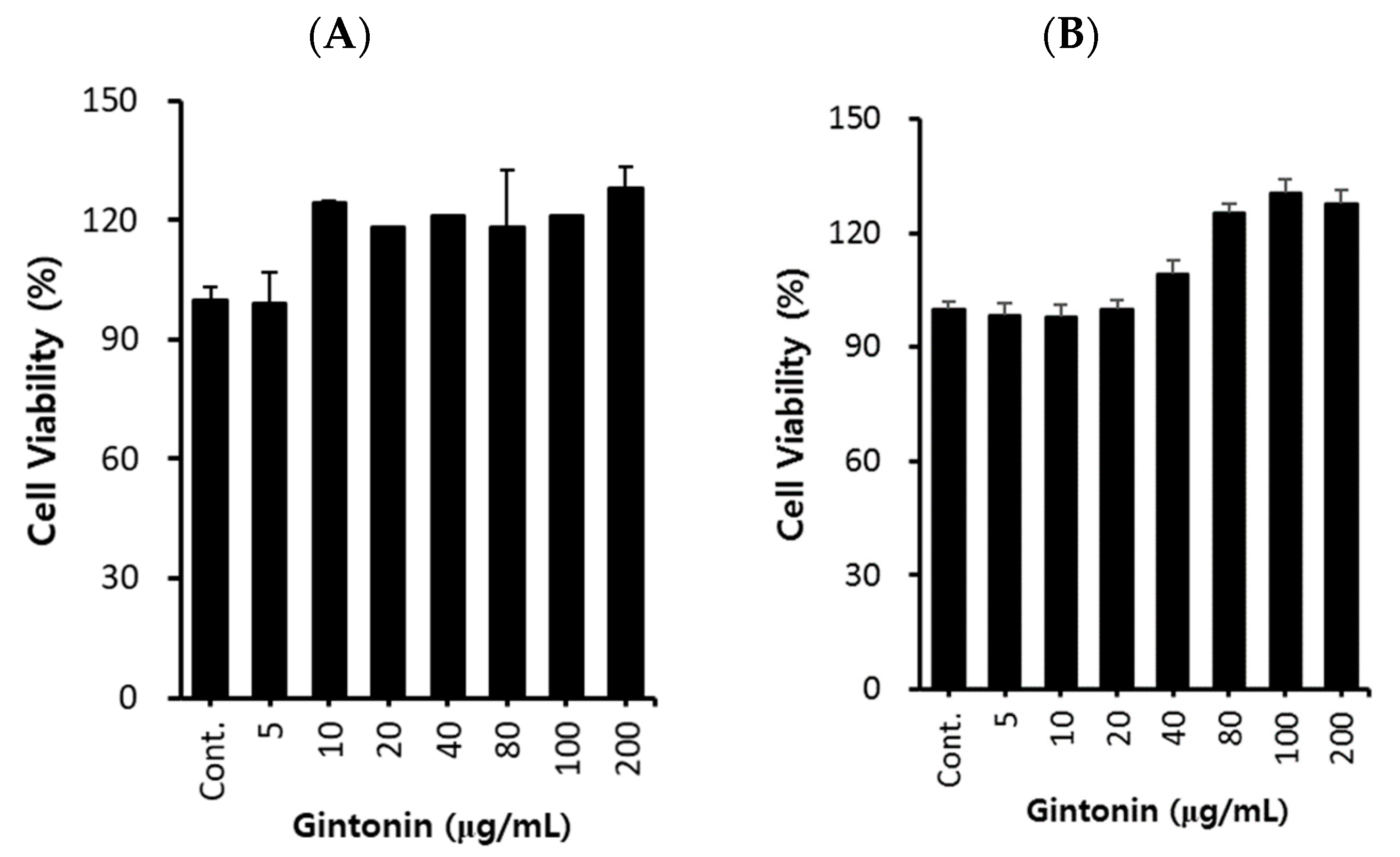
2.1. Effect of GT on A549 Cell Proliferation
2.2. GT Suppressed the Expression of Transcription Factors Associated with TGF-β1-Induced EM Transition in A549 Cells
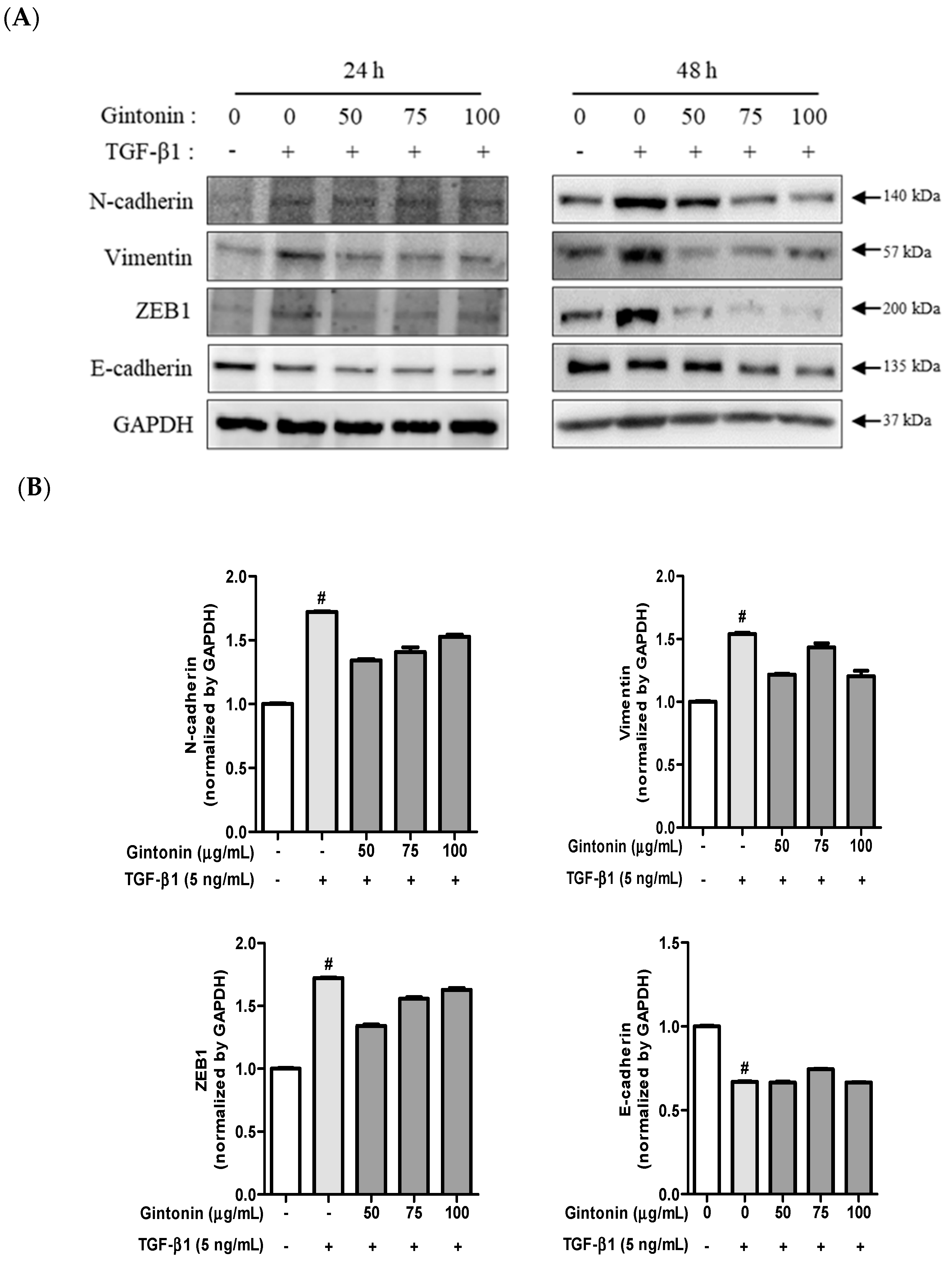
2.3. GT Suppressed the Expression of TGF-β1-Induced N-cad, Vim, and ZEB1 in A549 Cells
2.4. GT Suppressed the mRNA Expression of TGF-β1-Induced Mesenchymal Markers in A549 Cells
2.5. Effect of GT on Morphological Changes and Mesenchymal Cell Markers in TGF-β1-Induced EM Transition
3. Materials and Methods
3.1. Antibodies and Reagents
3.2. Preparation of GT from Ginseng
3.3. Cell Culture
3.4. A549 Cell Viability Analysis
3.5. Induction of EM Transition via TGF-β1 Treatment
3.6. Immunoblotting
3.7. Real-Time Quantitative Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
3.8. Immunofluorescence Staining for Confocal Microscopy
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schabath, M.B.; Cote, M.L. Cancer progress and priorities: Lung cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Travis, W.D.; Brambilla, E.; Riely, G.J. New pathologic classification of lung cancer: Relevance for clinical practice and clinical trials. J. Clin. Oncol. 2013, 31, 992–1001. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, N.; Gheldof, A.; Tatari, M.; Christofori, G. EMT as the ultimate survival mechanism of cancer cells. Semin. Cancer Biol. 2012, 22, 194–207. [Google Scholar] [CrossRef]
- Vleminckx, K.; Vakaet Jr, L.; Mareel, M.; Fiers, W.; Van Roy, F. Genetic manipulation of E-cadherin expression by epithelial tumor cells reveals an invasion suppressor role. Cell 1991, 66, 107–119. [Google Scholar] [CrossRef]
- Peinado, H.; Olmeda, D.; Cano, A. Snail, Zeb and bHLH factors in tumour progression: An alliance against the epithelial phenotype. Nat. Rev. Cancer 2007, 7, 415–428. [Google Scholar] [CrossRef]
- Jakowlew, S.B. Transforming growth factor-β in cancer and metastasis. Cancer Metastasis Rev. 2006, 25, 435–457. [Google Scholar] [CrossRef]
- Massagué, J. TGF-β in cancer. Cell 2008, 134, 215–230. [Google Scholar] [CrossRef]
- Derynck, R.; Zhang, Y.E. Smad-dependent and Smad-independent pathways in TGF-β family signalling. Nature 2003, 425, 577–584. [Google Scholar] [CrossRef]
- Lee, S.M.; Bae, B.S.; Park, H.W.; Ahn, N.G.; Cho, B.G.; Cho, Y.L.; Kwak, Y.S. Characterization of Korean Red Ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng Res. 2015, 39, 384–391. [Google Scholar] [CrossRef]
- Peng, Y.; Pan, W.; Cao, X.; Liu, C. Potential Oral Health Benefits of Ginseng and Its Extracts. Int. Dent. J. 2023, in press.
- Hwang, S.-H.; Lee, B.-H.; Choi, S.-H.; Kim, H.-J.; Jung, S.-W.; Kim, H.-S.; Shin, H.-C.; Park, H.J.; Park, K.H.; Lee, M.K.; et al. Gintonin, a novel ginseng-derived lysophosphatidic acid receptor ligand, stimulates neurotransmitter release. Neurosci. Lett. 2015, 584, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Im, D.-s.; Nah, S.-y. Yin and Yang of ginseng pharmacology: Ginsenosides vs GT. Acta Pharmacol. Sin. 2013, 34, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Pyo, M.K.; Choi, S.-H.; Shin, T.-J.; Hwang, S.H.; Lee, B.-H.; Kang, J.; Kim, H.-J.; Lee, S.-H.; Nah, S.-Y. A simple method for the preparation of crude GT from ginseng root, stem, and leaf. J. Ginseng. Res. 2011, 35, 209. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.; Hla, T.; Lynch, K.R.; Spiegel, S.; Moolenaar, W.H. International union of basic and clinical pharmacology. LXXVIII. Lysophospholipid receptor nomenclature. Pharmacol. Rev. 2010, 62, 579–587. [Google Scholar] [CrossRef]
- Hwang, S.H.; Shin, T.J.; Choi, S.H.; Cho, H.J.; Lee, B.H.; Pyo, M.K.; Nah, S.Y. Gintonin, newly identified compounds from ginseng, is novel lysophosphatidic acids-protein complexes and activates G protein-coupled lysophosphatidic acid receptors with high affinity. Mol. Cells 2012, 33, 151–162. [Google Scholar] [CrossRef]
- Hwang, S.H.; Shin, E.-J.; Shin, T.-J.; Lee, B.-H.; Choi, S.-H.; Kang, J.; Kim, H.-J.; Kwon, S.-H.; Jang, C.-G.; Lee, J.-H. Gintonin, a ginseng-derived lysophosphatidic acid receptor ligand, attenuates Alzheimer’s disease-related neuropathies: Involvement of non-amyloidogenic processing. J. Alzheimer’s Dis. 2012, 31, 207–223. [Google Scholar] [CrossRef]
- Moon, J.; Choi, S.H.; Shim, J.Y.; Park, H.J.; Oh, M.J.; Kim, M.; Nah, S.Y. Gintonin administration is safe and potentially beneficial in cognitively impaired elderly. Alzheimer Dis. Assoc. Disord. 2018, 32, 85–87. [Google Scholar] [CrossRef]
- Ikram, M.; Ullah, R.; Khan, A.; Kim, M.O. Ongoing research on the role of gintonin in the management of neurodegenerative disorders. Cells. 2020, 9, 1464. [Google Scholar] [CrossRef]
- Kim, H.-J.; Kim, D.-J.; Shin, E.-J.; Lee, B.-H.; Choi, S.-H.; Hwang, S.-H.; Rhim, H.; Cho, I.-H.; Kim, H.-C.; Nah, S.-Y. Effects of gintonin-enriched fraction on hippocampal cell proliferation in wild-type mice and an APPswe/PSEN-1 double Tg mouse model of Alzheimer’s disease. Neurochem. Int. 2016, 101, 56–65. [Google Scholar] [CrossRef]
- Kim, H.J.; Park, S.D.; Lee, R.M.; Lee, B.H.; Choi, S.H.; Hwang, S.H.; Nah, S.Y. Gintonin attenuates depressive-like behaviors associated with alcohol withdrawal in mice. J. Affect. Disord. 2017, 215, 23–29. [Google Scholar] [CrossRef]
- Hwang, S.H.; Lee, B.-H.; Kim, H.-J.; Cho, H.-J.; Shin, H.-C.; Im, K.-S.; Choi, S.-H.; Shin, T.-J.; Lee, S.-M.; Nam, S.W. Suppression of metastasis of intravenously-inoculated B16/F10 melanoma cells by the novel ginseng-derived ingredient, GT: Involvement of autotaxin inhibition. Int. J. Oncol. 2013, 42, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.-H.; Won, K.-J.; Lee, R.; Cho, H.-S.; Hwang, S.-H.; Nah, S.-Y. Wound healing effect of GT involves Lysophosphatidic acid receptor/vascular endothelial growth factor signaling pathway in keratinocytes. Int. J. Mol. Sci. 2021, 22, 10155. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.L.; Robin, T.P.; Ford, H.L. Molecular Pathways: Targeting the TGF-β Pathway for Cancer TherapyTargeting the TGF-β Pathway in Cancer. Clin. Cancer. Res. 2012, 18, 4514–4521. [Google Scholar] [CrossRef]
- Massagué, J.; Gomis, R.R. The logic of TGFβ signaling. FEBS Lett. 2006, 580, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- Mittal, V. Epithelial mesenchymal transition in tumor metastasis. Annu. Rev. Pathol.-Mech. 2018, 13, 395–412. [Google Scholar] [CrossRef]
- Choi, J.H.; Hwang, Y.P.; Kim, H.G.; Khanal, T.; Do, M.T.; Jin, S.W.; Han, H.J.; Lee, H.S.; Lee, Y.C.; Chung, Y.C. Saponins from the roots of Platycodon grandiflorum suppresses TGFβ1-induced epithelial-mesenchymal transition via repression of PI3K/Akt, ERK1/2 and Smad2/3 pathway in human lung carcinoma A549 cells. Nutr. Cancer 2014, 66, 140–151. [Google Scholar] [CrossRef]
- Thiery, J.P.; Acloque, H.; Huang, R.Y.; Nieto, M.A. Epithelial-mesenchymal transitions in development and disease. Cell 2009, 139, 871–890. [Google Scholar] [CrossRef]
- Gonzalez, D.M.; Medici, D. Signaling mechanisms of the epithelial-mesenchymal transition. Sci. Signal 2014, 7, re8. [Google Scholar] [CrossRef]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial–mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef]
- Lee, K.; Jin, H.; Chei, S.; Oh, H.J.; Choi, S.H.; Nah, S.Y.; Lee, B.Y. The gintonin-enriched fraction of ginseng regulates lipid metabolism and browning via the cAMP-protein kinase a signaling pathway in mice white adipocytes. Biomolecules 2020, 10, 1048. [Google Scholar] [CrossRef]
- Zhang, H.J.; Wang, H.Y.; Zhang, H.T.; Su, J.M.; Zhu, J.; Wang, H.B.; Chen, X.F. Transforming growth factor-β1 promotes lung adenocarcinoma invasion and metastasis by epithelial-to-mesenchymal transition. Mol. Cell. Biochem. 2011, 355, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hwan Kim, S. CK2 inhibitor CX-4945 blocks TGF-β1-induced epithelial-to-mesenchymal transition in A549 human lung adenocarcinoma cells. PLoS ONE 2013, 8, e74342. [Google Scholar] [CrossRef] [PubMed]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal. Transduct. Target Ther. 2020, 5, 1–17. [Google Scholar] [CrossRef]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288. [Google Scholar] [CrossRef]
- Ko, H.; So, Y.; Jeon, H.; Jeong, M.H.; Choi, H.K.; Ryu, S.H.; Lee, S.W.; Yoon, H.G.; Choi, K.C. TGF-beta1-induced epithelial-mesenchymal transition and acetylation of Smad2 and Smad3 are negatively regulated by EGCG in human A549 lung cancer cells. Cancer Lett. 2013, 335, 205e13. [Google Scholar] [CrossRef] [PubMed]
- Ko, H.; Jeon, H.; Lee, D.; Choi, H.K.; Kang, K.S.; Choi, K.C. Sanguiin H6 suppresses TGFbeta induction of the epithelial-mesenchymal transition and inhibits migration and invasion in A549 lung cancer. Bioorg. Med. Chem. Lett. 2015, 25, 5508e13. [Google Scholar]
- Ko, H. Geraniin inhibits TGF-beta1-induced epithelial-mesenchymal transition and suppresses A549 lung cancer migration, invasion and anoikis resistance. Bioorg. Med. Chem. Lett. 2015, 25, 3529e34. [Google Scholar]
- Jin, S.; Park, J.Y.; Kim, H.Y.; Kim, Y.; Park, S.K.; Jang, Y.J.; Lee, Y.; Kim, H.J.; Bae, Y.C.; Rhim, H.; et al. Gintonin enhances performance of mice in rotarod test: Involvement of lysophosphatidic acid receptors and catecholamine release. Neurochem. Int. 2014, 75, 24–31. [Google Scholar]
- Choi, S.H.; Lee, B.H.; Kim, H.J.; Jung, S.W.; Lee, E.H.; Kim, H.M.; Kim, K.T.; Rhim, H.; Hwang, S.H.; Nah, S.Y. Inhibitory Effects of Gintonin on Tumor Growth in Colorectal Cancer. J. Ginseng. Res. 2017, 41, 207–213. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.J.; Nah, S.-Y.; Park, I.-H.; Shin, M.-S.; Kang, K.S. Gintonin Isolated from Ginseng Inhibits the Epithelial—Mesenchymal Transition Induced by TGF-β in A549 Lung Cancer Cells. Plants 2023, 12, 2013. https://doi.org/10.3390/plants12102013
Kim SJ, Nah S-Y, Park I-H, Shin M-S, Kang KS. Gintonin Isolated from Ginseng Inhibits the Epithelial—Mesenchymal Transition Induced by TGF-β in A549 Lung Cancer Cells. Plants. 2023; 12(10):2013. https://doi.org/10.3390/plants12102013
Chicago/Turabian StyleKim, Sung Jin, Seung-Yeol Nah, Il-Ho Park, Myoung-Sook Shin, and Ki Sung Kang. 2023. "Gintonin Isolated from Ginseng Inhibits the Epithelial—Mesenchymal Transition Induced by TGF-β in A549 Lung Cancer Cells" Plants 12, no. 10: 2013. https://doi.org/10.3390/plants12102013