
Agathis robusta Bark Essential Oil Effectiveness against COVID-19: Chemical Composition, In Silico and In Vitro Approaches



Abstract
:1. Introduction
2. Results
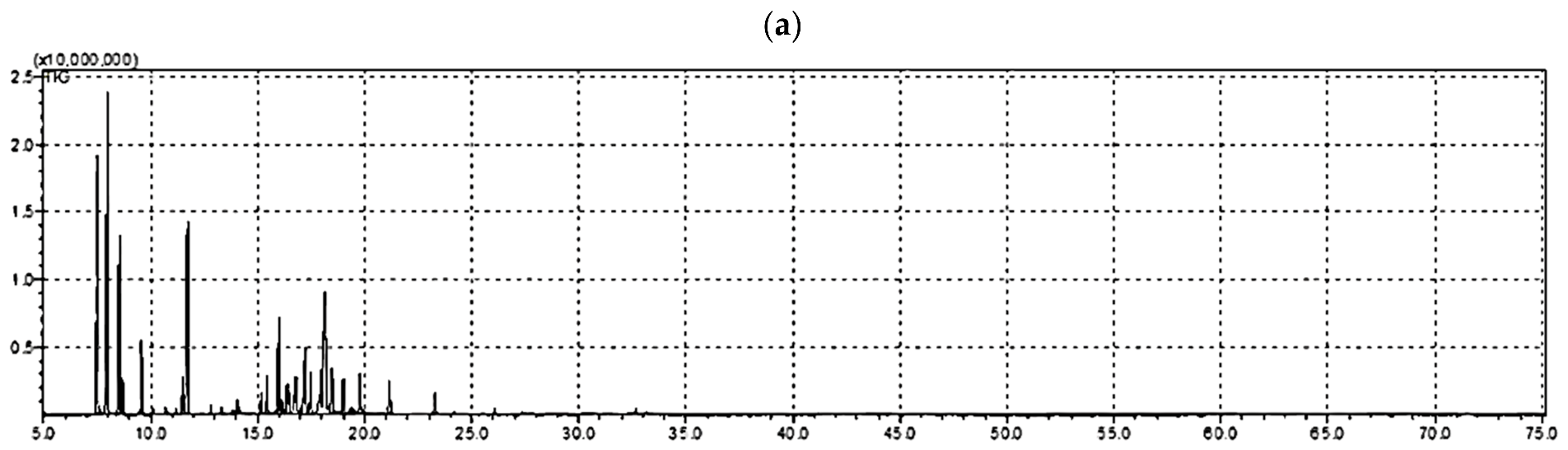
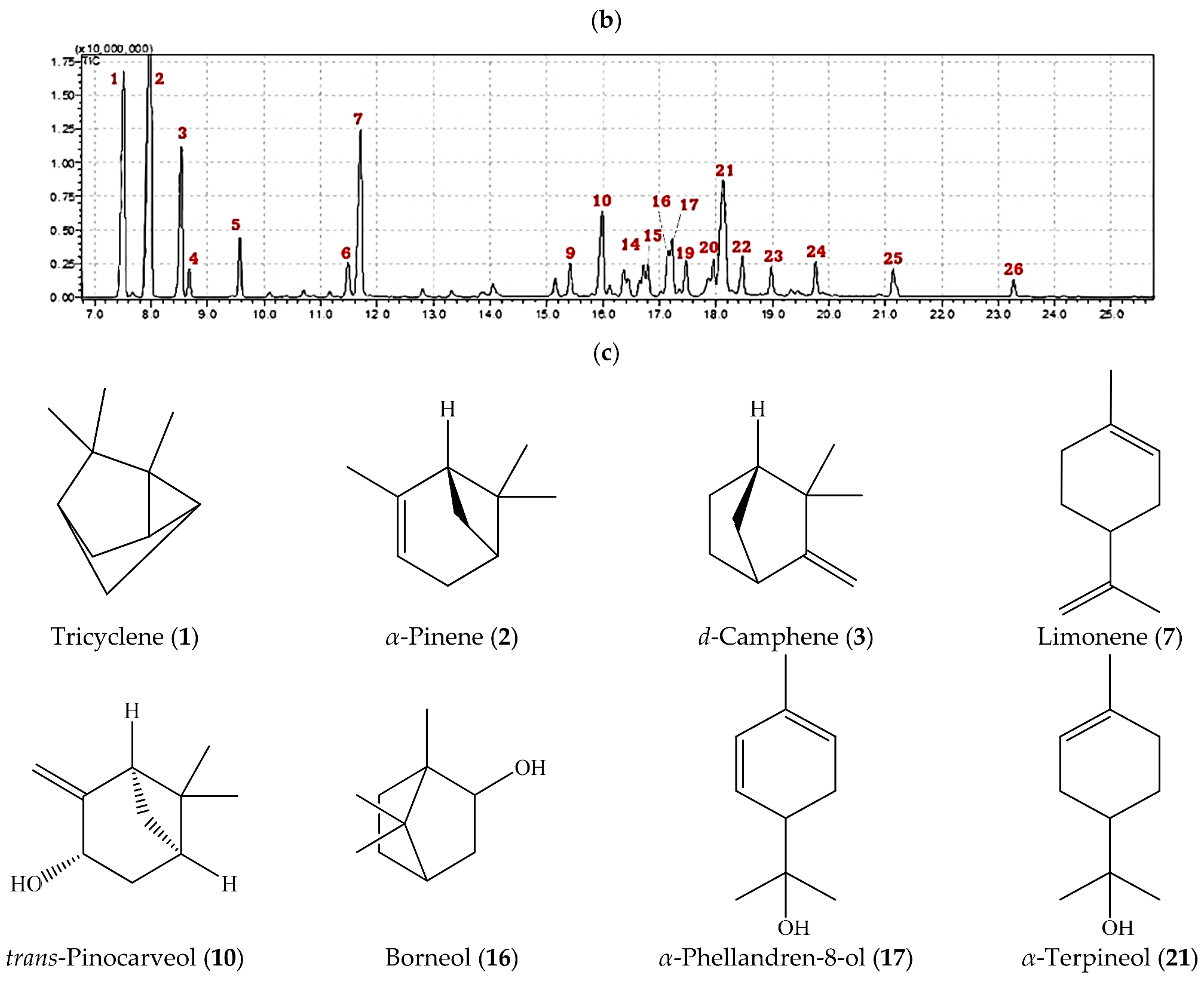
2.1. Volatile Constituents in A. robusta Bark EO
2.2. Molecular Modelling and In Silico Study
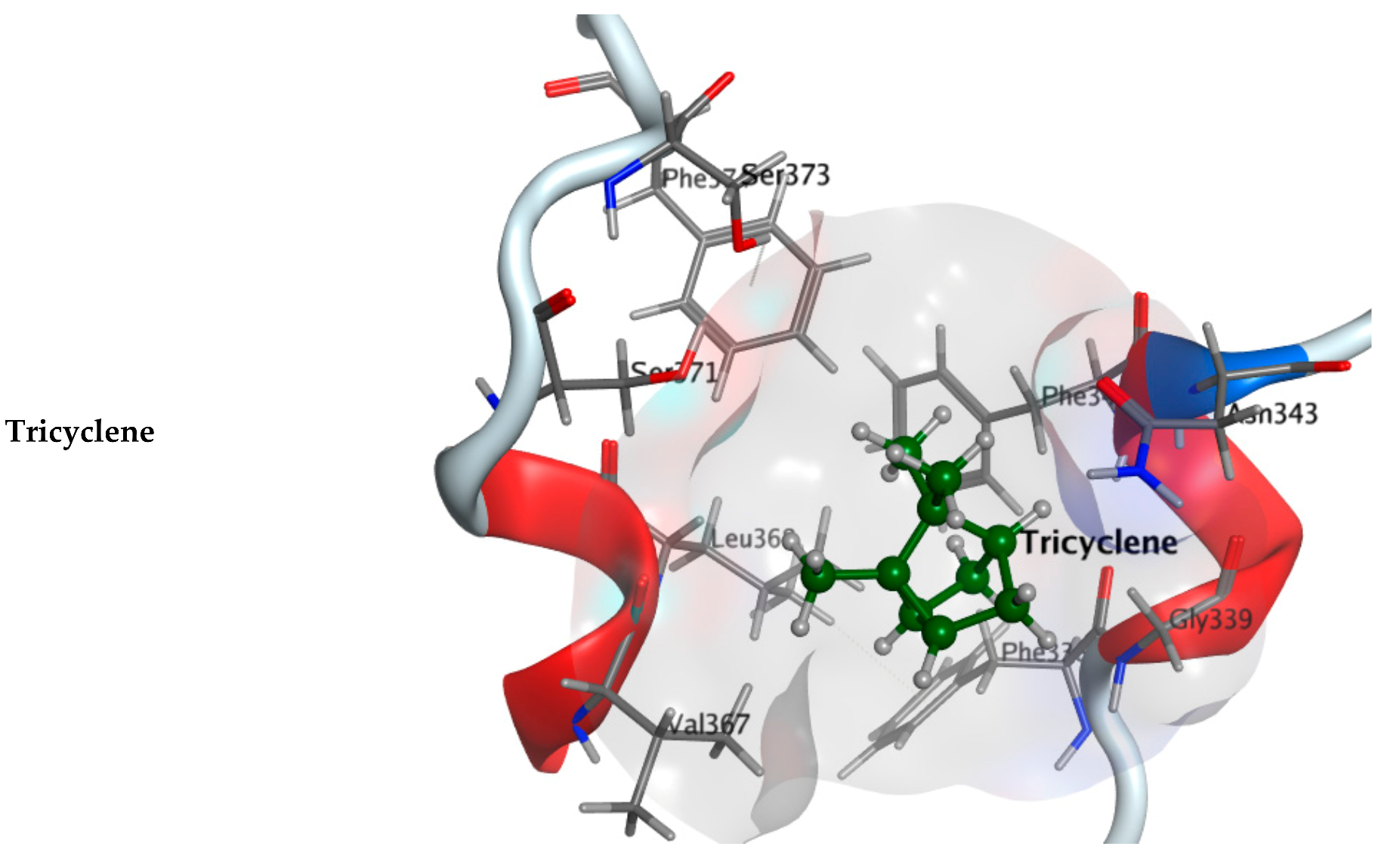
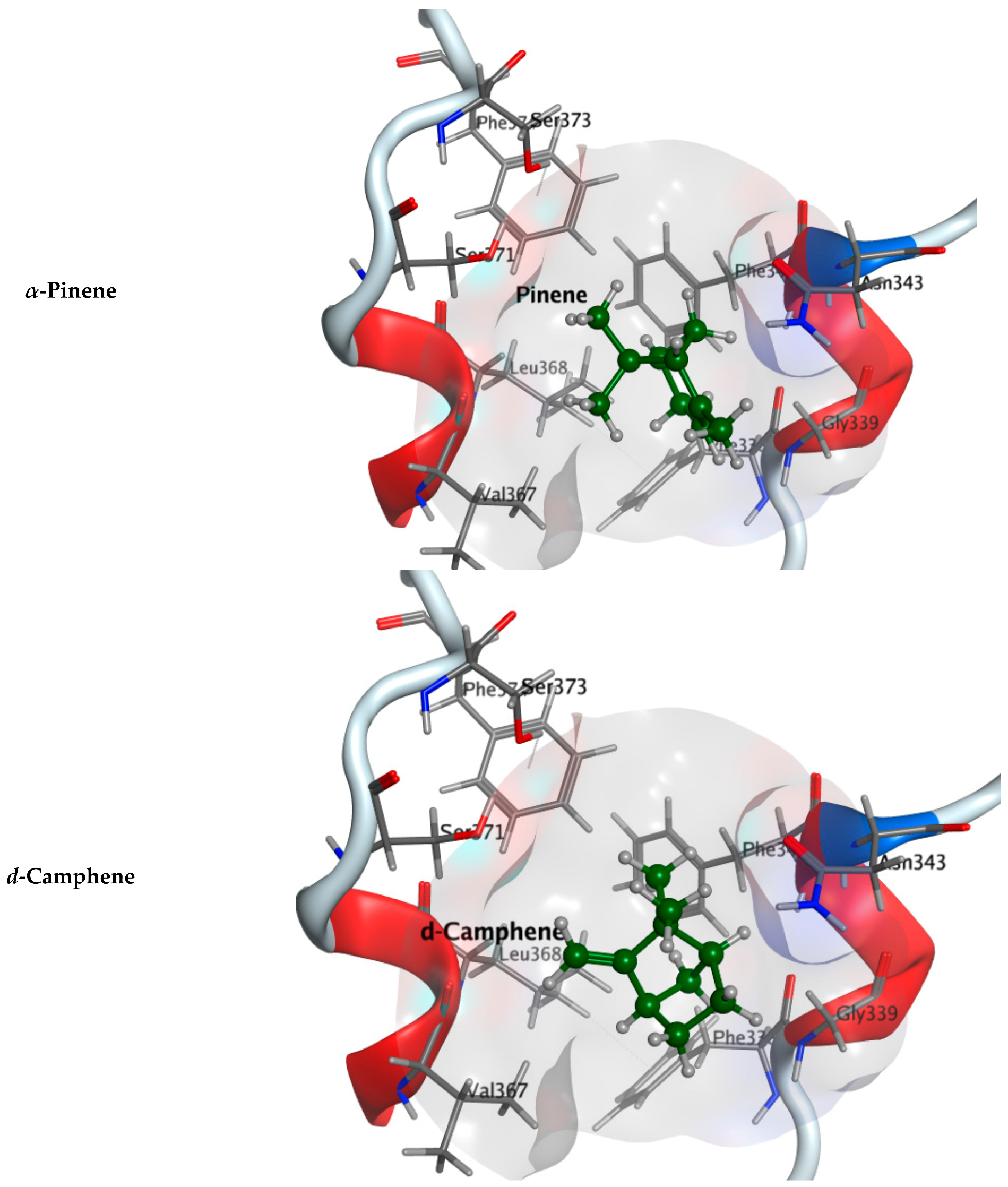
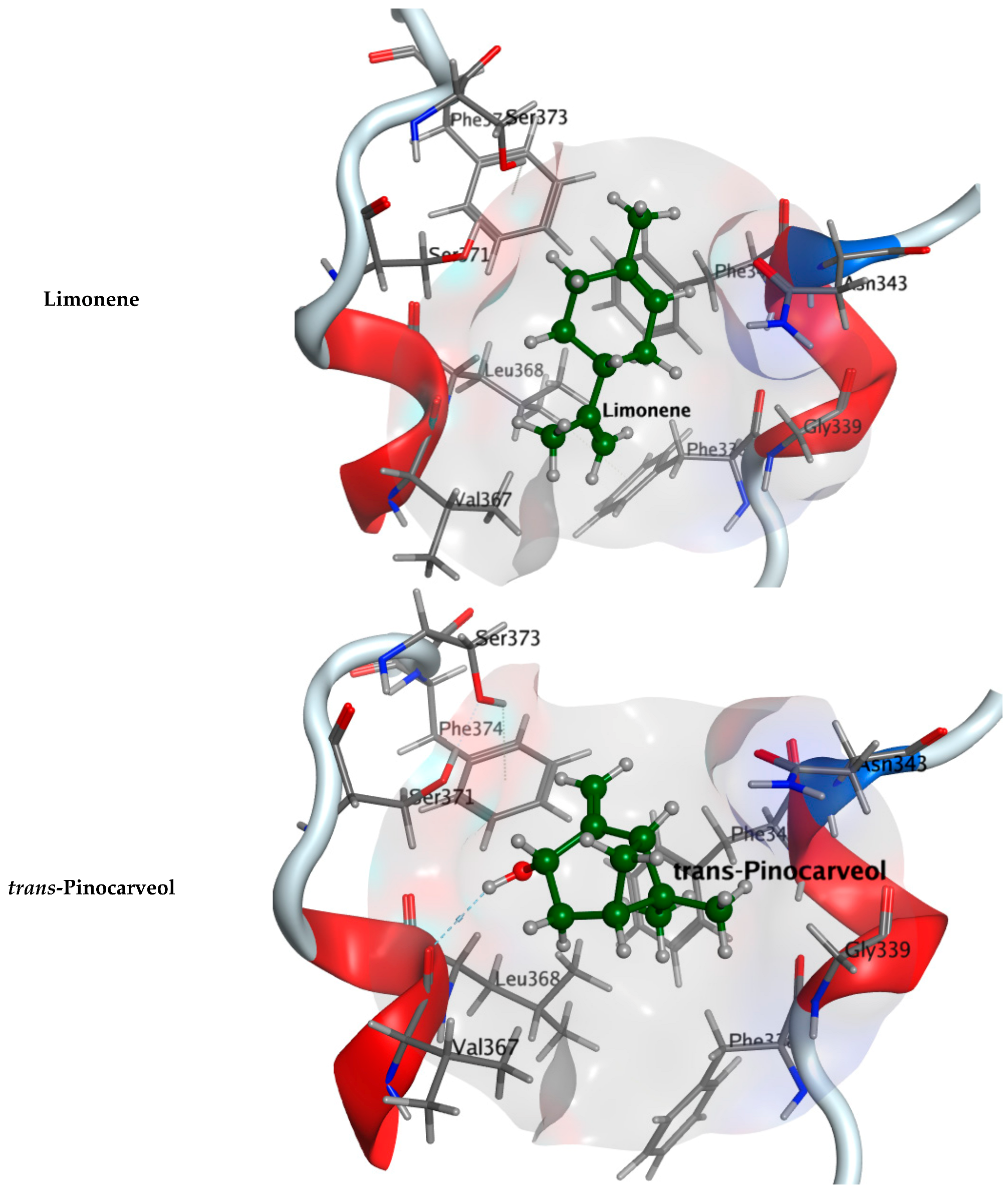
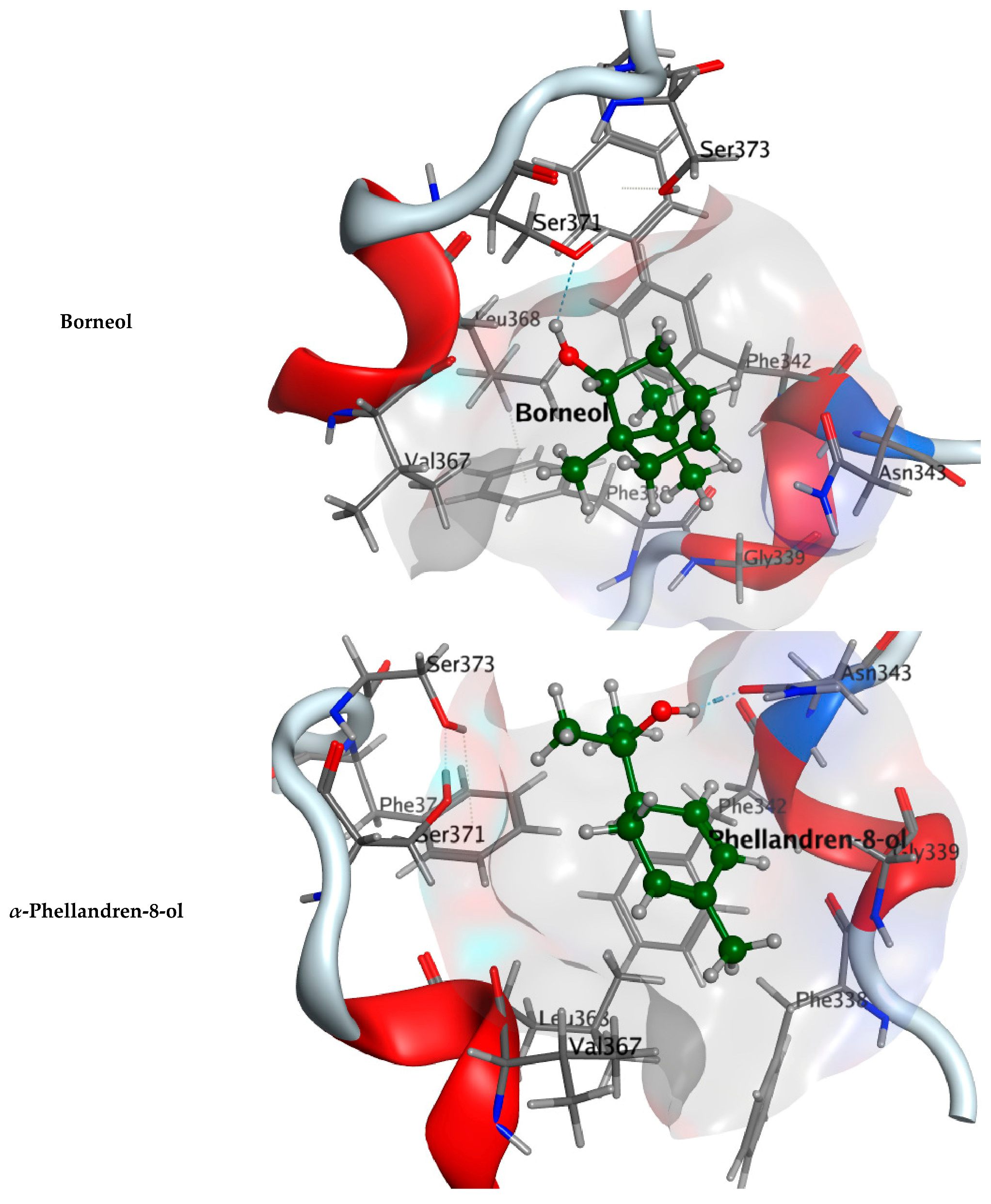
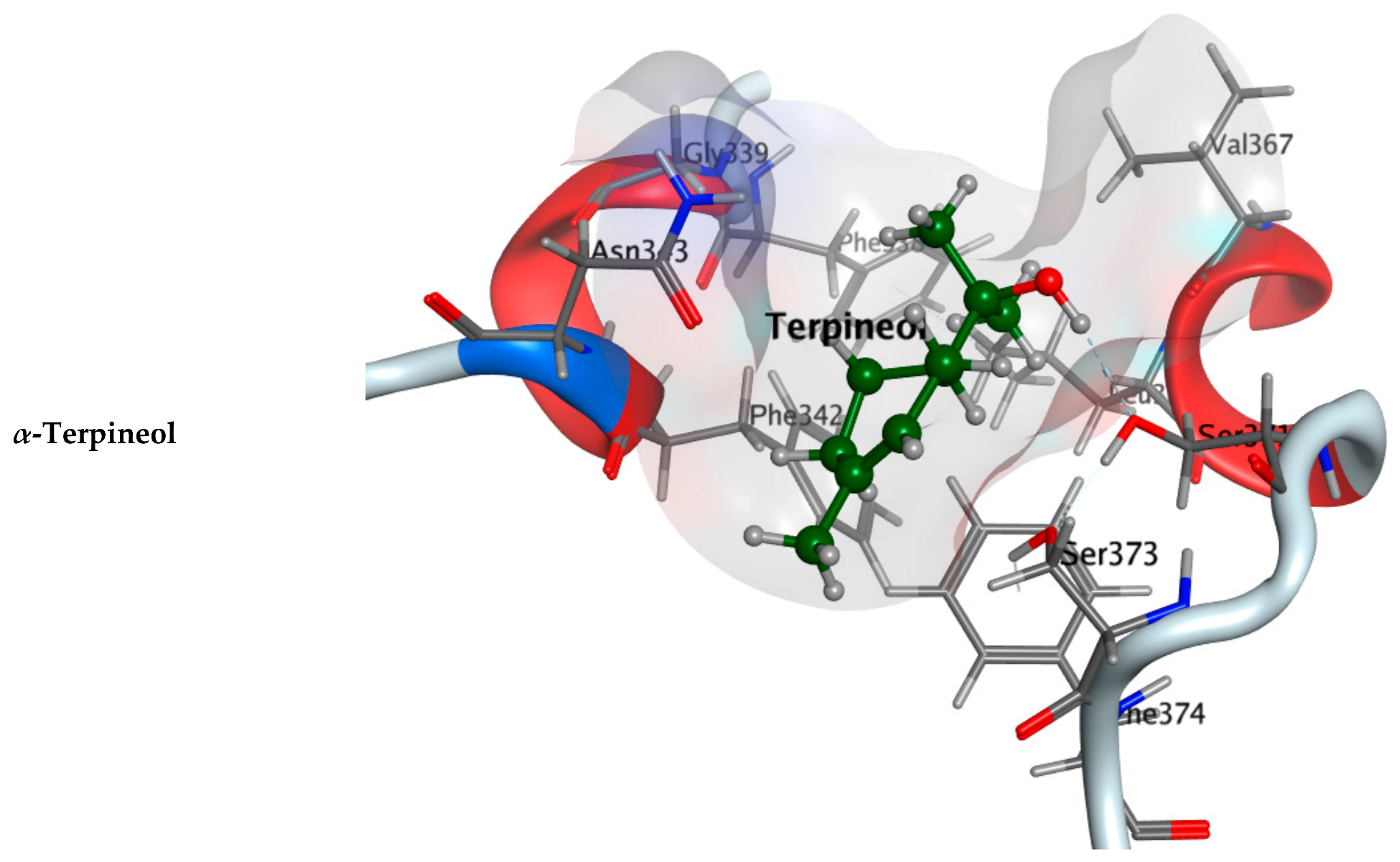
2.2.1. Molecular Docking Study
2.2.2. Flexible Alignments
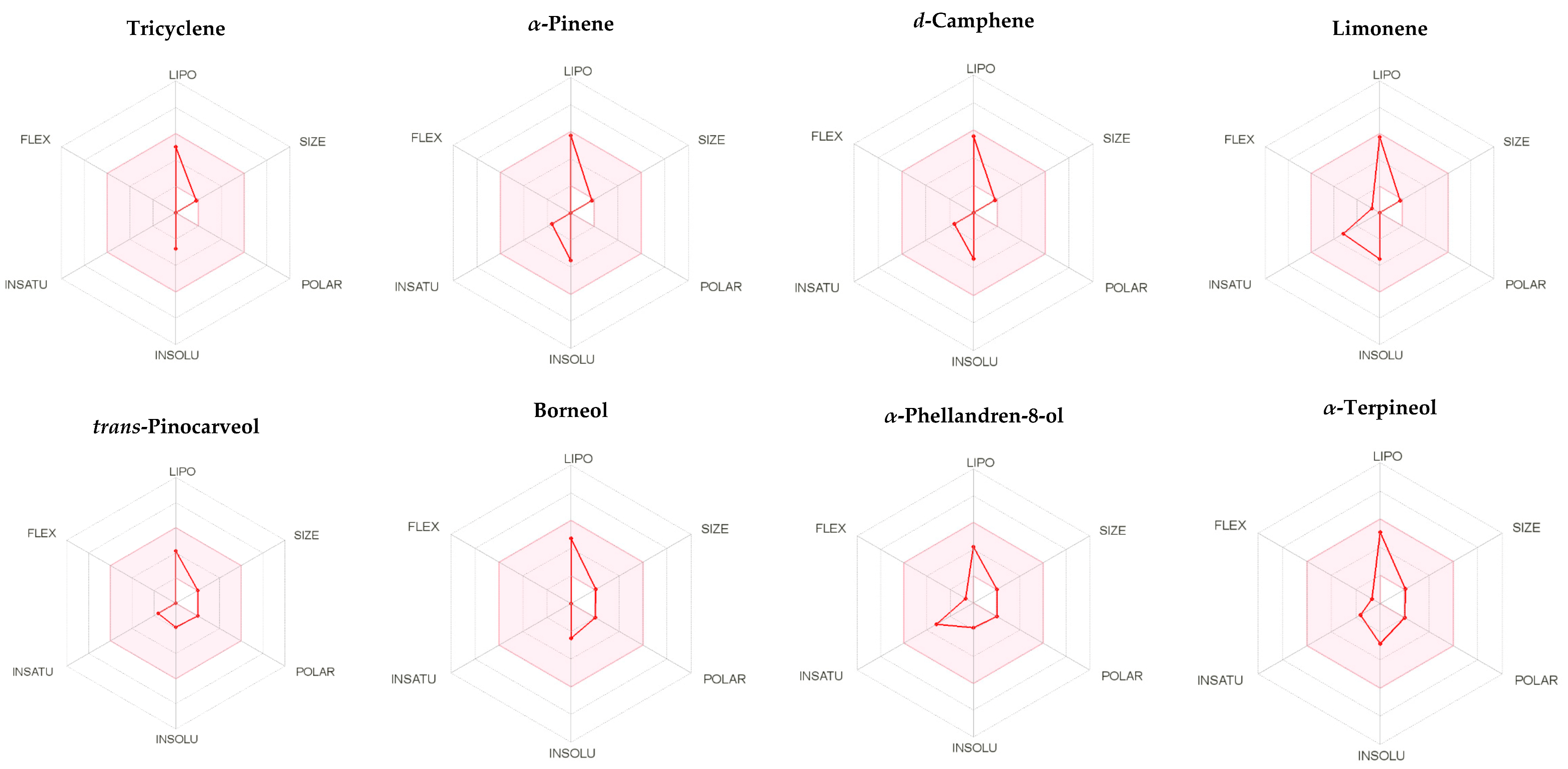
2.3. ADME Assessment of the Major Components of A. robusta Bark EO
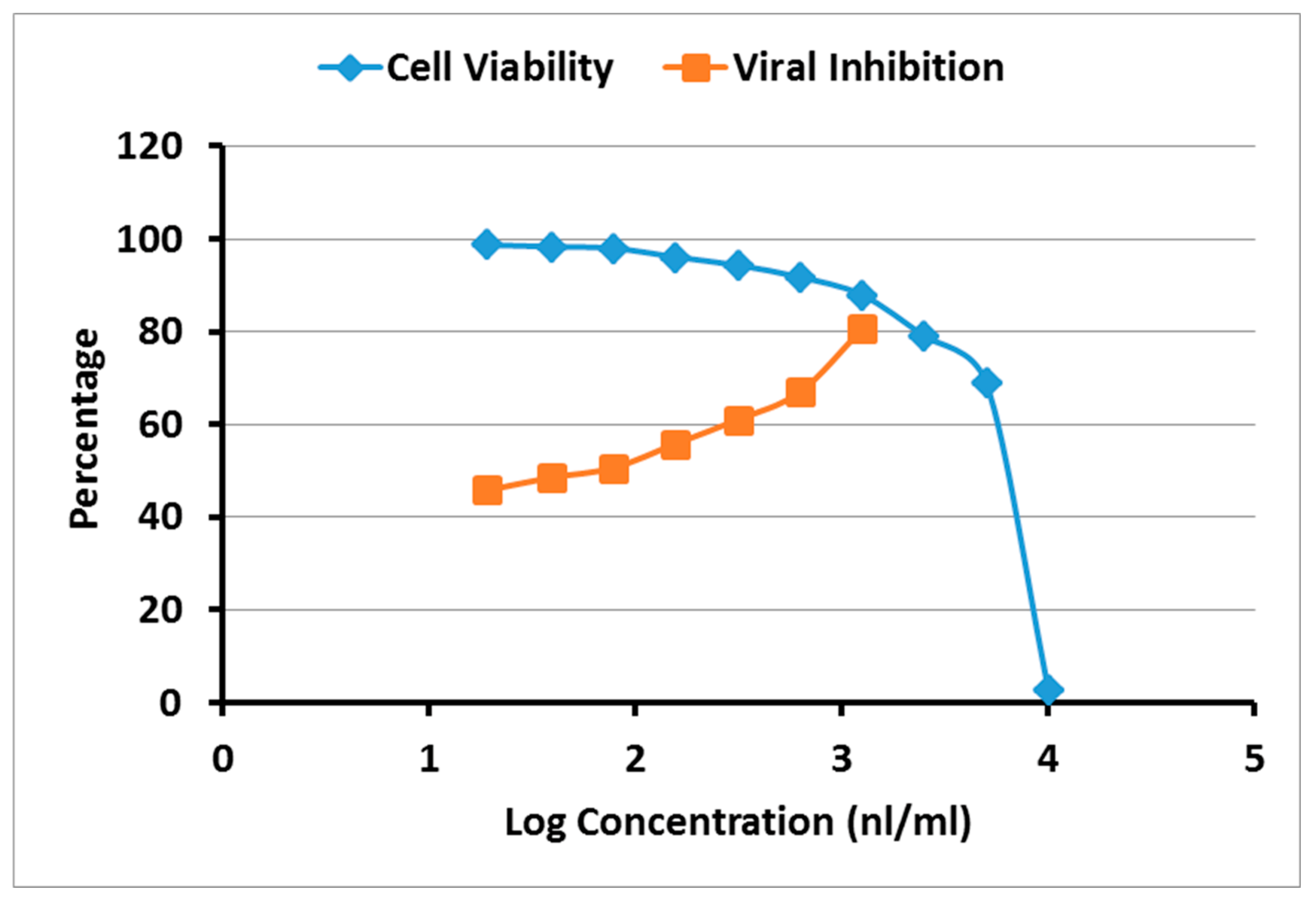
2.4. Cytotoxicity and Antiviral Activity of A. robusta Bark EO
3. Discussion
4. Material and Methods
4.1. Plant Material
4.2. Isolation of Bark EO
4.3. GC-FID and GC-MS Analyses
4.4. Molecular Modelling and In Silico Study
4.4.1. Molecular Docking Study
4.4.2. Protein Preparation
4.4.3. Ligand Preparation
4.4.4. Flexible Alignments
4.5. ADME Assessment of the Major Components of A. robusta Bark EO
4.6. MTT Cytotoxicity Assay
4.7. Inhibitory Concentration 50 (IC50) Determination
4.8. Calculation of Selectivity Index
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, O.V.; Rocha, G.B.; Paluch, A.S.; Costa, L.T. Repurposing approved drugs as inhibitors of SARS-CoV-2 S-protein from molecular modeling and virtual screening. J. Biomol. Struct. Dyn. 2021, 39, 3924–3933. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wu, T.; Liu, Q.; Yang, Z. The SARS-CoV-2 outbreak: What we know. Int. J. Infect. Dis. 2020, 94, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, S.; Zarbock, A. Coronaviruses and SARS-CoV-2: A brief overview. Anesth. Analg. 2020, 131, 93–96. [Google Scholar] [CrossRef]
- Lau, S.K.; Poon, R.W.; Wong, B.H.; Wang, M.; Huang, Y.; Xu, H.; Guo, R.; Li, K.S.; Gao, K.; Chan, K.-H. Coexistence of different genotypes in the same bat and serological characterization of Rousettus bat coronavirus HKU9 belonging to a novel Betacoronavirus subgroup. J. Virol. 2010, 84, 11385–11394. [Google Scholar] [CrossRef] [Green Version]
- Valencia, D.N. Brief review on COVID-19: The 2020 pandemic caused by SARS-CoV-2. Cureus 2020, 12, e7386. [Google Scholar] [CrossRef] [Green Version]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, P.; Wang, J.; Feng, J.; Zhou, H.; Li, X.; Zhong, W.; Hao, P. Evolution of the novel coronavirus from the ongoing Wuhan outbreak and modeling of its spike protein for risk of human transmission. Sci. China Life Sci. 2020, 63, 457–460. [Google Scholar] [CrossRef] [Green Version]
- Fehr, A.R.; Perlman, S. (Eds.) Coronaviruses: An Overview of Their Replication and Pathogenesis; Humana Press: New York, NY, USA, 2015; Volume 1282, pp. 1–23. [Google Scholar]
- Cui, J.; Li, F.; Shi, Z.-L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Fang, L.; Yang, Z.; Xu, S.; Lv, M.; Sun, Z.; Chen, J.; Wang, D.; Gao, J.; Xiao, S. Identification of novel proteolytically inactive mutations in coronavirus 3C-like protease using a combined approach. FASEB J. 2019, 33, 14575–14587. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Wan, Y.; Luo, C.; Ye, G.; Geng, Q.; Auerbach, A.; Li, F. Cell entry mechanisms of SARS-CoV-2. Proc. Natl. Acad. Sci. USA 2020, 117, 11727–11734. [Google Scholar] [CrossRef]
- Zu, Z.Y.; Jiang, M.D.; Xu, P.P.; Chen, W.; Ni, Q.Q.; Lu, G.M.; Zhang, L.J. Coronavirus disease 2019 (COVID-19): A perspective from China. Radiology 2020, 296, E15–E25. [Google Scholar] [CrossRef] [Green Version]
- Rossi, G.A.; Sacco, O.; Mancino, E.; Cristiani, L.; Midulla, F. Differences and similarities between SARS-CoV and SARS-CoV-2: Spike receptor-binding domain recognition and host cell infection with support of cellular serine proteases. Infection 2020, 48, 665–669. [Google Scholar] [CrossRef]
- Heald-Sargent, T.; Gallagher, T. Ready, set, fuse! The coronavirus spike protein and acquisition of fusion competence. Viruses 2012, 4, 557–580. [Google Scholar] [CrossRef] [Green Version]
- Follis, K.E.; York, J.; Nunberg, J.H. Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell–cell fusion but does not affect virion entry. Virology 2006, 350, 358–369. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zhou, Q.; Li, Y.; Garner, L.V.; Watkins, S.P.; Carter, L.J.; Smoot, J.; Gregg, A.C.; Daniels, A.D.; Jervey, S. Research and Development on Therapeutic Agents and Vaccines for COVID-19 and Related Human Coronavirus Diseases; ACS Publications: Washington, DC, USA, 2020. [Google Scholar]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C. Structure of M pro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Zhao, Y.; Cao, L.; Wang, T.; Sun, Q.; Ming, Z.; Zhang, L. Structure of the RNA-dependent RNA polymerase from COVID-19 virus. Science 2020, 368, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Anirudhan, V.; Du, R.; Cui, Q.; Rong, L. RNA-dependent RNA polymerase of SARS-CoV-2 as a therapeutic target. J. Med. Virol. 2021, 93, 300–310. [Google Scholar] [CrossRef]
- Allam, A.E.; Amen, Y.; Ashour, A.; Assaf, H.K.; Hassan, H.A.; Abdel-Rahman, I.M.; Sayed, A.M.; Shimizu, K. In silico study of natural compounds from sesame against COVID-19 by targeting M pro, PL pro and RdRp. RSC Adv. 2021, 11, 22398–22408. [Google Scholar] [CrossRef]
- Williamson, G.; Kerimi, A. Testing of natural products in clinical trials targeting the SARS-CoV-2 (COVID-19) viral spike protein-angiotensin converting enzyme-2 (ACE2) interaction. Biochem. Pharmacol. 2020, 178, 114123. [Google Scholar] [CrossRef]
- Ma, L.; Yao, L. Antiviral effects of plant-derived essential oils and their components: An updated review. Molecules 2020, 25, 2627. [Google Scholar] [CrossRef]
- Wink, M. Potential of DNA intercalating alkaloids and other plant secondary metabolites against SARS-CoV-2 causing COVID-19. Diversity 2020, 12, 175. [Google Scholar] [CrossRef]
- Schnitzler, P.; Astani, A.; Reichling, J. Antiviral effects of plant-derived essential oils and pure oil components. In Lipids and Essential Oils; Wiley: Hoboken, NJ, USA, 2011; Volume 239. [Google Scholar]
- Asif, M.; Saleem, M.; Saadullah, M.; Yaseen, H.S.; Al Zarzour, R. COVID-19 and therapy with essential oils having antiviral, anti-inflammatory, and immunomodulatory properties. Inflammopharmacology 2020, 28, 1153–1161. [Google Scholar] [CrossRef]
- Boland, D.J.; Brooker, M.I.H.; Chippendale, G.; Hall, N.; Hyland, B.; Johnston, R.; Kleinig, D.; McDonald, M.; Turner, J. Forest Trees of Australia; CSIRO Publishing: Melbourne, Austrilia, 2006. [Google Scholar]
- Williams, C. Medicinal Plants in Australia Volume 2: Gums, Resins, Tannin and Essential Oils; Rosenberg Publishing: Sydney, Austrilia, 2011; Volume 2. [Google Scholar]
- Boysen, B.M.; Evans, M.N.; Baker, P.J. δ 18O in the Tropical Conifer Agathis robusta Records ENSO-Related Precipitation Variations. PLoS ONE 2014, 9, e102336. [Google Scholar] [CrossRef] [Green Version]
- Farjon, A. A Handbook of the World’s Conifers (2 Vols.); Brill: Leiden, The Netherlands, 2010; Volume 1. [Google Scholar]
- Baker, R.T.; Smith, H.G. A Research on the Pines of Australia; WA Gullick, Government Printer: Sydney, Australia, 1910. [Google Scholar]
- Smith, H.G. Recent Work on the Australian Pines; Vacher & Sons: London, UK, 1911. [Google Scholar]
- Brophy, J.J.; Goldsack, R.J.; Wu, M.Z.; Fookes, C.J.; Forster, P.I. The steam volatile oil of Wollemia nobilis and its comparison with other members of the Araucariaceae (Agathis and Araucaria). Biochem. Syst. Ecol. 2000, 28, 563–578. [Google Scholar] [CrossRef]
- Verma, R.S.; Padalia, R.C.; Goswami, P.; Verma, S.K.; Chauhan, A.; Darokar, M.P. Chemical composition and antibacterial activity of the essential oil of Kauri Pine [Agathis robusta (C. Moore ex F. Muell.) FM Bailey] from India. J. Wood Chem. Technol. 2016, 36, 270–277. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456. [Google Scholar]
- Astani, A.; Schnitzler, P. Antiviral activity of monoterpenes beta-pinene and limonene against herpes simplex virus in vitro. Iran. J. Microbiol. 2014, 6, 149–155. [Google Scholar]
- Astani, A.; Reichling, J.; Schnitzler, P. Comparative study on the antiviral activity of selected monoterpenes derived from essential oils. Phytother. Res. PTR 2010, 24, 673–679. [Google Scholar] [CrossRef]
- Reichling, J. Antiviral and Virucidal Properties of Essential Oils and Isolated Compounds—A Scientific Approach. Planta Med. 2021. [Google Scholar] [CrossRef]
- Sobrinho, A.C.N.; de Morais, S.M.; Marinho, M.M.; de Souza, N.V.; Lima, D.M. Antiviral activity on the Zika virus and larvicidal activity on the Aedes spp. of Lippia alba essential oil and β-caryophyllene. Ind. Crops Prod. 2021, 162, 113281. [Google Scholar] [CrossRef]
- Sharma, A.D. Eucalyptol (1, 8 cineole) from eucalyptus essential oil a potential inhibitor of COVID 19 corona virus infection by molecular docking studies. Preprints 2020, 2020030455. [Google Scholar] [CrossRef] [Green Version]
- Thuy, B.T.P.; My, T.T.A.; Hai, N.T.T.; Hieu, L.T.; Hoa, T.T.; Thi Phuong Loan, H.; Triet, N.T.; Anh, T.T.V.; Quy, P.T.; Tat, P.V. Investigation into SARS-CoV-2 resistance of compounds in garlic essential oil. ACS Omega 2020, 5, 8312–8320. [Google Scholar] [CrossRef] [PubMed]
- Torres Neto, L.; Monteiro, M.L.G.; Galvan, D.; Conte-Junior, C.A. An Evaluation of the Potential of Essential Oils against SARS-CoV-2 from in silico Studies through the Systematic Review Using a Chemometric Approach. Pharmaceuticals 2021, 14, 1138. [Google Scholar] [CrossRef]
- Mieres-Castro, D.; Ahmar, S.; Shabbir, R.; Mora-Poblete, F. Antiviral Activities of Eucalyptus Essential Oils: Their Effectiveness as Therapeutic Targets against Human Viruses. Pharmaceuticals 2021, 14, 1210. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wu, N.; Zu, Y.; Fu, Y. Comparative anti-infectious bronchitis virus (IBV) activity of (-)-pinene: Effect on nucleocapsid (N) protein. Molecules 2011, 16, 1044–1054. [Google Scholar] [CrossRef] [PubMed]
- Meeran, M.N.; Seenipandi, A.; Javed, H.; Sharma, C.; Hashiesh, H.M.; Goyal, S.N.; Jha, N.K.; Ojha, S. Can limonene be a possible candidate for evaluation as an agent or adjuvant against infection, immunity, and inflammation in COVID-19? Heliyon 2021, 7, e05703. [Google Scholar] [CrossRef] [PubMed]
- Tshibangu, D.S.; Matondo, A.; Lengbiye, E.M.; Inkoto, C.L.; Ngoyi, E.M.; Kabengele, C.N.; Bongo, G.N.; Gbolo, B.Z.; Kilembe, J.T.; Mwanangombo, D.T. Possible effect of aromatic plants and essential oils against COVID-19: Review of their antiviral activity. J. Complement. Altern. Med. Res. 2020, 11, 10–22. [Google Scholar] [CrossRef]
- Hachlafi, N.E.; Aanniz, T.; Menyiy, N.E.; Baaboua, A.E.; Omari, N.E.; Balahbib, A.; Shariati, M.A.; Zengin, G.; Fikri-Benbrahim, K.; Bouyahya, A. In Vitro and in vivo biological investigations of camphene and its mechanism insights: A review. Food Rev. Int. 2021, 37, 1–28. [Google Scholar] [CrossRef]
- Sokolova, A.; Yarovaya, O.; Semenova, M.; Shtro, A.; Orshanskaya, I.; Zarubaev, V.; Salakhutdinov, N. Synthesis and In Vitro study of novel borneol derivatives as potent inhibitors of the influenza A virus. Med. Chem. Comm. 2017, 8, 960–963. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, A.S.; Kovaleva, K.S.; Yarovaya, O.I.; Bormotov, N.I.; Shishkina, L.N.; Serova, O.A.; Sergeev, A.A.; Agafonov, A.P.; Maksuytov, R.A.; Salakhutdinov, N.F. (+)-Camphor and (−)-borneol derivatives as potential anti-orthopoxvirus agents. Arch. Pharm. 2021, 354, 2100038. [Google Scholar] [CrossRef]
- Verma, S.; Twilley, D.; Esmear, T.; Oosthuizen, C.B.; Reid, A.-M.; Nel, M.; Lall, N. Anti-SARS-CoV natural products with the potential to inhibit SARS-CoV-2 (COVID-19). Front. Pharmacol. 2020, 11, 1514. [Google Scholar] [CrossRef]
- Indrayanto, G.; Putra, G.S.; Suhud, F. Validation of in-vitro bioassay methods: Application in herbal drug research. Profiles Drug Subst. Excip. Relat. Methodol. 2021, 46, 273–307. [Google Scholar]
- Perveen, K.; Bokhari, N.A.; Siddique, I.; Al-Rashid, S.A. Antifungal activity of essential oil of Commiphora molmol Oleo Gum Resin. J. Essent. Oil Bear. Plants 2018, 21, 667–673. [Google Scholar] [CrossRef]
- Bahmanzadegan, A.; Rowshan, V.; Saharkhiz, M.J. Essential oil composition of Myrtus communis L. under different storage conditions. J. Essent. Oil Bear. Plants 2015, 18, 1467–1475. [Google Scholar] [CrossRef]
- Berka-Zougali, B.; Ferhat, M.-A.; Hassani, A.; Chemat, F.; Allaf, K.S. Comparative study of essential oils extracted from Algerian Myrtus communis L. leaves using microwaves and hydrodistillation. Int. J. Mol. Sci. 2012, 13, 4673–4695. [Google Scholar] [CrossRef] [Green Version]
- Hendel, N.; Napoli, E.; Sarri, M.; Saija, A.; Cristani, M.; Nostro, A.; Ginestra, G.; Ruberto, G. Essential oil from aerial parts of wild Algerian rosemary: Screening of chemical composition, antimicrobial and antioxidant activities. J. Essent. Oil Bear. Plants 2019, 22, 1–17. [Google Scholar] [CrossRef]
- Environment, M.O.; Chemical Computing Group Inc. Montreal Quebec Canada. Available online: http://www.chemcomp.com (accessed on 2 January 2019).
- Wu, Y.; Wang, F.; Shen, C.; Peng, W.; Li, D.; Zhao, C.; Li, Z.; Li, S.; Bi, Y.; Yang, Y. A noncompeting pair of human neutralizing antibodies block COVID-19 virus binding to its receptor ACE2. Science 2020, 368, 1274–1278. [Google Scholar] [CrossRef]
- Yin, W.; Luan, X.; Li, Z.; Zhou, Z.; Wang, Q.; Gao, M.; Wang, X.; Zhou, F.; Shi, J.; You, E. Structural basis for inhibition of the SARS-CoV-2 RNA polymerase by suramin. Nat. Struct. Mol. Biol. 2021, 28, 319–325. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Mostafa, A.; Kandeil, A.; AMM Elshaier, Y.; Kutkat, O.; Moatasim, Y.; Rashad, A.A.; Shehata, M.; Gomaa, M.R.; Mahrous, N.; Mahmoud, S.H. FDA-approved drugs with potent In Vitro antiviral activity against severe acute respiratory syndrome coronavirus 2. Pharmaceuticals 2020, 13, 443. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name a | Rt b | RI c | Reported RI d | Area Percentage |
|---|---|---|---|---|---|
| 1 | Tricyclene | 7.509 | 927 | 921 | 11.89 |
| 2 | α-Pinene | 7.983 | 940 | 932 | 19.49 |
| 3 | d-Camphene | 8.529 | 953 | 946 | 7.13 |
| 4 | 2,4-Thujadiene | 8.672 | 966 | 953 | 1.06 |
| 5 | L-β-Pinene | 9.57 | 983 | 974 | 2.36 |
| 6 | m-Cymene | 11.485 | 1030 | 1018 | 1.58 |
| 7 | Limonene | 11.707 | 1040 | 1029 | 9.37 |
| 8 | γ-Terpinene | 12.807 | 1068 | 1059 | 0.30 |
| 9 | α-Campholenal | 15.421 | 1140 | 1126 | 1.33 |
| 10 | trans-Pinocarveol | 15.988 | 1152 | 1135 | 4.95 |
| 11 | Camphor | 16.119 | 1162 | 1145 | 0.43 |
| 12 | Camphene hydrate | 16.447 | 1164 | 1146 | 0.73 |
| 13 | Trans-Pinocamphone | 16.642 | 1170 | 1158 | 0.66 |
| 14 | Pinocarvone | 16.72 | 1176 | 1160 | 1.40 |
| 15 | Isoborneol | 16.794 | 1177 | 1161 | 1.30 |
| 16 | Borneol | 17.156 | 1183 | 1165 | 2.32 |
| 17 | α-Phellandren-8-ol | 17.223 | 1186 | 1170 | 2.51 |
| 18 | cis-Verbenol | 17.351 | 1189 | 1172 | 0.29 |
| 19 | L-terpinen-4-ol | 17.471 | 1191 | 1174 | 1.66 |
| 20 | Myrtenal | 17.957 | 1204 | 1195 | 1.41 |
| 21 | α-Terpineol | 18.125 | 1208 | 1186 | 9.59 |
| 22 | Verbenone | 18.472 | 1214 | 1205 | 1.93 |
| 23 | trans-Carveol | 18.98 | 1229 | 1215 | 1.25 |
| 24 | (-)-Carvone | 19.767 | 1276 | 1239 | 1.39 |
| 25 | Bornyl acetate | 21.139 | 1304 | 1287 | 1.29 |
| 26 | α-Terpineol acetate | 23.272 | 1368 | 1346 | 0.72 |
| Total | 88.34 | ||||
| Oil Components Classified According to Common Structures | Serial Numbers of Compounds Belonging to Each Class | Area Percentage | Total Number of Compounds |
|---|---|---|---|
| Monoterpenes | 1–26 | 88.37 | 26 |
| Oxygenation | |||
| Oxygenated | 9–26 | 35.16 | 18 |
| Alcohols | 10, 12, 15, 16, 17, 18, 19, 21, 23 | 24.60 | 9 |
| Aldehydes | 9, 20 | 2.74 | 2 |
| Esters | 25, 26 | 2.01 | 2 |
| Ketones | 11, 13, 14, 22, 24 | 5.81 | 5 |
| Cyclization | |||
| Acyclic | - | - | - |
| Monocyclic | 6, 7, 8, 9, 17, 19, 21, 23, 24, 26 | 29.7 | 10 |
| Bicyclic | 2, 3, 4, 5, 10, 11, 12, 13, 14, 15, 16, 18, 20, 22, 25 | 46.75 | 15 |
| Tricyclic | 1 | 11.89 | 1 |
| Component Name | COVID-19 Mpro (PDB: 6LU7) | COVID-19 RdRp (PDB: 7D4F) | COVID-19 RBD (PDB: 7BZ5) |
|---|---|---|---|
| Tricyclene | −4.6674 | −4.10620 | −4.0962 |
| α-Pinene | −4.7570 | −4.2618 | −3.8556 |
| d-Camphene | −4.4746 | −3.9284 | −4.0584 |
| Limonene | −4.7005 | −4.5654 | −3.9563 |
| trans-Pinocarveol | −4.59218 | −4.2639 | −4.1561 |
| Borneol | −4.1916 | −4.0815 | −3.9368 |
| α-Phellandren-8-ol | −4.9801 | −4.3839 | −4.0385 |
| α-Terpineol | −5.0752 | −4.7213 | −4.2190 |
| Co-crystallized ligand | −8.4596 | −7.7170 | −4.5304 |
| Component | Flexible Alignment | S kcal/mol |
|---|---|---|
| trans-Pinocarveol |  | −146.505341 |
| Borneol |  | −22.2655106 |
| α-Phellandren-8-ol |  | −48.4288139 |
| α-Terpineol |  | −51.053772 |
| Tricyclene | α-Pinene | d-Camphene | Limonene | Trans-Pinocarveol | Borneol | α-Phellandren-8-ol | α-Terpineol | |
|---|---|---|---|---|---|---|---|---|
| M.wt. | 136.23 | 136.23 | 136.23 | 136.23 | 152.23 | 154.25 | 152.23 | 154.25 |
| HBA | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 |
| HBD | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 |
| GI absorption | Low | Low | Low | Low | High | High | High | High |
| BBB permeant | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| P-gp substrate | No | No | No | No | No | No | No | No |
| CYP1A2 inhibitor | No | No | No | No | No | No | No | No |
| CYP2C19 inhibitor | No | No | No | No | No | No | No | No |
| CYP2C9 inhibitor | No | Yes | Yes | Yes | No | No | No | No |
| CYP2D6 inhibitor | No | No | No | No | No | No | No | No |
| CYP3A4 inhibitor | No | No | No | No | No | No | No | No |
| Lipinski’s violation | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| MLOGP | 4.43 | 4.29 | 4.29 | 3.27 | 2.30 | 2.45 | 2.20 | 2.30 |
| Xlogp3 | 3.24 | 4.48 | 4.22 | 4.57 | 1.79 | 2.72 | 1.81 | 3.39 |
| TPSA | 0 | 0 | 0 | 0 | 20.23 | 20.23 | 20.23 | 20.23 |
| Log S (ESOL), water solubility | −2.73 | −3.51 | −3.34 | −3.50 | −1.91 | −2.51 | −1.86 | −2.87 |
| Fraction Csp3 | 1 | 0.8 | 0.8 | 0.6 | 0.8 | 1 | 0.6 | 0.8 |
| Num. rotatable bonds | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, M.E.; Tawfeek, N.; Elbaramawi, S.S.; Fikry, E. Agathis robusta Bark Essential Oil Effectiveness against COVID-19: Chemical Composition, In Silico and In Vitro Approaches. Plants 2022, 11, 663. https://doi.org/10.3390/plants11050663
Mohamed ME, Tawfeek N, Elbaramawi SS, Fikry E. Agathis robusta Bark Essential Oil Effectiveness against COVID-19: Chemical Composition, In Silico and In Vitro Approaches. Plants. 2022; 11(5):663. https://doi.org/10.3390/plants11050663
Chicago/Turabian StyleMohamed, Maged E., Nora Tawfeek, Samar S. Elbaramawi, and Eman Fikry. 2022. "Agathis robusta Bark Essential Oil Effectiveness against COVID-19: Chemical Composition, In Silico and In Vitro Approaches" Plants 11, no. 5: 663. https://doi.org/10.3390/plants11050663