
Dual Beneficial Effects of α-Spinasterol Isolated from Aster pseudoglehnii on Glucose Uptake in Skeletal Muscle Cells and Glucose-Stimulated Insulin Secretion in Pancreatic β-Cells

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of α-Spinasterol
2.2. Effect of α-Spinasterol on Glucose Uptake in Skeletal Muscle Cells
2.3. Effect of α-Spinasterol on the Protein Expression of P-IRS-1, IRS-1, P-AMPK, AMPK, and GLUT-4
2.4. Effect of α-Spinasterol on GSIS
2.5. Effect of α-Spinasterol on the Protein Expression of P-IRS-2, IRS-2 (Ser731), PPARγ, and PDX-1
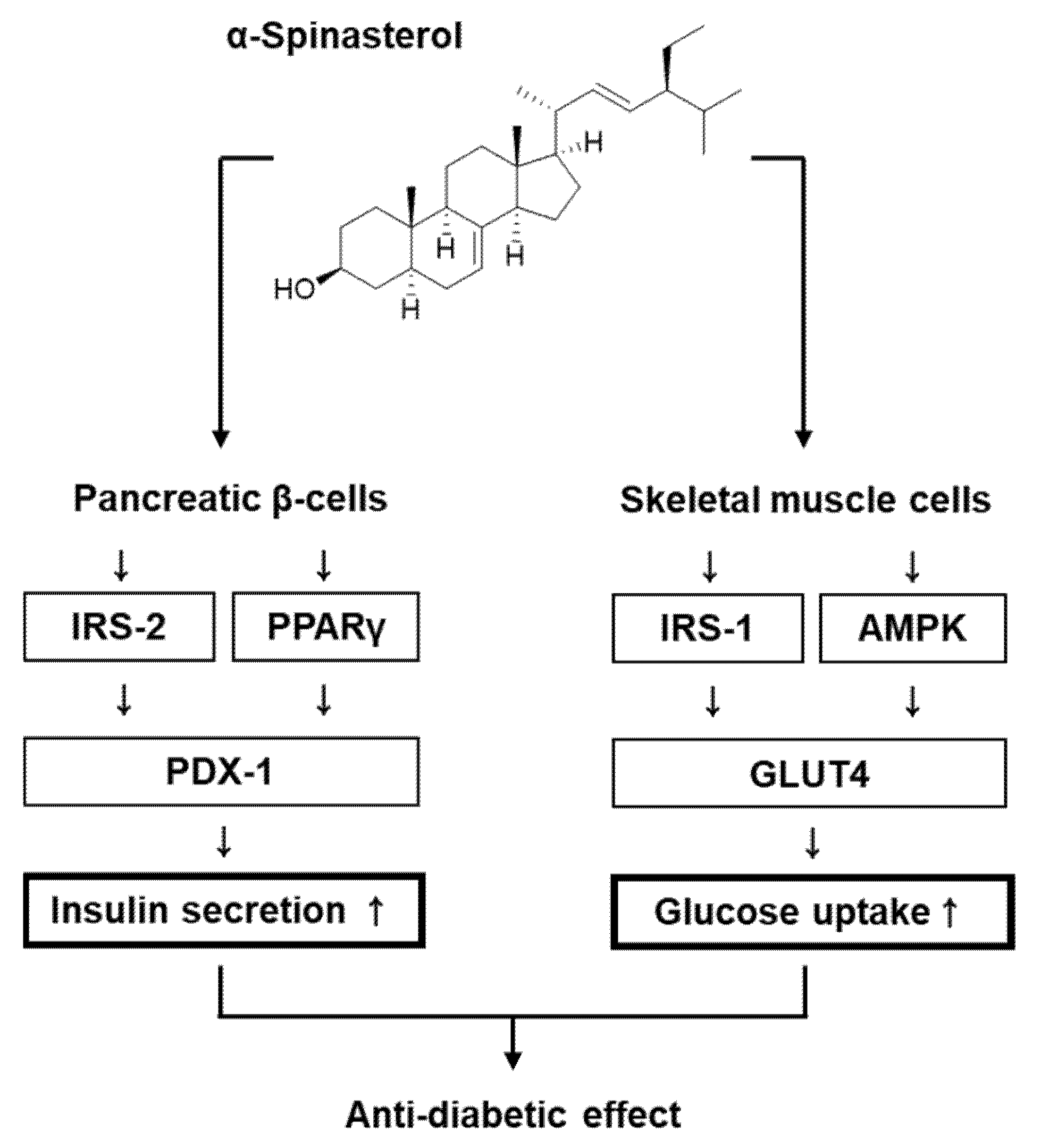
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Isolation
α-Spinasterol
4.4. Cell Culture and Chemicals
4.5. Cell Viability Assay
4.6. Glucose Uptake Assay
4.7. GSIS Assay
4.8. ADP/ATP Ratio Assay
4.9. Western Blot Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef]
- Teng, S.; Huang, P. The effect of type 2 diabetes mellitus and obesity on muscle progenitor cell function. Stem Cell Res. Ther. 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Soundharrajan, I.; Kim, D.H.; Srisesharam, S.; Kuppusamy, P.; Sivanesan, R.; Choi, K.C. Limonene promotes osteoblast differentiation and 2-deoxy-d-glucose uptake through p38MAPK and Akt signaling pathways in C2C12 skeletal muscle cells. Phytomedicine 2018, 45, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, A.; Pari, L. Myrtenal ameliorates hyperglycemia by enhancing GLUT2 through Akt in the skeletal muscle and liver of diabetic rats. Chem. Biol. Interact. 2016, 256, 161–166. [Google Scholar] [CrossRef]
- Zielińska-Błajet, M.; Feder-Kubis, J. Monoterpenes and their derivatives—Recent development in biological and medical applications. Int. J. Mol. Sci. 2020, 21, 7078. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Go, G.-W.; Imm, J.-Y. Promotion of glucose uptake in C2C12 myotubes by cereal flavone tricin and its underlying molecular mechanism. J. Agric. Food Chem. 2017, 65, 3819–3826. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.; Jones, L.C.; de Koning, E.; Hansen, B.C.; Matthews, D.R. Decreased insulin secretion in type 2 diabetes: A problem of cellular mass or function? Diabetes 2001, 50, S169. [Google Scholar] [CrossRef] [Green Version]
- Meda, P.; Schuit, F. Glucose-stimulated insulin secretion: The hierarchy of its multiple cellular and subcellular mechanisms. Diabetologia 2013, 56, 2552–2555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halperin, F.; Lopez, X.; Manning, R.; Kahn, C.R.; Kulkarni, R.N.; Goldfine, A.B. Insulin augmentation of glucose-stimulated insulin secretion is impaired in insulin-resistant humans. Diabetes 2012, 61, 301–309. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.; Hwang, B.S.; Choi, P.; Kim, T.; Kim, Y.; Song, B.G.; Yamabe, N.; Hwang, G.S.; Kang, K.S.; Ham, J. Hypoxylonol F Isolated from Annulohypoxylon annulatum improves insulin secretion by regulating pancreatic β-cell metabolism. Biomolecules 2019, 9, 335. [Google Scholar] [CrossRef] [Green Version]
- Park, E.-Y.; Kim, H.-J.; Kim, Y.-K.; Park, S.-U.; Choi, J.-E.; Cha, J.-Y.; Jun, H.-S. Increase in insulin secretion induced by panax ginseng berry extracts contributes to the amelioration of hyperglycemia in streptozotocininduced diabetic mice. J. Ginseng Res. 2012, 36, 153. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, P. Recent trends in therapeutic approaches for diabetes management: A comprehensive update. J Diabetes Res. 2015, 2015, 340838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, D. Illustrated Book of Korean Medicinal Herbs, 3rd ed.; Kyo-hak Publishing Co.: Seoul, Korea, 1998; Volume 107. [Google Scholar]
- Lee, H.-M.; Yang, G.; Ahn, T.-G.; Kim, M.-D.; Nugroho, A.; Park, H.-J.; Lee, K.-T.; Park, W.; An, H.-J. Antiadipogenic effects of Aster glehni extract: In vivo and in vitro effects. Evid.-Based Complementary Altern. Med. 2013, 2013, 859624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Bae, H.J.; Park, J.H.; Zhang, J.; Koo, B.; Lim, M.K.; Han, E.H.; Lee, S.H.; Jung, S.Y.; Lew, J.H. Aster glehni extract ameliorates scopolamine-induced cognitive impairment in mice. J. Med. Food 2019, 22, 685–695. [Google Scholar] [CrossRef]
- Zhang, L.-J.; Yang, X.-D.; Xu, L.-Z.; Zou, Z.-M.; Yang, S.-L. A new sterol glycoside from Securidaca inappendiculata. J. Asian Nat. Prod. Res. 2005, 7, 649–653. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.I.; Kim, K.J.; Choi, M.K.; Keum, K.S.; Lee, S.; Ahn, S.H.; Back, S.H.; Song, J.H.; Ju, Y.S.; Choi, B.K. α-Spinasterol isolated from the root of Phytolacca americana and its pharmacological property on diabetic nephropathy. Planta Med. 2004, 70, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y.; Shin, I.S.; Kyoung, H.; Seo, C.S.; Son, J.-K.; Shin, H.K. α-Spinasterol from Melandrium firmum attenuates benign prostatic hyperplasia in a rat model. Mol. Med. Rep. 2014, 9, 2362–2366. [Google Scholar] [CrossRef] [PubMed]
- Borges, F.R.; Silva, M.D.; Córdova, M.M.; Schambach, T.R.; Pizzolatti, M.G.; Santos, A.R. Anti-inflammatory action of hydroalcoholic extract, dichloromethane fraction and steroid α-spinasterol from Polygala sabulosa in LPS-induced peritonitis in mice. J. Ethnopharmacol. 2014, 151, 144–150. [Google Scholar] [CrossRef]
- Brusco, I.; Camponogara, C.; Carvalho, F.B.; Schetinger, M.R.C.; Oliveira, M.S.; Trevisan, G.; Ferreira, J.; Oliveira, S.M. α-Spinasterol: A COX inhibitor and a transient receptor potential vanilloid 1 antagonist presents an antinociceptive effect in clinically relevant models of pain in mice. Br. J. Pharmacol. 2017, 174, 4247–4262. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhou, J.; Wang, T.; Zhao, L.; Ye, G.; Shi, F.; Li, Y.; Tang, H.; Dong, Q.; Zhou, X. A novel method for synthesis of α-spinasterol and its antibacterial activities in combination with ceftiofur. Fitoterapia 2017, 119, 12–19. [Google Scholar] [CrossRef]
- Richter, E.A.; Hargreaves, M. Exercise, GLUT4, and skeletal muscle glucose uptake. Physiol. Rev. 2013, 93, 993–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, C.H.Y.; Hsu, C.-L.; Lin, T.-Y.; Chen, W.-T.; Wang, Y.-C.; Huang, H.-C.; Juan, H.-F. ZNF322A-mediated protein phosphorylation induces autophagosome formation through modulation of IRS1-AKT glucose uptake and HSP-elicited UPR in lung cancer. J. Biomed. Sci. 2020, 27, 75. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, F.M. ATP-sensitive potassium channelopathies: Focus on insulin secretion. J. Clin. Investig. 2005, 115, 2047–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seino, S. Cell signalling in insulin secretion: The molecular targets of ATP, cAMP and sulfonylurea. Diabetologia 2012, 55, 2096–2108. [Google Scholar] [CrossRef]
- Kusunoki, M.; Hayashi, M.; Shoji, T.; Uba, T.; Tanaka, H.; Sumi, C.; Matsuo, Y.; Hirota, K. Propofol inhibits stromatoxin-1-sensitive voltage-dependent K+ channels in pancreatic β-cells and enhances insulin secretion. PeerJ 2019, 7, e8157. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, L.; Li, Y.; Chen, H.; Sun, Z. Nifedipine protects INS-1 β-cell from high glucose-induced ER stress and apoptosis. Int. J. Mol. Sci. 2011, 12, 7569–7580. [Google Scholar] [CrossRef] [Green Version]
- Bardy, G.; Virsolvy, A.; Quignard, J.; Ravier, M.; Bertrand, G.; Dalle, S.; Cros, G.; Magous, R.; Richard, S.; Oiry, C. Quercetin induces insulin secretion by direct activation of L-type calcium channels in pancreatic beta cells. Br. J. Pharmacol. 2013, 169, 1102–1113. [Google Scholar] [CrossRef] [Green Version]
- Santini, E.; Fallahi, P.; Ferrari, S.M.; Masoni, A.; Antonelli, A.; Ferrannini, E. Effect of PPAR-γ activation and inhibition on glucose-stimulated insulin release in INS-1e cells. Diabetes 2004, 53, S79–S83. [Google Scholar] [CrossRef] [Green Version]
- Yajima, K.; Hirose, H.; Fujita, H.; Seto, Y.; Fujita, H.; Ukeda, K.; Miyashita, K.; Kawai, T.; Yamamoto, Y.; Ogawa, T. Combination therapy with PPARγ and PPARα agonists increases glucose-stimulated insulin secretion in db/db mice. Am. J. Physiol.-Endocrinol. Metab. 2003, 284, E966–E971. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Hwang, Y.-C.; Koo, S.-H.; Park, K.S.; Lee, M.-S.; Kim, K.-W.; Lee, M.-K. PPAR-γ activation increases insulin secretion through the up-regulation of the free fatty acid receptor GPR40 in pancreatic β-cells. PLoS ONE 2013, 8, e50128. [Google Scholar] [CrossRef] [Green Version]
- Brissova, M.; Shiota, M.; Nicholson, W.E.; Gannon, M.; Knobel, S.M.; Piston, D.W.; Wright, C.V.; Powers, A.C. Reduction in pancreatic transcription factor PDX-1 impairs glucose-stimulated insulin secretion. J. Biol. Chem. 2002, 277, 11225–11232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gauthier, B.R.; Wiederkehr, A.; Baquié, M.; Dai, C.; Powers, A.C.; Kerr-Conte, J.; Pattou, F.; MacDonald, R.J.; Ferrer, J.; Wollheim, C.B. PDX1 deficiency causes mitochondrial dysfunction and defective insulin secretion through TFAM suppression. Cell Metab. 2009, 10, 110–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K.; Iemitsu, M.; Aizawa, K.; Ajisaka, R. Testosterone and DHEA activate the glucose metabolism-related signaling pathway in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E961–E968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, G.; Allard, C.; Morford, J.J.; Xu, W.; Liu, S.; Molinas, A.J.; Butcher, S.M.; Fine, N.H.; Blandino-Rosano, M.; Sure, V.N. Androgen excess in pancreatic β cells and neurons predisposes female mice to type 2 diabetes. JCI Insight 2018, 3, e98607. [Google Scholar] [CrossRef]
- Wong, W.P.; Tiano, J.P.; Liu, S.; Hewitt, S.C.; Le May, C.; Dalle, S.; Katzenellenbogen, J.A.; Katzenellenbogen, B.S.; Korach, K.S.; Mauvais-Jarvis, F. Extranuclear estrogen receptor-α stimulates NeuroD1 binding to the insulin promoter and favors insulin synthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 13057–13062. [Google Scholar] [CrossRef] [Green Version]
- Yun, M.; Yi, Y.S. Regulatory roles of ginseng on inflammatory caspases, executioners of inflammasome activation. J. Ginseng Res. 2020, 44, 373–385. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Kim, J.-Y.; Kwon, H.C.; Kwon, J.; Jang, D.S.; Kang, K.S. Dual Beneficial Effects of α-Spinasterol Isolated from Aster pseudoglehnii on Glucose Uptake in Skeletal Muscle Cells and Glucose-Stimulated Insulin Secretion in Pancreatic β-Cells. Plants 2022, 11, 658. https://doi.org/10.3390/plants11050658
Lee D, Kim J-Y, Kwon HC, Kwon J, Jang DS, Kang KS. Dual Beneficial Effects of α-Spinasterol Isolated from Aster pseudoglehnii on Glucose Uptake in Skeletal Muscle Cells and Glucose-Stimulated Insulin Secretion in Pancreatic β-Cells. Plants. 2022; 11(5):658. https://doi.org/10.3390/plants11050658
Chicago/Turabian StyleLee, Dahae, Ji-Young Kim, Hak Cheol Kwon, Jaeyoung Kwon, Dae Sik Jang, and Ki Sung Kang. 2022. "Dual Beneficial Effects of α-Spinasterol Isolated from Aster pseudoglehnii on Glucose Uptake in Skeletal Muscle Cells and Glucose-Stimulated Insulin Secretion in Pancreatic β-Cells" Plants 11, no. 5: 658. https://doi.org/10.3390/plants11050658