
A Major Diplotaxis harra-Derived Bioflavonoid Glycoside as a Protective Agent against Chemically Induced Neurotoxicity and Parkinson’s Models; In Silico Target Prediction; and Biphasic HPTLC-Based Quantification

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. The Phytochemical and Antioxidant Evaluation of DHE, and Isolation of a Major Antioxidant Compound
2.1.1. The Total Phenolic and Total Flavonoid Contents (TPC and TFC), and the Antioxidant Activity of DHE
2.1.2. Isolation and Identification of IR3G
2.2. The Anti-Inflammatory, Neuroprotective, and Locomotion Deficit Inhibitory Activities
2.2.1. The In Vitro Anti-Inflammatory Activity of DHE and IR3G on iNOS and COX-2 Expression
2.2.2. The Protective Effect of IR3G against the In Vitro 6-OHDA-Induced Neurotoxicity
2.2.3. The Protective Effect of IR3G on the In Vivo 6-OHDA-Induced Deficits in Locomotor Activity
2.3. In Silico Study
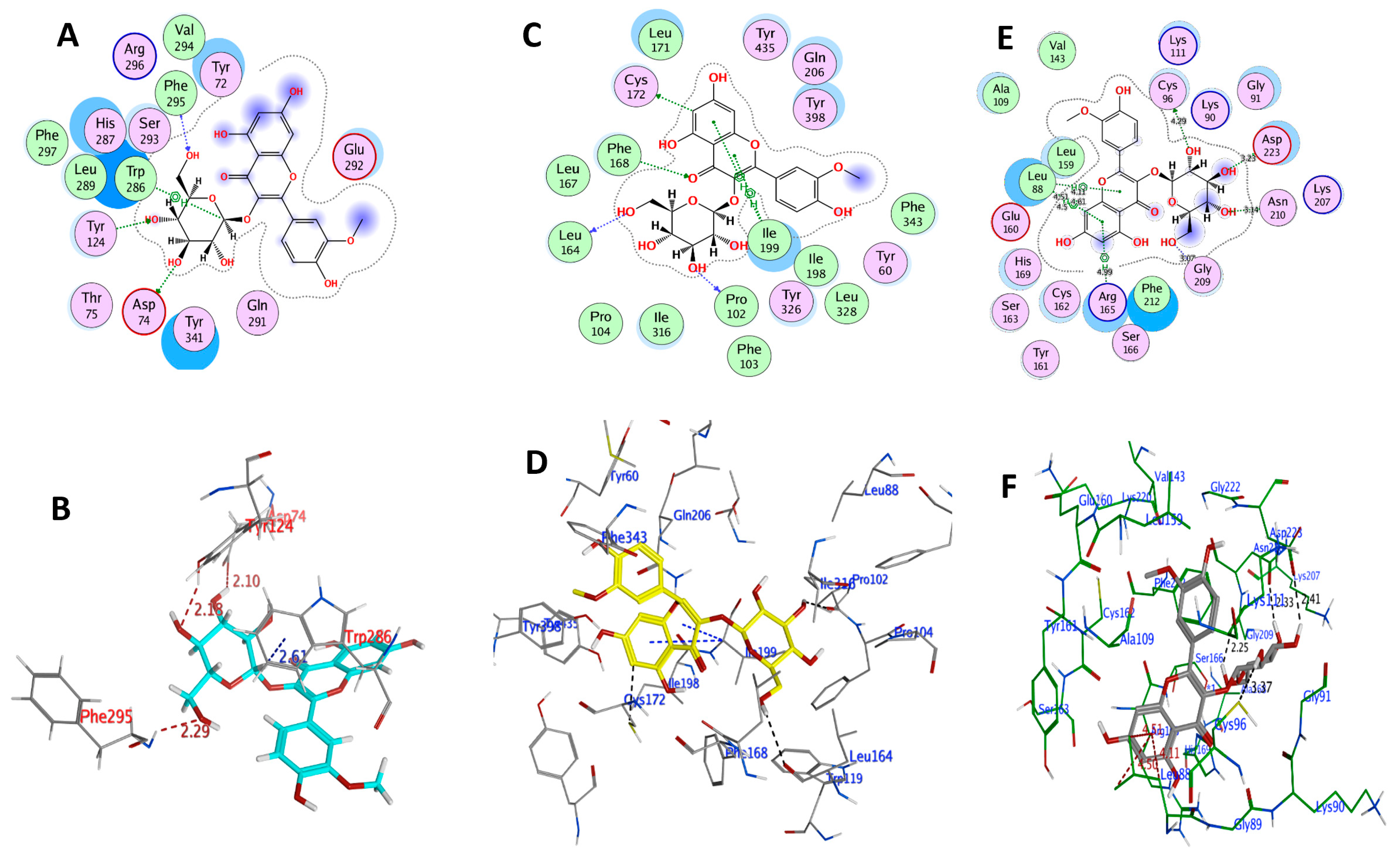
2.3.1. The Molecular Docking Study of IR3G and Its Aglycone IR
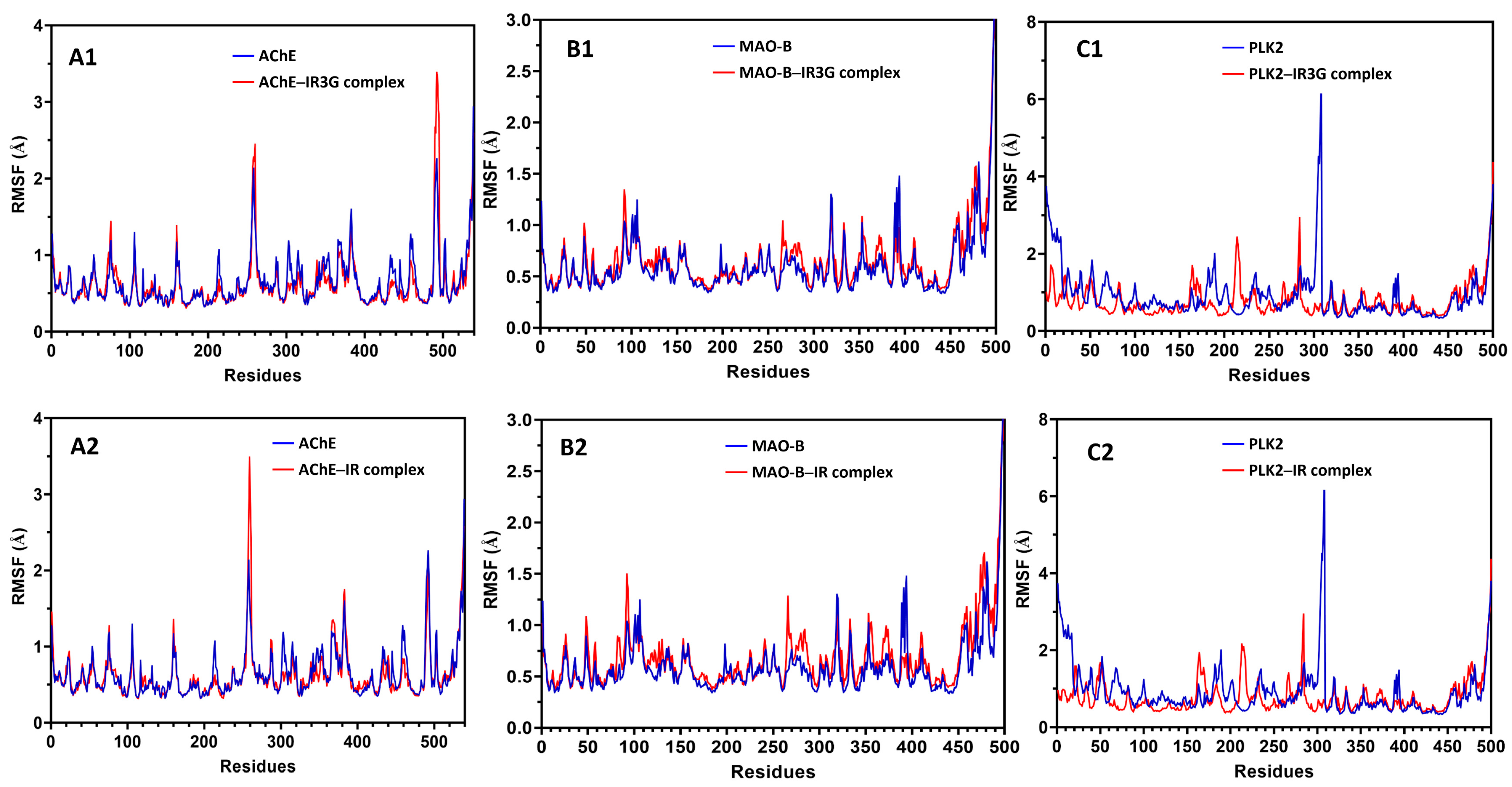
2.3.2. MD Simulation Study of Protein–Ligand Complexes
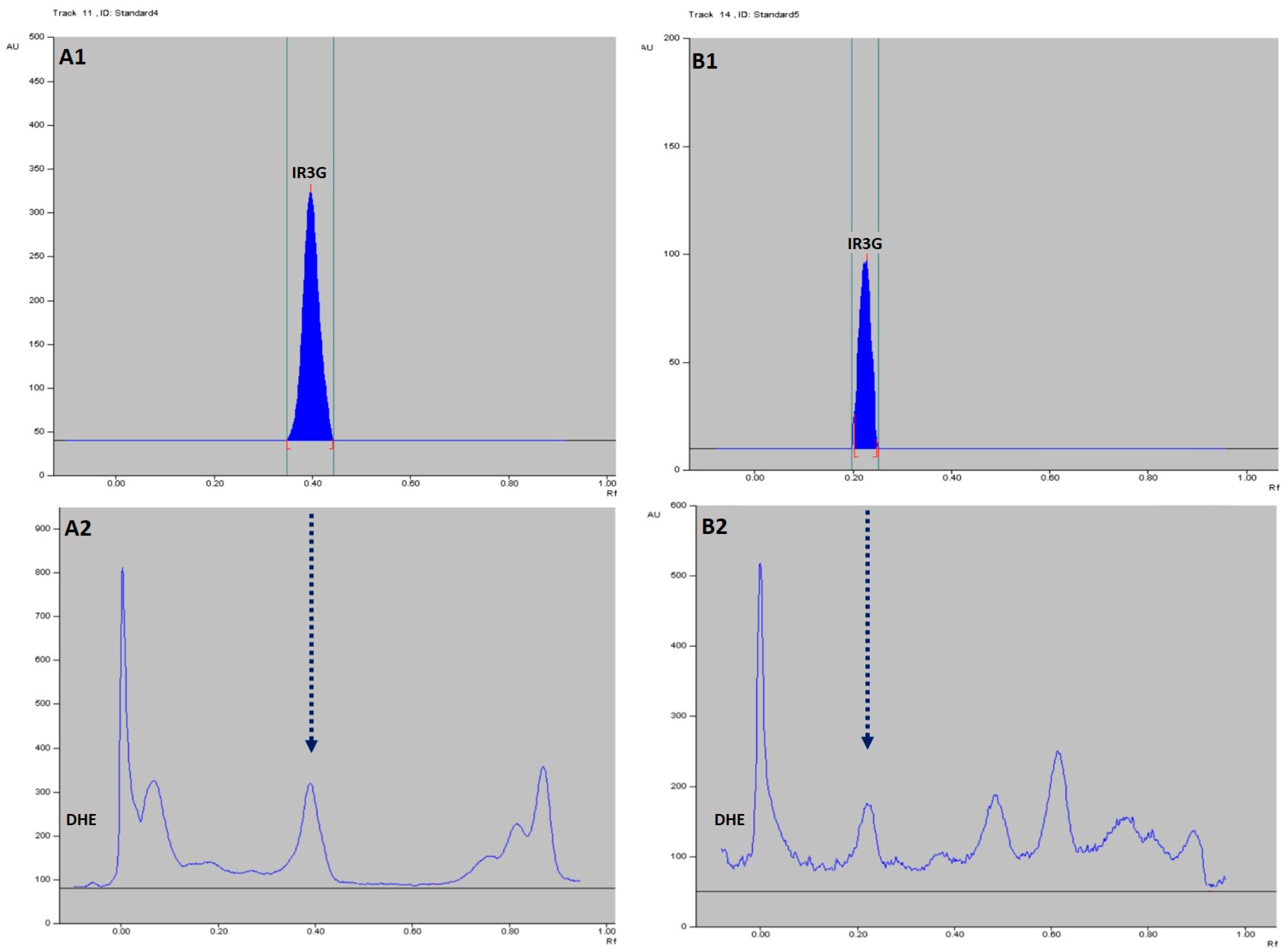
2.4. HPTLC-Based Quantification of IR3G in DHE
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction and Isolation of IR3G
4.3. Determination of the Total Phenolic Content (TPC), Total Flavonoid Content (TFC), and Antioxidant Activity of DHE
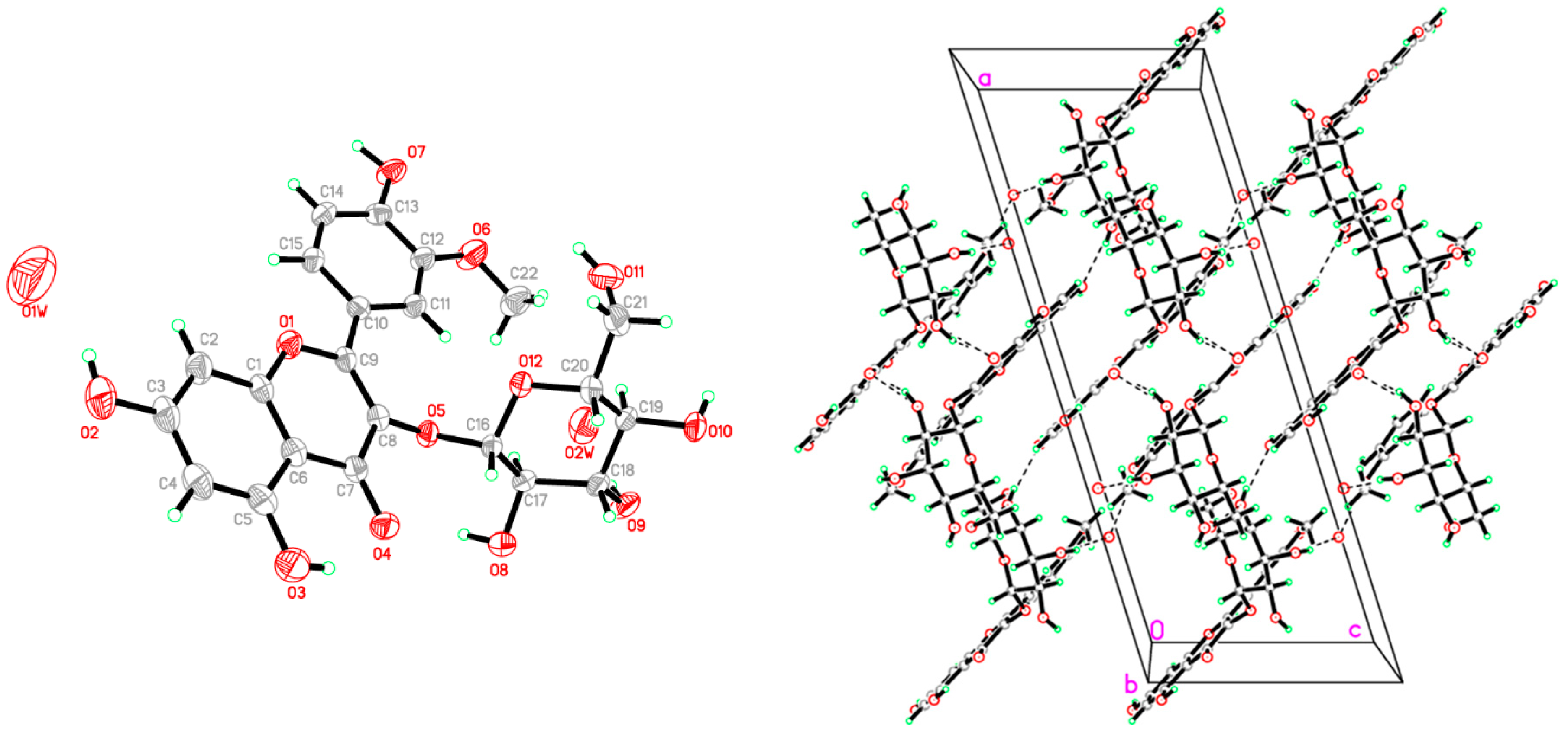
4.4. Crystallographic Study
4.5. Bioassays
4.5.1. In Vitro Anti-Inflammatory Assay
4.5.2. In Vitro Neuroprotective Assay
4.5.3. Locomotor Activity Assay in Zebrafish
4.6. In Silico Study
4.6.1. Molecular Docking Study
4.6.2. Molecular Dynamics (MD) Simulation Study
4.7. Biphasic Qualitative Identification and Quantitative Estimation of IR3G in DHE by HPTLC Densitometry
4.7.1. HPTLC Instrumentation and Conditions
4.7.2. Preparation of Standard Stock Solutions and Calibration Curves
4.7.3. HPTLC Densitometric Quantitative Determination of IR3G in DHE and Method Validation
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Simonian, N.A.; Coyle, J.T. Oxidative stress in neurodegenerative diseases. Annu. Rev. Pharmacol. Toxicol. 1996, 36, 83–106. [Google Scholar] [CrossRef] [PubMed]
- Poulet, B.; Beier, F. Targeting oxidative stress to reduce osteoarthritis. Arthritis Res. Ther. 2016, 18, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, G.; Sharma, A.; Bhatnagar, A. Role of oxidative stress in pathophysiology of rheumatoid arthritis: Insights into NRF2-KEAP1 signalling. Autoimmunity 2021, 54, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.C.; Harre, U.; Purohit, P.; Dietel, K.; Kienhofer, D.; Hahn, J.; Baum, W.; Herrmann, M.; Schett, G.; Mielenz, D. Neurodegeneration enhances the development of arthritis. J. Immunol. 2017, 198, 2394–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, J.; Mathew, D. Nonsteroidal Anti-Inflammatory Drugs Toxicity; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Mallah, K.; Couch, C.; Borucki, D.M.; Toutonji, A.; Alshareef, M.; Tomlinson, S. Anti-inflammatory and neuroprotective agents in clinical trials for CNS disease and injury: Where do we go from here? Front. Immunol. 2020, 11, 2021. [Google Scholar] [CrossRef]
- Rainsford, K.D. Anti-inflammatory drugs in the 21st century. Subcell. Biochem. 2007, 42, 3–27. [Google Scholar]
- Choudhary, M.; Kumar, V.; Malhotra, H.; Singh, S. Medicinal plants with potential anti-arthritic activity. J. Intercult. Ethnopharmacol. 2015, 4, 147–179. [Google Scholar] [CrossRef]
- Physicians’ Desk Reference) Staff. PDR® for Herbal Medicines, 4th ed.; Thomson Healthcare Inc.: Montvale, NJ, USA, 2007. [Google Scholar]
- Nasri, I.; Chawech, R.; Girardi, C.; Mas, E.; Ferrand, A.; Vergnolle, N.; Fabre, N.; Mezghani-Jarraya, R.; Racaud-Sultan, C. Anti-inflammatory and anticancer effects of flavonol glycosides from Diplotaxis harra through GSK3beta regulation in intestinal cells. Pharm. Biol. 2017, 55, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Jdir, H.; Khemakham, B.; Najjaa, H.; Chakroun, M.; Jridi, M.; Ben Arfa, A.; Ben Ali, Y.; Zouari, N. Anti-inflammatory and anti-proliferative activities of the wild edible cruciferous: Diplotaxis simplex. Pharm. Biol. 2016, 54, 2111–2118. [Google Scholar] [CrossRef] [Green Version]
- Oueslati, S.; Ellili, A.; Legault, J.; Pichette, A.; Ksouri, R.; Lachaal, M.; Karray-Bouraoui, N. Phenolic content, antioxidant and anti-inflammatory activities of Tunisian Diplotaxis simplex (Brassicaceae). Nat. Prod. Res. 2015, 29, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Bahloul, N.; Bellili, S.; Aazza, S.; Cherif, A.; Faleiro, M.L.; Antunes, M.D.; Miguel, M.G.; Mnif, W. Aqueous extracts from Tunisian Diplotaxis: Phenol content, antioxidant and anti-Acetylcholinesterase activities, and impact of exposure to simulated gastrointestinal fluids. Antioxidants 2016, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Gleel, W.A.; Hassanien, M.F.R. Antioxidant properties and lipid profile of Diplotaxis harra, Pulicaria incisa and Avicennia marina. Acta Aliment. 2012, 41, 143–151. [Google Scholar] [CrossRef]
- Mititelu, R.R.; Padureanu, R.; Bacanoiu, M.; Padureanu, V.; Docea, A.O.; Calina, D.; Barbulescu, A.L.; Buga, A.M. Inflammatory and oxidative stress markers-mirror tools in rheumatoid arthritis. Biomedicine 2020, 8, 125. [Google Scholar] [CrossRef]
- Ferreira, H.B.; Melo, T.; Paiva, A.; Domingues, M.D.R. Insights in the role of lipids, oxidative stress and inflammation in rheumatoid arthritis unveiled by new trends inlipidomic investigations. Antioxidants 2021, 10, 45. [Google Scholar] [CrossRef]
- Montoya, T.; Sanchez-Hidalgo, M.; Castejon, M.L.; Rosillo, M.A.; Gonzalez-Benjumea, A.; Alarcon-de-la-Lastra, C. Dietary oleocanthal supplementation prevents inflammation and oxidative stress in collagen-induced arthritis in mice. Antioxidants 2021, 10, 650. [Google Scholar] [CrossRef]
- Falleha, H.; Msilini, N.; Oueslati, S.; Ksouri, R.; Magne, C.; Lachaâl, M.; Karray-Bouraoui, N. Diplotaxis harra and Diplotaxis simplex organs: Assessment of phenolics and biological activities before and after fractionation. Ind. Crop. Prod. 2013, 45, 141–147. [Google Scholar] [CrossRef]
- Jdir, H.; Khemakham, B.; Chakroun, M.; Zouari, S.; Ben Ali, Y.; Zouari, N. Diplotaxis simplex suppresses postprandial hyperglycemia in mice by inhibiting key-enzymes linked to type 2 diabetes. Rev. Bras. Farmacog. 2015, 25, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Campo, C. Biology of Brassica coenospecies. In Developments in Plant Genetics and Breeding; Gómez-Campo, C., Ed.; Elsevier Science: Amsterdam, The Netherlands, 1999; Volume 4. [Google Scholar]
- Oberprieler, C.; Zimmer, C.; Bog, M. Are there morphological and life-history traits under climate-dependent differential selection in S Tunesian Diplotaxis harra (Forssk.) Boiss. (Brassicaceae) populations? Ecol. Evol. 2017, 8, 1047–1062. [Google Scholar] [CrossRef] [Green Version]
- Mohammed–Ibtisam, M.I.; Doka, G.M. Check list of flora and vegetation of Hafer Albatin region, northeastern Saudi Arabia. MOJ Ecol. Environ. Sci. 2018, 3, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Atta, E.M.; Hashem, A.I.; Ahmed, A.M.; Elqosy, S.M.; Jaspars, M.; El-Sharkawy, E.R. Phytochemical studies on Diplotaxis harra growing in Sinai. Eur. J. Chem. 2011, 2, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, M.M.D.; El-Sharkawy, E.R.; Matloub, A.A. Cytotoxic flavonoids from Diplotaxis harra (Forssk.) Boiss. growing in Sinai. J. Med. Plants Res. 2011, 5, 5099–5103. [Google Scholar]
- Kassem, M.E.; Afifi, M.S.; Marzouk, M.M.; Mostafa, M.A. Two new flavonol glycosides and biological activities of Diplotaxis harra (Forssk.) Boiss. Nat. Prod. Res. 2013, 27, 2272–2280. [Google Scholar] [CrossRef]
- Hussein, S.R.; Marzouk, M.M.; Kassem, M.E.; Abdel Latif, R.R.; Mohammed, R.S. Chemosystematic significance of flavonoids isolated from Diplotaxis acris (Brassicaceae) and related taxa. Nat. Prod. Res. 2017, 31, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Hashem, F.A.; Saleh, M.M. Antimicrobial components of some Cruciferae plants (Diplotaxis harra Forsk. and Erucaria microcarpa Boiss). Phytother. Res. 1999, 13, 329–332. [Google Scholar] [CrossRef]
- Benzekri, R.; Bouslama, L.; Papetti, A.; Snoussi, M.; Benslimene, I.; Hamami, M.; Limam, F. Isolation and identification of an antibacterial compound from Diplotaxis harra (Forssk.) Boiss. Ind. Crop. Prod. 2016, 80, 228–234. [Google Scholar] [CrossRef]
- Feng, C.W.; Wen, Z.H.; Huang, S.Y.; Hung, H.C.; Chen, C.H.; Yang, S.N.; Chen, N.F.; Wang, H.M.; Hsiao, C.D.; Chen, W.F. Effects of 6-hydroxydopamine exposure on motor activity and biochemical expression in zebrafish (Danio rerio) larvae. Zebrafish 2014, 11, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Fiametti, L.O.; Correa, C.N.; Castro, L.M. Peptide profile of zebrafish brain in a 6-OHDA-induced Parkinson model. Zebrafish 2021, 18, 55–65. [Google Scholar] [CrossRef]
- Elyasi, L.; Jahanshahi, M.; Jameie, S.B.; Hamid Abadi, H.G.; Nikmahzar, E.; Khalili, M.; Jameie, M.; Jameie, M. 6-OHDA mediated neurotoxicity in SH-SY5Y cellular model of Parkinson disease suppressed by pretreatment with hesperidin through activating L-type calcium channels. J. Basic Clin. Physiol. Pharmacol. 2020, 32, 11–17. [Google Scholar] [CrossRef]
- Naglah, A.M.; Ahmed, A.F.; Wen, Z.H.; Al-Omar, M.A.; Amr Ael, G.; Kalmouch, A. New inducible nitric oxide synthase and cyclooxygenase-2 inhibitors, nalidixic acid linked to isatin Schiff bases via certain L-amino acid bridges. Molecules 2016, 21, 498. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.F.; Hsieh, Y.T.; Wen, Z.H.; Wu, Y.C.; Sheu, J.H. Polyoxygenated sterols from the Formosan soft coral Sinularia gibberosa. J. Nat. Prod. 2006, 69, 1275–1279. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Kosaka, T.; Kakimura, J.I.; Matsuoka, Y.; Kohno, Y.; Nomura, Y.; Taniguchi, T. Protective effects of the antiparkinsonian drugs talipexole and pramipexole against 1-methyl-4-phenylpyridinium-induced apoptotic death in human neuroblastoma SH-SY5Y cells. Mol. Pharmacol. 1998, 54, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.F.; Chakraborty, C.; Sung, C.S.; Feng, C.W.; Jean, Y.H.; Lin, Y.Y.; Hung, H.C.; Huang, T.Y.; Huang, S.Y.; Su, T.M.; et al. Neuroprotection by marine-derived compound, 11-dehydrosinulariolide, in an in vitro Parkinson’s model: A promising candidate for the treatment of Parkinson’s disease. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Hilla, S.A.; Liu, X.P.; Borla, M.A.; José, J.V.; O’Malley, D.M. Neurokinematic modeling of complex swimmingpatterns of the larval zebrafish. Neurocomputing 2005, 65–66, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Hollman, P.C.H. Absorption, bioavailability, and metabolism of flavonoids. Pharm. Biol. 2004, 42, 74–83. [Google Scholar] [CrossRef]
- Bokkenheuser, V.D.; Shackleton, C.H.; Winter, J. Hydrolysis of dietary flavonoid glycosides by strains of intestinal Bacteroides from humans. Biochem. J. 1987, 248, 953–956. [Google Scholar] [CrossRef] [Green Version]
- Bennion, B.J.; Essiz, S.G.; Lau, E.Y.; Fattebert, J.L.; Emigh, A.; Lightstone, F.C. A wrench in the works of human acetylcholinesterase: Soman induced conformational changes revealed by molecular dynamics simulations. PLoS ONE 2015, 10, e0121092. [Google Scholar] [CrossRef] [Green Version]
- Junaid, M.; Islam, N.; Hossain, M.K.; Ullah, M.O.; Halim, M.A. Metal based donepezil analogues designed to inhibit human acetylcholinesterase for Alzheimer’s disease. PLoS ONE 2019, 14, e0211935. [Google Scholar] [CrossRef]
- Merzoug, A.; Boucherit, H.; Khaled, R.; Chefiri, A.; Chikhi, A.; Bensegueni, A. Molecular docking study of the acetylcholinesterase inhibition. Curr. Issues Pharm. Med. Sci. 2021, 34, 20–27. [Google Scholar] [CrossRef]
- Kumar, B.; Sheetal, S.; Mantha, A.K.; Kumar, V. Recent developments on the structure–activity relationship studies of MAO inhibitors and their role in different neurological disorders. RSC Adv. 2016, 6, 42660–42683. [Google Scholar] [CrossRef]
- Edmondson, D.E.; Binda, C.; Mattevi, A. Structural insights into the mechanism of amine oxidation by monoamine oxidases A and B. Arch. Biochem. Biophys. 2007, 464, 269–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhujbal, S.P.; Keretsu, S.; Cho, S.J. A combined molecular docking and 3D-QSAR studies on tetrahydropteridin derivatives as PLK2 antagonists. Bull. Korean Chem. Soc. 2019, 40, 796–802. [Google Scholar] [CrossRef]
- Abuelizz, H.A.; Marzouk, M.; Bakhiet, A.; Abdel-Aziz, M.M.; Ezzeldin, E.; Rashid, H.; Al-Salahi, R. In silico study and biological screening of benzoquinazolines as potential antimicrobial agents against methicillin-resistant Staphylococcus aureus, carbapenem-resistant Klebsiella pneumoniae, and fluconazole-resistant Candida albicans. Microb. Pathog. 2021, 160, 105157. [Google Scholar] [CrossRef] [PubMed]
- Brito, A.; Ramirez, J.E.; Areche, C.; Sepulveda, B.; Simirgiotis, M.J. HPLC-UV-MS profiles of phenolic compounds and antioxidant activity of fruits from three citrus species consumed in Northern Chile. Molecules 2014, 19, 17400–17421. [Google Scholar] [CrossRef]
- Dorszewska, J.; Kowalska, M.; Prendecki, M.; Piekut, T.; Kozlowska, J.; Kozubski, W. Oxidative stress factors in Parkinson’s disease. Neural Regen. Res. 2021, 16, 1383–1391. [Google Scholar] [CrossRef]
- Saviano, A.; Casillo, G.M.; Raucci, F.; Pernice, A.; Santarcangelo, C.; Piccolo, M.; Ferraro, M.G.; Ciccone, M.; Sgherbini, A.; Pedretti, N.; et al. Supplementation with ribonucleotide-based ingredient (Ribodiet(R)) lessens oxidative stress, brain inflammation, and amyloid pathology in a murine model of Alzheimer. Biomed. Pharmacother. 2021, 139, 111579. [Google Scholar] [CrossRef]
- Yao, Y.; Chinnici, C.; Tang, H.; Trojanowski, J.Q.; Lee, V.M.; Pratico, D. Brain inflammation and oxidative stress in a transgenic mouse model of Alzheimer-like brain amyloidosis. J. Neuroinflamm. 2004, 1, 21. [Google Scholar] [CrossRef] [Green Version]
- Okayama, Y. Oxidative stress in allergic and inflammatory skin diseases. Curr. Drug Targets Inflamm. Allergy 2005, 4, 517–519. [Google Scholar] [CrossRef]
- Kloska, M.; Mankowska-Wierzbicka, D.; Czlapka-Matyasik, M.; Dobrowolska, A.; Grzymislawski, M. Oxidative stress in etiopathogenesis of inflammatory bowel diseases. Postepy Biochem. 2020, 66, 143–150. [Google Scholar]
- Pickering, R.J. Oxidative stress and inflammation in cardiovascular diseases. Antioxidants 2021, 10, 171. [Google Scholar] [CrossRef]
- Bes, C.; Altunrende, B.; Yilmaz Turkoglu, S.; Yildiz, N.; Soy, M. Parkinsonism in elderly rheumatoid arthritis patients. Clin. Ter. 2014, 165, 19–21. [Google Scholar] [PubMed]
- Jagadeesan, A.J.; Murugesan, R.; Vimala Devi, S.; Meera, M.; Madhumala, G.; Vishwanathan Padmaja, M.; Ramesh, A.; Banerjee, A.; Sushmitha, S.; Khokhlov, A.N.; et al. Current trends in etiology, prognosis and therapeutic aspects of Parkinson’s disease: A review. Acta Biomed. 2017, 88, 249–262. [Google Scholar] [PubMed]
- Hoang, T.; Choi, D.K.; Nagai, M.; Wu, D.C.; Nagata, T.; Prou, D.; Wilson, G.L.; Vila, M.; Jackson-Lewis, V.; Dawson, V.L.; et al. Neuronal NOS and cyclooxygenase-2 contribute to DNA damage in a mouse model of Parkinson disease. Free Radic. Biol. Med. 2009, 47, 1049–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.P.; Vafeiadou, K.; Williams, R.J.; Vauzour, D. Neuroinflammation: Modulation by flavonoids and mechanisms of action. Mol. Asp. Med. 2012, 33, 83–97. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, B.R.; Wang, X.; Kuang, F.; Duan, X.L.; Jiao, X.Y.; Ju, G. ERK1/2 and p38 mitogen-activated protein kinase mediate iNOS-induced spinal neuron degeneration after acute traumatic spinal cord injury. Life Sci. 2006, 79, 1895–1905. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.J.; Lee, J.Y.; Oh, M.S.; Pak, Y.K.; Park, K.S.; Oh, T.H.; Yune, T.Y. Inhibition of inflammation and oxidative stress by Angelica dahuricae radix extract decreases apoptotic cell death and improves functional recovery after spinal cord injury. J. Neurosci. Res. 2012, 90, 243–256. [Google Scholar] [CrossRef]
- Gul, A.; Kunwar, B.; Mazhar, M.; Faizi, S.; Ahmed, D.; Shah, M.R.; Simjee, S.U. Rutin and rutin-conjugated gold nanoparticles ameliorate collagen-induced arthritis in rats through inhibition of NF-kappaB and iNOS activation. Int. Immunopharmacol. 2018, 59, 310–317. [Google Scholar] [CrossRef]
- Chen, L.; Hsieh, M.S.; Ho, H.C.; Liu, Y.H.; Chou, D.T.; Tsai, S.H. Stimulation of inducible nitric oxide synthase by monosodium urate crystals in macrophages and expression of iNOS in gouty arthritis. Nitric Oxide Biol. Chem. 2004, 11, 228–236. [Google Scholar] [CrossRef]
- Sakaguchi, Y.; Shirahase, H.; Ichikawa, A.; Kanda, M.; Nozaki, Y.; Uehara, Y. Effects of selective iNOS inhibition on type II collagen-induced arthritis in mice. Life Sci. 2004, 75, 2257–2267. [Google Scholar] [CrossRef]
- Ma, T.; Kandhare, A.D.; Mukherjee-Kandhare, A.A.; Bodhankar, S.L. Fisetin, a plant flavonoid ameliorates doxorubicin-induced cardiotoxicity in experimental rats: The decisive role of caspase-3, COX-II, cTn-I, iNOs and TNF-alpha. Mol. Biol. Rep. 2019, 46, 105–118. [Google Scholar] [CrossRef]
- Wang, G.J.; Chen, Y.M.; Wang, T.M.; Lee, C.K.; Chen, K.J.; Lee, T.H. Flavonoids with iNOS inhibitory activity from Pogonatherum crinitum. J. Ethnopharmacol. 2008, 118, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-kappaB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-kappaB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007, 2007, 45673. [Google Scholar]
- Gao, X.; Cassidy, A.; Schwarzschild, M.A.; Rimm, E.B.; Ascherio, A. Habitual intake of dietary flavonoids and risk of Parkinson disease. Neurology 2012, 78, 1138–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kukull, W.A. An apple a day to prevent Parkinson disease: Reduction of risk by flavonoids. Neurology 2012, 78, 1112–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shishtar, E.; Rogers, G.T.; Blumberg, J.B.; Au, R.; Jacques, P.F. Long-term dietary flavonoid intake and risk of Alzheimer disease and related dementias in the Framingham Offspring Cohort. Am. J. Clin. Nutr. 2020, 112, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Mandour, D.A.; Bendary, M.A.; Alsemeh, A.E. Histological and imunohistochemical alterations of hippocampus and prefrontal cortex in a rat model of Alzheimer like-disease with a preferential role of the flavonoid “hesperidin”. J. Mol. Histol. 2021, 52, 1043–1065. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Spencer, J.P. Flavonoids, cognition, and dementia: Actions, mechanisms, and potential therapeutic utility for Alzheimer disease. Free Radic. Biol. Med. 2012, 52, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.; Khokhlov, A.; Kopenkin, S.; Bart, B.; Ermolova, T.; Kantemirova, R.; Mazurov, V.; Bell, M.; Caldron, P.; Pillai, L.; et al. Efficacy and safety of flavocoxid compared with naproxen in subjects with osteoarthritis of the knee—A subset analysis. Adv. Ther. 2010, 27, 953–962. [Google Scholar] [CrossRef]
- Elisha, I.L.; Dzoyem, J.P.; McGaw, L.J.; Botha, F.S.; Eloff, J.N. The anti-arthritic, anti-inflammatory, antioxidant activity and relationships with total phenolics and total flavonoids of nine South African plants used traditionally to treat arthritis. BMC Complement. Altern. Med. 2016, 16, 307. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.F.; Zhang, S.L.; He, X.; Yang, X.L.; Wu, H.T.; Lin, B.Q.; Jiang, C.P.; Wang, J.; Yu, C.H.; Yang, Z.L.; et al. Antioxidant effects of Genkwa flos flavonoids on Freunds adjuvant-induced rheumatoid arthritis in rats. J. Ethnopharmacol. 2014, 153, 793–800. [Google Scholar] [CrossRef]
- Magalingam, K.B.; Radhakrishnan, A.; Haleagrahara, N. Protective effects of flavonol isoquercitrin, against 6-hydroxy dopamine (6-OHDA)-induced toxicity in PC12 cells. BMC Res. Notes 2014, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerry, N.; Rice-Evans, C. Inhibition of peroxynitrite-mediated oxidation of dopamine by flavonoid and phenolic antioxidants and their structural relationships. J. Neurochem. 1999, 73, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Cirmi, S.; Maugeri, A.; Lombardo, G.E.; Russo, C.; Musumeci, L.; Gangemi, S.; Calapai, G.; Barreca, D.; Navarra, M. A flavonoid-rich extract of mandarin juice counteracts 6-OHDA-induced oxidative stress in SH-SY5Y cells and modulates Parkinson-related genes. Antioxidants 2021, 10, 539. [Google Scholar] [CrossRef]
- Ma, Z.G.; Wang, J.; Jiang, H.; Liu, T.W.; Xie, J.X. Myricetin reduces 6-hydroxydopamine-induced dopamine neuron degeneration in rats. Neuroreport 2007, 18, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Ghaffari, F.; Moghaddam, A.H.; Zare, M. Neuroprotective effect of quercetin nanocrystal in a 6-hydroxydopamine model of Parkinson disease: Biochemical and behavioral evidence. Basic Clin. Neurosci. 2018, 9, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Raza, S.S.; Javed, H.; Ahmad, A.; Khan, A.; Islam, F.; Safhi, M.M.; Islam, F. Rutin protects dopaminergic neurons from oxidative stress in an animal model of Parkinson’s disease. Neurotox. Res. 2012, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Pu, X.P. Neuroprotective effect of kaempferol against a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced mouse model of Parkinson’s disease. Biol. Pharm. Bull. 2011, 34, 1291–1296. [Google Scholar] [CrossRef] [Green Version]
- Lv, C.; Hong, T.; Yang, Z.; Zhang, Y.; Wang, L.; Dong, M.; Zhao, J.; Mu, J.; Meng, Y. Effect of quercetin in the 1-methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-induced mouse model of Parkinson’s disease. Evid. Based Complement. Alternat. Med. 2012, 2012, 928643. [Google Scholar] [CrossRef]
- Villaño, D.; Fernández-Pachón, M.S.; Troncoso, A.M.; García-Parrilla, M.C. Comparison of antioxidant activity of wine phenolic compounds and metabolites in vitro. Anal. Chim. Acta 2005, 538, 391–398. [Google Scholar] [CrossRef]
- Leopoldini, M.; Russo, N.; Toscano, M. The molecular basis of working mechanism of natural polyphenolic antioxidants. Food Chem. 2011, 125, 288–306. [Google Scholar] [CrossRef]
- Kejik, Z.; Kaplanek, R.; Masarik, M.; Babula, P.; Matkowski, A.; Filipensky, P.; Vesela, K.; Gburek, J.; Sykora, D.; Martasek, P.; et al. Iron complexes of flavonoids-antioxidant capacity and beyond. Int. J. Mol. Sci. 2021, 22, 646. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.L.; Zhang, C.W. Molecular events underlying Parkinson’s disease—An interwoven tapestry. Front Neurol 2013, 4, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Boer, V.C.; Dihal, A.A.; van der Woude, H.; Arts, I.C.; Wolffram, S.; Alink, G.M.; Rietjens, I.M.; Keijer, J.; Hollman, P.C. Tissue distribution of quercetin in rats and pigs. J. Nutr. 2005, 135, 1718–1725. [Google Scholar] [CrossRef] [Green Version]
- Youdim, K.A.; Qaiser, M.Z.; Begley, D.J.; Rice-Evans, C.A.; Abbott, N.J. Flavonoid permeability across an in situ model of the blood-brain barrier. Free Radic. Biol. Med. 2004, 36, 592–604. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, P.; Malik, N.; Sobarzo-Sanchez, E.; Uriarte, E.; Khatkar, A. Quercetin and related chromenone derivatives as monoamine oxidase Inhibitors: Targeting neurological and mental Disorders. Molecules 2019, 24, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inglis, K.J.; Chereau, D.; Brigham, E.F.; Chiou, S.S.; Schöbel, S.; Frigon, N.L.; Yu, M.; Caccavello, R.J.; Nelson, S.; Motter, R.; et al. Polo-like kinase 2 (PLK2) phosphorylates alpha-synuclein at serine 129 in central nervous system. J. Biol. Chem. 2009, 284, 2598–2602. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Munishkina, L.A.; Fink, A.L.; Uversky, V.N. Effects of various flavonoids on the α-synuclein fibrillation process. Parkinsons Dis. 2010, 2010, 650794. [Google Scholar] [CrossRef] [Green Version]
- Saleem, U.; Chauhdary, Z.; Raza, Z.; Shah, S.; Rahman, M.U.; Zaib, P.; Ahmad, B. Anti-Parkinson’s activity of Tribulus terrestris via modulation of AChE, α-synuclein, TNF-α, and IL-1β. ACS Omega 2020, 5, 25216–25227. [Google Scholar] [CrossRef]
- Pagano, G.; Rengo, G.; Pasqualetti, G.; Femminella, G.D.; Monzani, F.; Ferrara, N.; Tagliati, M. Cholinesterase inhibitors for Parkinson’s disease: A systematic review and meta-analysis. J. Neurol. Neurosurg. Psychiatry 2015, 86, 767–773. [Google Scholar] [CrossRef]
- Ahmed, A.F.; Al-Qahtani, J.H.; Al-Yousef, H.M.; Al-Said, M.S.; Ashour, A.E.; Al-Sohaibani, M.; Rafatullah, S. Proanthocyanidin-rich date seed extract protects against chemically induced hepatorenal toxicity. J. Med. Food 2015, 18, 280–289. [Google Scholar] [CrossRef]
- Nenadis, N.; Wang, L.F.; Tsimidou, M.; Zhang, H.Y. Estimation of Scavenging Activity of Phenolic Compounds Using the ABTS•+ Assay. J. Agric. Food Chem. 2004, 52, 4669–4674. [Google Scholar] [CrossRef] [PubMed]
- Behery, F.A.; Naeem, Z.E.; Maatooq, G.T.; Amer, M.M.; Wen, Z.H.; Sheu, J.H.; Ahmed, A.F. Phenanthrenoids from Juncus acutus L. New natural lipopolysaccharide-inducible nitric oxide synthase inhibitors. Chem. Pharm. Bull. 2007, 55, 1264–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Feng, C.W.; Hung, H.C.; Huang, S.Y.; Chen, C.H.; Chen, Y.R.; Chen, C.Y.; Yang, S.N.; Wang, H.D.; Sung, P.J.; Sheu, J.H.; et al. Neuroprotective effect of the marine-derived compound 11-dehydrosinulariolide through DJ-1-related pathway in in vitro and in vivo models of Parkinson’s disease. Mar. Drugs 2016, 14, 187. [Google Scholar] [CrossRef]
- Brooks, B.R.; Brooks, C.L., III; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kale, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Al-Yousef, H.M.; Alhowiriny, T.A.; Alam, P.; Siddiqui, N.A.; Ahmed, A.F.; Al-Qahtani, J.H.; Amina, M. Simultaneous quantification of two phenolic biomarkers by a validated high-performance thin-layer chromatographic method in antimicrobial and antioxidant active ethyl acetate fraction of Allium cepa L. (Peel). JPC J. Planar Chromat. 2017, 30, 510–515. [Google Scholar] [CrossRef]
- Sheldrick, G.M. A short history of SHELX. Acta Crystallogr. A 2008, 64 Pt 1, 112–122. [Google Scholar]
- Sheldrick, G.M. SHELXTL‐PC (Version 5.1); Siemens Analytical. Instruments, Inc.: Madison, WI, USA, 1997. [Google Scholar]
- Allen, F.H.; Kennard, O.; Watson, D.G.; Brammer, L.; Orpen, A.G.; Taylor, R. Tables of Bond Lengths determined by X‐Ray and Neutron Diffraction. Part. 1. Bond Lengths in Organic Compounds. J. Chem. Soc. Chem. Soc. Perkins Trans. 2 1987, 12, S1–S19. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Value | |
|---|---|---|
| Phenol content a | (mg GAE/g) | 80.43 ± 0.01 |
| Flavonoid content b | (mg QUE/g) | 54.26 ± 0.03 |
| Antioxidant activity | (IC50 μg/mL) c | 247.4 [5.9] e |
| (IC50 μg/mL) d | 203.7 [6.8] f | |
| Compound Code | Target (PDB Code) | Ligand | Receptor | Interaction | Distance (Å) | E (kcal/mol) | Docking Score (kcal/mol) |
|---|---|---|---|---|---|---|---|
| IR3G | AChE (4M0E) | O 3 | OD2 ASP74 (A) | H-donor | 2.10 | −1.9 | −7.7045 |
| O 4 | OH TYR124 (A) | H-acceptor | 2.18 | −0.1 | |||
| O 6 | N PHE295 (A) | H-acceptor | 2.29 | −0.5 | |||
| C 17 | 6-ring TRP286 (A) | H-pi | 2.61 | −0.6 | |||
| MAO-B (6FVZ) | O 3 | O PRO102 (A) | H-donor | 2.72 | −1.8 | −5.863 | |
| O 6 | O LEU164 (A) | H-donor | 3.52 | −0.2 | |||
| C 29 | SG CYS172 (A) | H-donor | 3.06 | −0.3 | |||
| O 8 | CD1 PHE168 (A) | H-acceptor | 3.38 | 0.6 | |||
| 6-ring | CA ILE199 (A) | pi-H | 4.08 | −0.6 | |||
| 6-ring | CA ILE199 (A) | pi-H | 4.94 | −0.2 | |||
| PLK-2 (4I5P) | O 3 | OD1 ASP223 (A) | H-donor | 3.23 | −1.3 | −7.7340 | |
| O 4 | OD1 ASN210 (A) | H-donor | 3.14 | −1.2 | |||
| O 5 | SG CYS96 (A) | H-donor | 4.29 | −1.0 | |||
| O 6 | O GLY209 (A) | H-donor | 3.07 | −1.1 | |||
| 6-ring | CB LEU88 (A) | pi-H | 4.11 | −0.8 | |||
| 6-ring | CB LEU88 (A) | pi-H | 4.61 | −0.4 | |||
| 6-ring | CD1 LEU88 (A) | pi-H | 4.51 | −0.5 | |||
| 6-ring | CD2 LEU88 (A) | pi-H | 4.5 | −0.5 | |||
| 6-ring | CG ARG165 (A) | pi-H | 4.99 | −0.3 | |||
| IR | AChE (4M0E) | O 5 | N ARG296 (A) | H-acceptor | 3.31 | −2.2 | −6.8919 |
| C 23 | 5-ring TRP286 (A) | H-pi | 4.01 | −0.5 | |||
| 6-ring | CE1 PHE297 (A) | pi-H | 4.86 | −0.3 | |||
| 6-ring | 6-ring TRP286 (A) | pi-pi | 3.79 | −0.1 | |||
| MAO-B (6FVZ) | O 6 | O PRO102 (A) | H-donor | 1.85 | −3.2 | −7.5290 | |
| O 5 | CA PHE168 (A) | H-acceptor | 2.11 | −1.1 | |||
| 6-ring | CA CYS172 (A) | pi-H | 3.72 | −0.4 | |||
| 6-ring | CB ILE199 (A) | pi-H | 3.78 | −0.4 | |||
| PLK-2 (4I5P) | O 6 | O LEU88 (A) | H-donor | 3.09 | −1.5 | −6.4320 | |
| C 21 | SG CYS96 (A) | H-donor | 4.04 | −0.5 | |||
| O 2 | N CYS162 (A) | H-acceptor | 3.11 | −0.4 | |||
| O 5 | N CYS162 (A) | H-acceptor | 2.97 | −1.3 | |||
| 6-ring | CB LEU88 (A) | pi-H | 4.69 | −0.3 | |||
| 6-ring | CB LEU88 (A) | pi-H | 4.08 | −0.4 | |||
| 6-ring | CD1 LEU88 (A) | pi-H | 4.08 | −0.8 | |||
| 6-ring | CD1 LEU88 (A) | pi-H | 4.47 | −0.5 | |||
| 6-ring | CD2 LEU88 (A) | pi-H | 4.04 | −0.7 | |||
| 6-ring | CG ARG165 (A) | pi-H | 4.75 | −0.4 |
| NP-HPTLC | RP-HPTLC | |
|---|---|---|
| Linearity range (ng/band) | 100–400 | 100–500 |
| Regression equation | Y = 7.933 + 0.4428X | Y= 0.816 + 0.346X |
| Correlation coefficient (r2) | 0.99296 | 0.99018 |
| Slope | 0.4428 | 0.346 |
| Standard deviation (Sdv) | 5.43% | 7.09% |
| Limit of detection (LOD; ng) | 40.47 | 67.6 |
| Limit of quantification (LOQ; ng) | 122.63 | 204.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, A.F.; Wen, Z.-H.; Bakheit, A.H.; Basudan, O.A.; Ghabbour, H.A.; Al-Ahmari, A.; Feng, C.-W. A Major Diplotaxis harra-Derived Bioflavonoid Glycoside as a Protective Agent against Chemically Induced Neurotoxicity and Parkinson’s Models; In Silico Target Prediction; and Biphasic HPTLC-Based Quantification. Plants 2022, 11, 648. https://doi.org/10.3390/plants11050648
Ahmed AF, Wen Z-H, Bakheit AH, Basudan OA, Ghabbour HA, Al-Ahmari A, Feng C-W. A Major Diplotaxis harra-Derived Bioflavonoid Glycoside as a Protective Agent against Chemically Induced Neurotoxicity and Parkinson’s Models; In Silico Target Prediction; and Biphasic HPTLC-Based Quantification. Plants. 2022; 11(5):648. https://doi.org/10.3390/plants11050648
Chicago/Turabian StyleAhmed, Atallah F., Zhi-Hong Wen, Ahmed H. Bakheit, Omer A. Basudan, Hazem A. Ghabbour, Abdullah Al-Ahmari, and Chien-Wei Feng. 2022. "A Major Diplotaxis harra-Derived Bioflavonoid Glycoside as a Protective Agent against Chemically Induced Neurotoxicity and Parkinson’s Models; In Silico Target Prediction; and Biphasic HPTLC-Based Quantification" Plants 11, no. 5: 648. https://doi.org/10.3390/plants11050648