
Genome Estimation and Phytochemical Compound Identification in the Leaves and Callus of Abrus precatorius: A Locally Endangered Plant from the Flora of Saudi Arabia

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Molecular Phylogenetic Identification of Abrus precatorius
2.2. Callus Induction
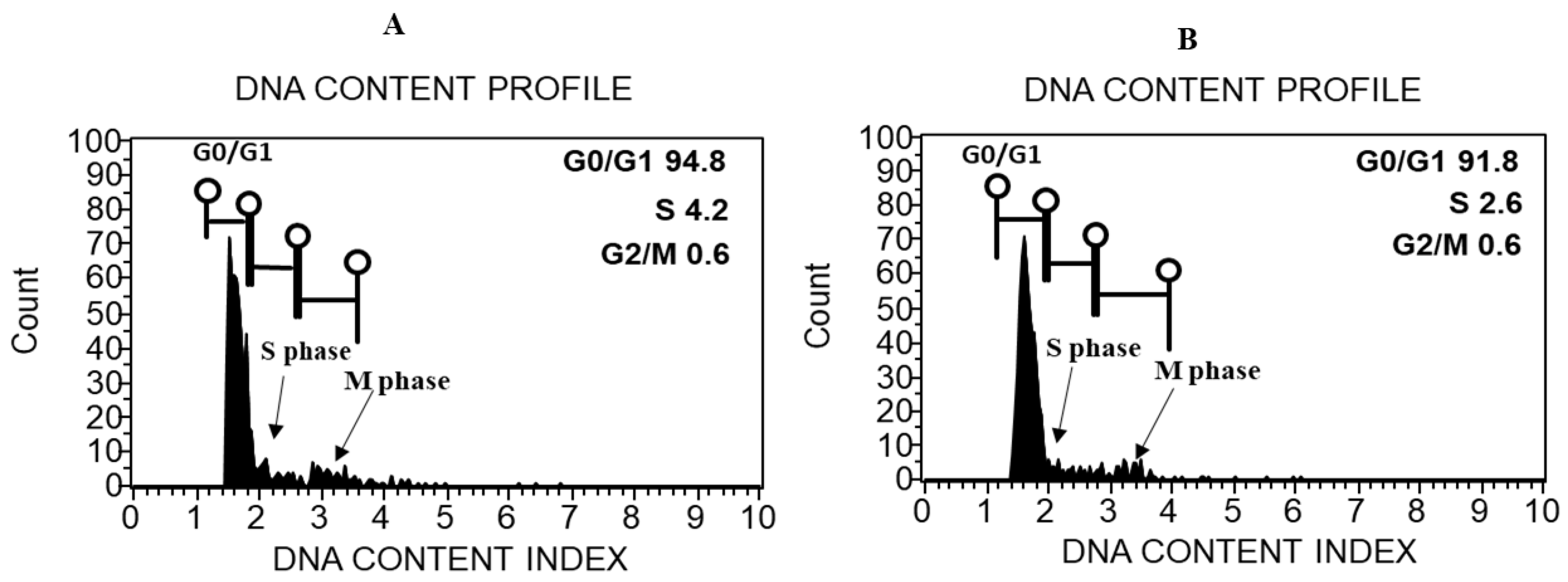
2.3. DNA Content Estimation through Flow Cytometric Analysis
2.4. Phytochemical Analysis
2.4.1. Phenol and Flavonoid Contents
2.4.2. GC-MS Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material Collection
4.2. Callus Induction
4.3. DNA Extraction and ITS Identification
4.4. Genome Size Estimation
4.4.1. Nuclei Extraction
4.4.2. Flow Cytometric Analysis
4.5. Total Phenolic and Total Flavonoid Content
4.5.1. Extract Preparation
4.5.2. Total Phenolic Content Estimation
4.5.3. Total Flavonoid Content Estimation
4.6. HPLC Quantification of Gallic Acid, Quercetin, and Rutin
4.7. Gas Chromatography Mass Spectrometry Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rahman, M.A.; Mossa, J.S.; Al-Said, M.S.; Al-Yahya, M.A. Medicinal plant diversity in the flora of Saudi Arabia 1: A report on seven plant families. Fitoterapia 2004, 75, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Al-Khulaidi, A.; Al-Sagheer, N.A.; Al-Turki, T.; Filimban, F. Inventory of most rare and endangered plant species in Albaha region Saudi Arabia. Ijbpas 2018, 7, 443–460. [Google Scholar]
- Thomas, J.; El-Sheikh, M.A.; Alatar, A.A. Endemics and endangered species in the biodiversity hotspot of the Shada Mountains, Saudi Arabia. J. Arid. Land 2017, 9, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Al-Asmari, A.K.; Abbasmanthiri, R.; Osman, N.M.A.; Al-Asmari, B.A. Endangered Saudi Arabian plants having ethnobotanical evidence as antidotes for scorpion envenoming. Clin. Phytosci. 2020, 6, 1–13. [Google Scholar] [CrossRef]
- Abbas, A.M.; Al-Kahtani, M.A.; Alfaifi, M.Y.; Elbehairi, S.E.I.; Badry, M.O. Floristic diversity and phytogeography of Jabal Fayfa: A subtropical dry zone, south-west Saudi Arabia. Diversity 2020, 12, 345. [Google Scholar] [CrossRef]
- Garaniya, N.; Bapodra, A. Ethno botanical and Phytophrmacological potential of Abrus precatorius L.: A review. Asian Pac. J. Trop. Biomed. 2014, 4, S27–S34. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, M.; Siddiqui, N.; Gupta, S. Abrus precatorius (L.): An evaluation of traditional herb. J. Pharm. Res. 2013, 3, 3296–3315. [Google Scholar]
- Parthasarathy, V.; TV, A.K. Screening of potential GCMS derived antimigraine compound from the leaves of Abrus precatorius Linn to target “calcitonin gene related peptide” receptor using in silico analysis. Food Sci. Hum. Wellness 2019, 8, 34–39. [Google Scholar]
- Wan-Ibrahim, W.S.; Tuan Ismail, T.N.N.; Mohd-Salleh, S.F.; Ismail, N. GC-MS analysis of phytochemical compounds in aqueous leaf extract of Abrus precatorius. Pertanika J. Trop. Agric. Sci. 2018, 41, 241–250. [Google Scholar]
- Pavithra, K.; Uddandrao, V.S.; Mathavan, S.; Gobeeswaran, N.; Vadivukkarasi, S.; Ganapathy, S. Identification of bioactive factors from Abrus precatorius by GC–MS, NMR and evaluation of its antioxidant activity. Mater. Today: Proc. 2020, 26, 3518–3521. [Google Scholar] [CrossRef]
- Biswas, A.; Roy, M.; Miah, M.B.; Bhadra, S. In vitro propagation of Abrus precatorius L.-a rare medicinal plant of Chittagong hill tracts. Plant Tissue Cult. Biotechnol. 2007, 17, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Zafar, N.; Mujib, A.; Ali, M.; Tonk, D.; Gulzar, B.; Malik, M.; Sayeed, R.; Mamgain, J. Genome size analysis of field grown and tissue culture regenerated Rauvolfia serpentina (L.) by flow cytometry: Histology and scanning electron microscopic study for in vitro morphogenesis. Ind. Crops Prod. 2019, 128, 545–555. [Google Scholar] [CrossRef]
- Thiem, B.; Śliwińska, E. Flow cytometric analysis of nuclear DNA content in cloudberry (Rubus chamaemorus L.) in vitro cultures. Plant Sci. 2003, 164, 129–134. [Google Scholar] [CrossRef]
- Orzechowska, M.; Stępień, K.; Kamińska, T.; Siwińska, D. Chromosome variations in regenerants of Arabidopsis thaliana derived from 2-and 6-week-old callus detected using flow cytometry and FISH analyses. Plant Cell Tissue Organ Cult. (PCTOC) 2013, 112, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Galbraith, D.W. Analysis of higher plants by flow cytometry and cell sorting. Int. Rev. Cytol. 1989, 116, 165–228. [Google Scholar]
- Bourge, M.; Brown, S.C.; Siljak-Yakovlev, S. Flow cytometry as tool in plant sciences, with emphasis on genome size and ploidy level assessment. Genet. Appl. 2018, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Pellicer, J.; Powell, R.F.; Leitch, I.J. The application of flow cytometry for estimating genome size, ploidy level endopolyploidy, and reproductive modes in plants. In Molecular Plant Taxonomy; Springer: Cham, Switzerland, 2021; pp. 325–361. [Google Scholar]
- Liu, K.; Chen, Q.; Liu, Y.; Zhou, X.; Wang, X. Isolation and biological activities of decanal, linalool, valencene, and octanal from sweet orange oil. J. Food Sci. 2012, 77, C1156–C1161. [Google Scholar] [CrossRef]
- Nisha, S.R.; Jeeva, S.; Paul Raj, K. Chemical profiling and compound Isolation of different extracts of Boucerosia pauciflora Wight using GCMS Analysis. J. Emerg. Technol. Innov. Res. 2018, 5, 6. [Google Scholar]
- Caboni, P.; Ntalli, N.G.; Aissani, N.; Cavoski, I.; Angioni, A. Nematicidal activity of (E, E)-2, 4-decadienal and (E)-2-decenal from Ailanthus altissima against Meloidogyne javanica. J. Agric. Food Chem. 2012, 60, 1146–1151. [Google Scholar] [CrossRef]
- Kawuri, R.; Darmayasa, I. Bioactive compound from extract filtrat streptomyces sp. Sp1. as biocontrol of vibriosis on larvae of Macrobrachium rosenbergii shrimps. Hayati J. Biosci. 2019, 26, 15. [Google Scholar] [CrossRef]
- Sahin, N.; Kula, I.; Erdogan, Y. Investigation of antimicrobial activities of nonanoic acid derivatives. Fresenius Environ. Bull. 2006, 15, 141–143. [Google Scholar]
- PATIL, V.-I.R.; Donde, K.J.; Jadhav, S.B.; Malve, S.P. Synthesis and antimicrobial activity of 3-hydroxyimino-5-methyl-2-hexanone (himh) and its dioxime derivative. Acta Pol. Pharm. 2002, 59, 223–225. [Google Scholar] [PubMed]
- Yildiz, H. Chemical composition, antimicrobial, and antioxidant activities of essential oil and ethanol extract of Coriandrum sativum L. leaves from Turkey. Int. J. Food Prop. 2016, 19, 1593–1603. [Google Scholar] [CrossRef] [Green Version]
- Jenecius, A.; Uthayakumaria, F.; Mohan, V. GC-MS determination of bioactive components of Sauropus bacciformis blume (Euphorbiaceae). J. Curr. Chem. Pharm. Sci. 2012, 2, 347–358. [Google Scholar]
- Kumar Tyagi, A.; Bukvicki, D.; Gottardi, D.; Veljic, M.; Guerzoni, M.E.; Malik, A.; Marin, P.D. Antimicrobial potential and chemical characterization of serbian liverwort (Porella arboris-vitae): SEM and TEM observations. Evid.-Based Complement. Altern. Med. 2013, 2013, 382927. [Google Scholar] [CrossRef] [Green Version]
- Chanprapai, P.; Kubo, I.; Chavasiri, W. Anti-rice pathogenic microbial activity of Persicaria sp. Extracts. Sci. Technol. Asia 2018, 23, 32–41. [Google Scholar]
- Nahab, F.B.; Handforth, A.; Brown, T.; Shin, C.; Quesada, A.; Dong, C.; Haubenberger, D.; Hallett, M. Octanoic acid suppresses harmaline-induced tremor in mouse model of essential tremor. Neurotherapeutics 2012, 9, 635–638. [Google Scholar] [CrossRef] [Green Version]
- Yücel, T.B.; Karaoğlu, Ş.A.; Yaylı, N. Antimicrobial Activity and Composition of Rindera lanata (LAM.) Bunge var. canescens (ADC) Kosn. Essential oil Obtained by Hydrodistillation and Microwave Assisted Distillation. Rec. Nat. Prod. 2017, 11, 328–333. [Google Scholar]
- Bittencourt, M.L.; Ribeiro, P.R.; Franco, R.L.; Hilhorst, H.W.; de Castro, R.D.; Fernandez, L.G. Metabolite profiling, antioxidant and antibacterial activities of Brazilian propolis: Use of correlation and multivariate analyses to identify potential bioactive compounds. Food Res. Int. 2015, 76, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Ricciardelli, A.; Casillo, A.; Corsaro, M.M.; Tutino, M.L.; Parrilli, E.; van der Mei, H.C. Pentadecanal and pentadecanoic acid coatings reduce biofilm formation of Staphylococcus epidermidis on PDMS. Pathog. Dis. 2020, 78, ftaa012. [Google Scholar] [CrossRef]
- Vanitha, V.; Vijayakumar, S.; Nilavukkarasi, M.; Punitha, V.; Vidhya, E.; Praseetha, P. Heneicosane—A novel microbicidal bioactive alkane identified from Plumbago zeylanica L. Ind. Crops Prod. 2020, 154, 112748. [Google Scholar] [CrossRef]
- Shaaban, M.T.; Ghaly, M.F.; Fahmi, S.M. Antibacterial activities of hexadecanoic acid methyl ester and green-synthesized silver nanoparticles against multidrug-resistant bacteria. J. Basic Microbiol. 2021, 61, 557–568. [Google Scholar] [CrossRef] [PubMed]
- Aparna, V.; Dileep, K.V.; Mandal, P.K.; Karthe, P.; Sadasivan, C.; Haridas, M. Anti-inflammatory property of n-hexadecanoic acid: Structural evidence and kinetic assessment. Chem. Biol. Drug Des. 2012, 80, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Jenifer, D.R.; Malathy, B.; SS, A. In vitro and in silico studies on the biochemistry and anti-cancer activity of phytochemicals from Plumb. Zeylanica. Indian J. Biochem. Biophys. (IJBB) 2021, 58, 272–283. [Google Scholar]
- Hanafy, S.M.; Abd El-Shafea, Y.M.; Saleh, W.D.; Fathy, H.M. Chemical profiling, in vitro antimicrobial and antioxidant activities of pomegranate, orange and banana peel-extracts against pathogenic microorganisms. J. Genet. Eng. Biotechnol. 2021, 19, 1–10. [Google Scholar] [CrossRef]
- Ali, A.; Javaid, A.; Shoaib, A. GC-MS analysis and antifungal activity of methanolic root extract of Chenopodium album against Sclerotium rolfsii. Planta Daninha 2017, 35, 35. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Sin, Y.M.; Lee, S.O.; Lee, T.H. Enzymatic synthesis of an acylated phloroglucinol derivative with phloroglucinol and vinyl octanoate in acetonitrile. Biotechnol. Lett. 2002, 24, 181–184. [Google Scholar] [CrossRef]
- Yue, X.-F.; Shang, X.; Zhang, Z.-J.; Zhang, Y.-N. Phytochemical composition and antibacterial activity of the essential oils from different parts of sea buckthorn (Hippophae rhamnoides L.). J. Food Drug Anal. 2017, 25, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Kelechi Oleru, A.O.; Olayiwola, J.; Popoola, B. Potential Antimicrobial Substances from the Characterized Bioactive Compounds Extracted from Secondary Metabolites of Aspergillus terreus. Res. J. Microbiol. 2021, 16, 10. [Google Scholar]
- Uma, B.; Prabhakar, K.; Rajendran, S.; Lakshmi, S.Y. Studies on GC/MS spectroscopic analysis of some bioactive antimicrobial compounds from Cinnamomum zeylanicum. J. Med. Plants 2009, 8, 125–131. [Google Scholar]
- Begum, I.F.; Mohankumar, R.; Jeevan, M.; Ramani, K. GC–MS analysis of bio-active molecules derived from Paracoccus pantotrophus FMR19 and the antimicrobial activity against bacterial pathogens and MDROs. Indian J. Microbiol. 2016, 56, 426–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrufo, T.; Nazzaro, F.; Mancini, E.; Fratianni, F.; Coppola, R.; De Martino, L.; Agostinho, A.B.; De Feo, V. Chemical composition and biological activity of the essential oil from leaves of Moringa oleifera Lam. cultivated in Mozambique. Molecules 2013, 18, 10989–11000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhetso, T.; Shubharani, R.; Roopa, M.; Sivaram, V. Chemical constituents, antioxidant, and antimicrobial activity of Allium chinense G. Don. Future J. Pharm. Sci. 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Bharath, M.; Azeem, M.; Basha, S.; Keerthan, H. Antimicrobial Activity of Cinnamon Extracts against Foodborne Pathogens E. coli, S. tyhimurium and S. aureus and L. monocytogens. Curr. Nutr. Food Sci. 2016, 11, 66–72. [Google Scholar]
- Abubakar, M.N.; Majinda, R.R. GC-MS analysis and preliminary antimicrobial activity of Albizia adianthifolia (Schumach) and Pterocarpus angolensis (DC). Medicines 2016, 3, 3. [Google Scholar] [CrossRef] [Green Version]
- Lotfy, M.M.; Hassan, H.M.; Hetta, M.H.; El-Gendy, A.O.; Mohammed, R. Di-(2-ethylhexyl) Phthalate, a major bioactive metabolite with antimicrobial and cytotoxic activity isolated from River Nile derived fungus Aspergillus awamori. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 263–269. [Google Scholar] [CrossRef]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal identification using molecular tools: A primer for the natural products research community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Natl. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef] [Green Version]
- Pathak, M.R.; Mohamed, A.A.; Farooq, M. DNA barcoding and identification of medicinal plants in the kingdom of Bahrain. Am. J. Plant Sci. 2018, 9, 2757–2774. [Google Scholar] [CrossRef]
- Zhao, L.L.; Feng, S.J.; Tian, J.Y.; Wei, A.Z.; Yang, T.X. Internal transcribed spacer 2 (ITS 2) barcodes: A useful tool for identifying Chinese Zanthoxylum. Appl. Plant Sci. 2018, 6, e01157. [Google Scholar] [CrossRef]
- Prem, D.; Singh, S.; Gupta, P.P.; Singh, J.; Kadyan, S.P.S. Callus induction and de novo regeneration from callus in Guar (Cyamopsis tetragonoloba). Plant Cell Tissue Organ Cult. 2005, 80, 209–214. [Google Scholar] [CrossRef]
- Deng, X.; Xiong, Y.; Li, J.; Yang, D.; Liu, J.; Sun, H.; Song, H.; Wang, Y.; Ma, J.; Liu, Y. The Establishment of an Efficient Callus Induction System for Lotus (Nelumbo nucifera). Plants 2020, 9, 1436. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.I.; Rashid, H.; Yasmin, T.; Minhas, N.M. Effect of 2, 4-dichlorophenoxyacetic acid on callus induction from mature wheat (Triticum aestivum L.) seeds. Int. J. Agric. Biol. 2003, 6, 156–159. [Google Scholar]
- Sliwinska, E.; Thiem, B. Genome size stability in six medicinal plant species propagated in vitro. Biol. Plant. 2007, 51, 556–558. [Google Scholar] [CrossRef]
- Sakhanokho, H.F.; Babiker, E.M.; Smith, B.J.; Drackett, P.R. High-frequency somatic embryogenesis, nuclear DNA estimation of milkweed species (Asclepias latifolia, A. speciosa, and A. subverticillata), and genome size stability of regenerants. Plant Cell Tissue Organ Cult. (PCTOC) 2019, 137, 149–156. [Google Scholar] [CrossRef]
- Slazak, B.; Sliwinska, E.; Saługa, M.; Ronikier, M.; Bujak, J.; Słomka, A.; Göransson, U.; Kuta, E. Micropropagation of Viola uliginosa (Violaceae) for endangered species conservation and for somaclonal variation-enhanced cyclotide biosynthesis. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 120, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Lucia, G.; Castiglione, M.R.; Turrini, A.; Ronchi, V.N.; Geri, C. Cytogenetic and histological approach for early detection of “mantled” somaclonal variants of oil palm regenerated by somatic embryogenesis: First results on the characterization of regeneration system. Caryologia 2011, 64, 223–234. [Google Scholar] [CrossRef]
- Makowczyńska, J.; Andrzejewska-Golec, E.; Sliwinska, E. Nuclear DNA content in different plant materials of Plantago asiatica L. cultured in vitro. Plant Cell Tissue Organ Cult. 2008, 94, 65–71. [Google Scholar] [CrossRef]
- Borchert, T.; Fuchs, J.; Winkelmann, T.; Hohe, A. Variable DNA content of Cyclamen persicum regenerated via somatic embryogenesis: Rethinking the concept of long-term callus and suspension cultures. Plant Cell Tissue Organ Cult. 2007, 90, 255–263. [Google Scholar] [CrossRef]
- Park, J.-S.; Seong, Z.-K.; Kim, M.-S.; Ha, J.-H.; Moon, K.-B.; Lee, H.-J.; Lee, H.-K.; Jeon, J.-H.; Park, S.U.; Kim, H.-S. Production of flavonoids in callus cultures of Sophora flavescens Aiton. Plants 2020, 9, 688. [Google Scholar] [CrossRef]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and other phenolic compounds from medicinal plants for pharmaceutical and medical aspects: An overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef] [PubMed]
- Rameshkumar, R.; Satish, L.; Pandian, S.; Rathinapriya, P.; Rency, A.S.; Shanmugaraj, G.; Pandian, S.K.; Leung, D.W.; Ramesh, M. Production of squalene with promising antioxidant properties in callus cultures of Nilgirianthus ciliatus. Ind. Crops Prod. 2018, 126, 357–367. [Google Scholar] [CrossRef]
- Kousalya, L.; Bai, V.N. Effect of growth regulators on rapid micropropagation and antioxidant activity of Canscora decussata (Roxb.) Roem. & Schult.–A threatened medicinal plant. Asian Pac. J. Reprod. 2016, 5, 161–170. [Google Scholar]
- Gul, M.Z.; Ahmad, F.; Kondapi, A.K.; Qureshi, I.A.; Ghazi, I.A. Antioxidant and antiproliferative activities of Abrus precatorius leaf extracts-an in vitro study. BMC Complement. Altern. Med. 2013, 13, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, R.; Chaurasia, J.K.; Tiwari, K.N.; Singh, K. Comparative antioxidant study of stem and stem induced callus of Phyllanthus fraternus Webster—An important antiviral and hepatoprotective plant. Appl. Biochem. Biotechnol. 2013, 171, 2153–2164. [Google Scholar] [CrossRef]
- Siswadi, S.; Saragih, G.S. Phytochemical analysis of bioactive compounds in ethanolic extract of Sterculia quadrifida R. Br. AIP Conf. Proc. 2021, 2353, 030098. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil; Agricultural Experiment Station, University of California: Berkeley, CA, USA, 1950; Volume 347. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Cheng, T.; Xu, C.; Lei, L.; Li, C.; Zhang, Y.; Zhou, S. Barcoding the kingdom Plantae: New PCR primers for ITS regions of plants with improved universality and specificity. Mol. Ecol. Resour. 2016, 16, 138–149. [Google Scholar] [CrossRef]
- Dpooležel, J.; Binarová, P.; Lcretti, S. Analysis of nuclear DNA content in plant cells by flow cytometry. Biol. Plant. 1989, 31, 113–120. [Google Scholar] [CrossRef]
- Bennett, M.D.; Smith, J. Nuclear DNA amounts in angiosperms. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1976, 274, 227–274. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Ordonez, A.; Gomez, J.; Vattuone, M. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 2006, 97, 452–458. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Material | Gallic Acid | Quercetin | Rutin |
|---|---|---|---|
| Leaves | 180.77 ± 6.17 | 73.40854 ± 3.58 | 24.13197 ± 1.26 |
| Callus | 65.7061 ± 5.58 | 48.32039 ± 4.55 | 14.39261 ± 1.33 |
| Callus Extract | Leaf Extract | ||||||
|---|---|---|---|---|---|---|---|
| Compounds | RT | % | Bioactivity | Compounds | RT | % | Bioactivity |
| Decanal | 11.00 | 2.81 | Antioxidant, antimicrobial, and antitumor [18] | Furan, 2-butyltetrahydro | 3.125 | 0.78 | Chemopreventive properties [19] |
| 2-Decenal, (E)- | 11.89 | 10.57 | Nematicidal activity [20] | 3-Hexanone | 3.309 | 2.08 | Antimicrobial [21] |
| Nonanoic acid | 12.018 | 2.42 | Antimicrobial [22] | 2-Hexanone | 3.396 | 2.65 | Antibacterial activity [23] |
| Undecanal | 12.567 | 1.73 | Antimicrobial and antioxidant activities [24] | 3-Hexanol | 3.396 | 0.68 | Antimicrobial [25] |
| 2-Undecenal | 13.414 | 11.82 | Antimicrobial and antioxidant activities [24] | 2-Hexanol | 3.587 | 1.76 | Antimicrobial [26] |
| Dodecanal | 14.029 | 2.35 | Antimicrobial [27] | Octanoic acid | 10.571 | 4.04 | Tremor suppressing [28] |
| Tridecanal | 15.396 | 1.72 | Antimicrobial [29] | Decanal | 11.015 | 2.18 | Antioxidant, antimicrobial, and antitumor [18] |
| Tetradecanal | 16.679 | 1.57 | Antioxidant and antibacterial activities [30] | 2-Decenal, (E)- | 11.914 | 10.14 | Nematicidal activity [20] |
| Pentadecanal- | 17.896 | 1.78 | Antibacterial [31] | Nonanoic acid | 12.058 | 1.69 | Antimicrobial [22] |
| Heneicosane | 18.805 | 3.44 | Antimicrobial [32] | Undecanal | 12.569 | 1.44 | Antimicrobial and antioxidant activities [24] |
| Heneicosane | 19.889 | 2.46 | Antimicrobial [32] | 2-Undecenal | 13.213 | 1.25 | Antimicrobial and antioxidant activities [24] |
| Hexadecanoic acid, methyl ester | 20.205 | 8.35 | Antibacterial activities [33] | 2-Undecenal | 13.425 | 11.19 | Antimicrobial and antioxidant activities [24] |
| n-Hexadecanoic acid | 20.613 | 17.22 | Anti-inflammatory [34] | Nonane, 2-methyl-5-propyl- | 13.835 | 1.05 | Anti-cancer [35] |
| Octadecanoic acid, 2-propenyl ester | 21.721 | 7.16 | Antibacterial [36] | Dodecanal | 14.028 | 2.03 | Antimicrobial [27] |
| 9-Octadecenoic acid, methyl ester, (E)- | 21.987 | 6.99 | Antimicrobial and nematicidal [37] | Vinyl caprylate | 14.166 | 0.71 | Antimicrobial [38] |
| 2(3H)-Furanone, 5-dodecyldihydro- | 22.105 | 2.38 | Antibacterial [39] | Trifluoroacetic acid, n-tridecyl ester | 14.568 | 0.75 | Antibacterial, antifungal [40] |
| Palmitic acid vinyl ester | 23.157 | 3.00 | Antimicrobial activity [41] | 2-Dodecenal | 14.837 | 1.01 | Nematicidal activity [20] |
| Octadecanoic acid, 2-propenyl ester | 23.591 | 3.30 | Antibacterial [36] | Pentadecane | 15.184 | 0.78 | Antimicrobial and antioxidant activity [42] |
| Heneicosane | 23.753 | 1.68 | Antimicrobial activity [32] | Tetradecanal | 15.389 | 1.47 | Antioxidant and antibacterial activities [30] |
| Pentacosane | 24.624 | 2.80 | Antimicrobial activity [43] | Tetradecanal | 16.672 | 1.64 | Antioxidant and antibacterial activities [30] |
| Tetracontane | 25.459 | 2.84 | Antioxidant and antimicrobial activity [44] | Decanoic acid, 2-propenyl ester | 17.416 | 0.97 | Antimicrobial activity [45] |
| Tetratriacontane | 26.269 | 1.61 | Antibacterial and antifungal [46] | Pentadecanal | 17.886 | 2.17 | Antibacterial (Ricciardelli, et al., 2020) |
| Hexadecanoic acid, methyl ester | 20.195 | 2.74 | Antibacterial activities [33] | ||||
| n-Hexadecanoic acid | 20.615 | 1.28 | Anti-inflammatory [34] | ||||
| Octadecanoic acid, 2-propenyl ester | 21.711 | 21.77 | Antibacterial [36] | ||||
| Palmitic acid vinyl ester | 23.151 | 8.83 | Antimicrobial activity [41] | ||||
| Octadecanoic acid, 2-propenyl ester | 23.579 | 10.60 | Antibacterial [36] | ||||
| Bis(2-ethylhexyl) phthalate | 25.985 | 2.33 | Antimicrobial and cytotoxic activity [47] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Qurainy, F.; Tarroum, M.; Khan, S.; Nadeem, M.; Gaafar, A.-R.Z.; Alansi, S.; Alfarraj, N.S. Genome Estimation and Phytochemical Compound Identification in the Leaves and Callus of Abrus precatorius: A Locally Endangered Plant from the Flora of Saudi Arabia. Plants 2022, 11, 567. https://doi.org/10.3390/plants11040567
Al-Qurainy F, Tarroum M, Khan S, Nadeem M, Gaafar A-RZ, Alansi S, Alfarraj NS. Genome Estimation and Phytochemical Compound Identification in the Leaves and Callus of Abrus precatorius: A Locally Endangered Plant from the Flora of Saudi Arabia. Plants. 2022; 11(4):567. https://doi.org/10.3390/plants11040567
Chicago/Turabian StyleAl-Qurainy, Fahad, Mohamed Tarroum, Salim Khan, Mohammad Nadeem, Abdel-Rhman Z. Gaafar, Saleh Alansi, and Norah S. Alfarraj. 2022. "Genome Estimation and Phytochemical Compound Identification in the Leaves and Callus of Abrus precatorius: A Locally Endangered Plant from the Flora of Saudi Arabia" Plants 11, no. 4: 567. https://doi.org/10.3390/plants11040567