
Cereal Germination under Low Oxygen: Molecular Processes


Abstract
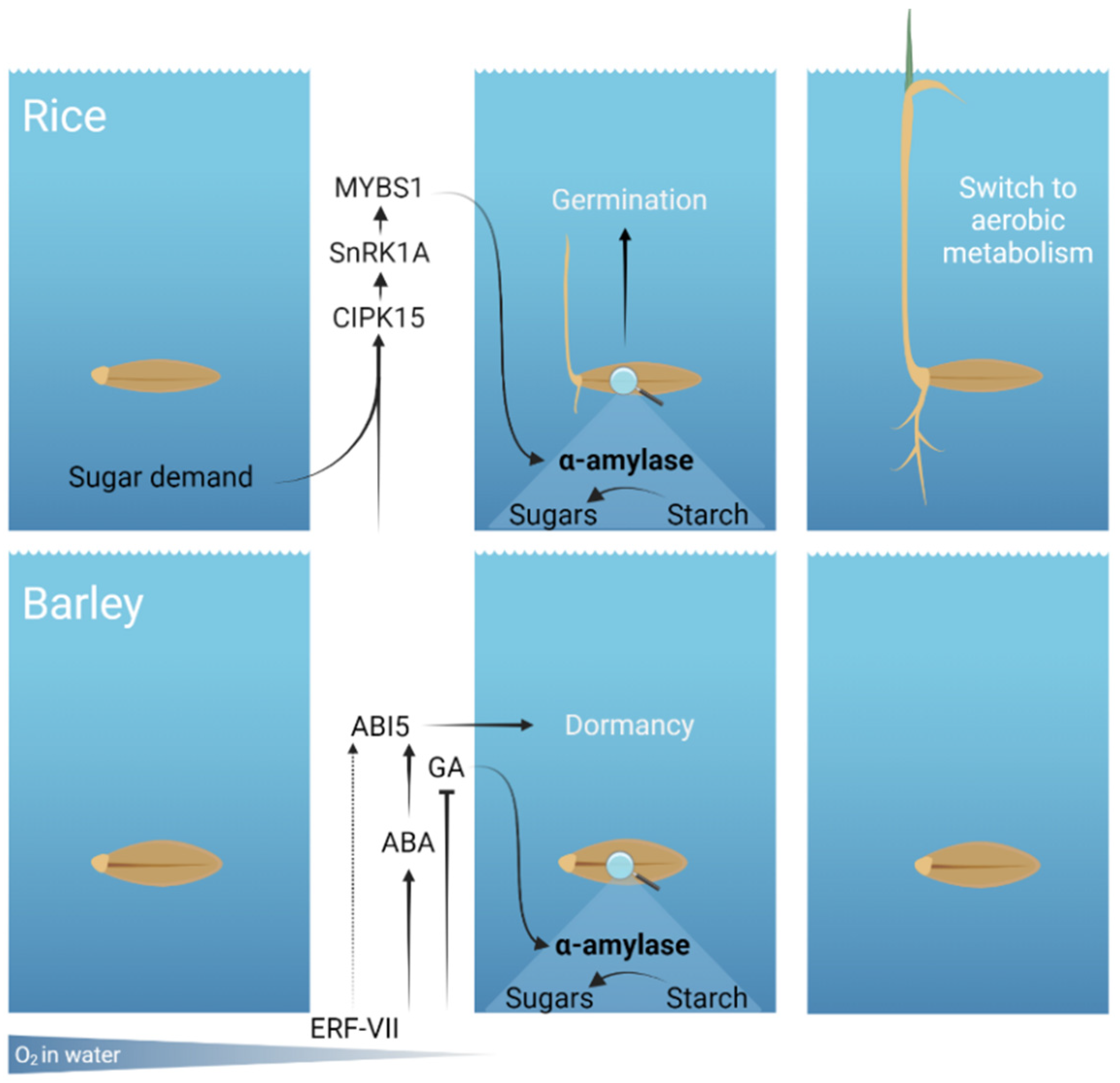
:1. Introduction
2. Rice Germinates under Anoxia and Submergence
2.1. Rice Molecular Mechanism Finalized to Starch Degradation under Low Oxygen
2.2. Rice Coleoptile Elongation under Low Oxygen
3. Barley Is Unable to Germinate under Anoxia and Prolonged Submergence
3.1. Hypoxia Affects Hormonal Regulation in Barley Grains
3.2. Molecular Mechanisms Regulating Barley Sensitivity to Low Oxygen
4. Wheat Response to Anoxia and Submergence during Seed Germination
5. The N-Degron Pathway for Low Oxygen Sensing during Germination
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bassu, S.; Asseng, S.; Motzo, R.; Giunta, F. Optimising sowing date of durum wheat in a variable Mediterranean environment. F. Crop. Res. 2009, 111, 109–118. [Google Scholar] [CrossRef]
- Cho, H.Y.; Loreti, E.; Shih, M.C.; Perata, P. Energy and sugar signaling during hypoxia. New Phytol. 2021, 229, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Voesenek, L.A.C.J.; Bailey-Serres, J. Flood adaptive traits and processes: An overview. New Phytol. 2015, 206, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Geigenberger, P. Response of plant metabolism to too little oxygen. Curr. Opin. Plant Biol. 2003, 6, 247–256. [Google Scholar] [CrossRef]
- Van Dongen, J.T.; Licausi, F. Oxygen sensing and signaling. Annu. Rev. Plant Biol. 2015, 66, 345–367. [Google Scholar] [CrossRef]
- Shukla, V.; Lombardi, L.; Iacopino, S.; Pencik, A.; Novak, O.; Perata, P.; Giuntoli, B.; Licausi, F. Endogenous hypoxia in lateral root primordia controls root architecture by antagonizing auxin signaling in Arabidopsis. Mol. Plant 2019, 12, 538–551. [Google Scholar] [CrossRef] [Green Version]
- Weits, D.A.; Kunkowska, A.B.; Kamps, N.C.W.; Portz, K.M.S.; Packbier, N.K.; Nemec Venza, Z.; Gaillochet, C.; Lohmann, J.U.; Pedersen, O.; van Dongen, J.T.; et al. An apical hypoxic niche sets the pace of shoot meristem activity. Nature 2019, 569, 714–717. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [Green Version]
- Borisjuk, L.; Rolletschek, H. The oxygen status of the developing seed. New Phytol. 2009, 182, 17–30. [Google Scholar] [CrossRef]
- Larson, L.A. The effect soaking pea seeds with or without seed coats has on seedling growth. Plant Physiol. 1968, 43, 255–259. [Google Scholar] [CrossRef]
- Yu, S.-M.; Lo, S.-F.; Ho, T.-H.D. Source-sink communication: Regulated by hormone, nutrient, and stress cross-signaling. Trends Plant Sci. 2015, 20, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Alpi, A.; Beevers, H. Effect of O2 concentration on rice seedlings. Plant Physiol. 1983, 71, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Perata, P.; Matsukura, C.; Vernieri, P.; Yamaguchi, J. Sugar repression of a gibberellin-dependent signaling pathway in bar ley embryos. Plant Cell. 1997, 9, 2197–2208. [Google Scholar] [CrossRef]
- Guglielminetti, L.; Perata, P.; Alpi, A. Effect of anoxia on carbohydrate metabolism in rice seedlings. Plant Physiol. 1995, 108, 735–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guglielminetti, L.; Yamaguchi, J.; Perata, P.; Alpi, A. Amylolytic activities in cereal seeds under aerobic and anaerobic conditions. Plant Physiol. 1995, 109, 1069–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.S.; Karrer, E.E.; Thomas, B.R.; Chen, L.; Rodriguez, R.L. Three cis-elements required for rice alpha-amylase Amy3D expression during sugar starvation. Plant Mol. Biol. 1998, 36, 331–341. [Google Scholar] [CrossRef]
- Hwang, Y.-S.; Thomas, B.R.; Rodriguez, R.L. Differential expression of rice α-amylase genes during seedling development under anoxia. Plant Mol. Biol. 1999, 40, 911–920. [Google Scholar] [CrossRef]
- Lee, K.-W.; Chen, P.W.; Yu, S.-M. Metabolic adaptation to sugar/O2 deficiency for anaerobic germination and seedling growth in rice. Plant. Cell Environ. 2014, 37, 2234–2244. [Google Scholar] [CrossRef]
- Miro, B.; Ismail, A.M. Tolerance of anaerobic conditions caused by flooding during germination and early growth in rice (Oryza sativa L.). Front. Plant Sci. 2013, 4, 269. [Google Scholar] [CrossRef] [Green Version]
- Fincher, G.B. Molecular and cellular biology associated with endosperm mobilization in germinating cereal grains. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1989, 40, 305–346. [Google Scholar] [CrossRef]
- Lu, C.-A.; Ho, T.D.; Ho, S.-L.; Yu, S.-M. Three novel MYB proteins with one DNA binding repeat mediate sugar and hormone regulation of alpha-amylase gene expression. Plant Cell 2002, 14, 1963–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.-A.; Lin, C.-C.; Lee, K.-W.; Chen, J.-L.; Huang, L.-F.; Ho, S.-L.; Liu, H.-J.; Hsing, Y.-I.; Yu, S.-M. The SnRK1A protein kinase plays a key role in sugar signaling during germination and seedling growth of rice. Plant Cell 2007, 19, 2484–2499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubler, F.; Kalla, R.; Roberts, J.K.; Jacobsen, J.V. Gibberellin-regulated expression of a myb gene in barley aleurone cells: Evidence for Myb transactivation of a high-pI alpha-amylase gene promoter. Plant Cell 1995, 7, 1879–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubler, F.; Watts, R.J.; Kalla, R.; Matthews, P.; Keys, M.; Jacobsen, J.V. Cloning of a rice cDNA encoding a transcription factor homologous to barley GAMyb. Plant Cell Physiol. 1997, 38, 362–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, J.V.; Gubler, F.; Chandler, P.M. Gibberellin action in germinated cereal grains. In Plant Hormones: Physiology, Biochemistry and Molecular Biology; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 1995; pp. 246–271. ISBN 978-94-011-0473-9. [Google Scholar]
- Loreti, E.; Yamaguchi, J.; Alpi, A.; Perata, P. Gibberellins are not required for rice germination under anoxia. Plant Soil 2003, 253, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P.; Kamiya, Y. Gibberellin biosynthesis: Enzymes, genes and their regulation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 431–460. [Google Scholar] [CrossRef] [Green Version]
- Iacopino, S.; Licausi, F. The contribution of plant dioxygenases to hypoxia signaling. Front. Plant Sci. 2020, 11, 1008. [Google Scholar] [CrossRef]
- Lee, K.-W.; Chen, P.-W.; Lu, C.-A.; Chen, S.; Ho, T.-H.D.; Yu, S.-M. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Sci. Signal. 2009, 2, ra61. [Google Scholar] [CrossRef]
- Chaves-Sanjuan, A.; Sanchez-Barrena, M.J.; Gonzalez-Rubio, J.M.; Moreno, M.; Ragel, P.; Jimenez, M.; Pardo, J.M.; Martinez-Ripoll, M.; Quintero, F.J.; Albert, A. Structural basis of the regulatory mechanism of the plant CIPK family of protein kinases controlling ion homeostasis and abiotic stress. Proc. Natl. Acad. Sci. USA 2014, 111, E4532. [Google Scholar] [CrossRef] [Green Version]
- Ho, V.T.; Tran, A.N.; Cardarelli, F.; Perata, P.; Pucciariello, C. A calcineurin B-like protein participates in low oxygen signalling in rice. Funct. Plant Biol. 2017, 44, 917–928. [Google Scholar] [CrossRef]
- Ye, N.-H.; Wang, F.-Z.; Shi, L.; Chen, M.-X.; Cao, Y.-Y.; Zhu, F.-Y.; Wu, Y.-Z.; Xie, L.-J.; Liu, T.-Y.; Su, Z.-Z.; et al. Natural variation in the promoter of rice calcineurin B-like protein10 (OsCBL10) affects flooding tolerance during seed germination among rice subspecies. Plant J. 2018, 94, 612–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-S.; David Ho, T.-H.; Liu, L.; Lee, D.H.; Lee, C.-H.; Chen, Y.-R.; Lin, S.-Y.; Lu, C.-A.; Yu, S.-M. Sugar starvation-regulated MYBS2 and 14-3-3 protein interactions enhance plant growth, stress tolerance, and grain weight in rice. Proc. Natl. Acad. Sci. USA 2019, 116, 21925–21935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angaji, S.A.; Septiningsih, E.M.; Mackill, D.J.; Abdelbagi, M.I. QTLs associated with tolerance of flooding during germination in rice (Oryza sativa L.). Euphytica 2010, 172, 159–168. [Google Scholar] [CrossRef]
- Kretzschmar, T.; Pelayo, M.A.F.; Trijatmiko, K.R.; Gabunada, L.F.M.; Alam, R.; Jimenez, R.; Mendioro, M.S.; Slamet-Loedin, I.H.; Sreenivasulu, N.; Bailey-Serres, J.; et al. A trehalose-6-phosphate phosphatase enhances anaerobic germination tolerance in rice. Nat. Plants 2015, 1, 15124. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.J. Trehalose 6-phosphate: A signal of sucrose status. Biochem. J. 2008, 412, e1–e2. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Singh, P.; Kaladhar, V.C.; Manbir; Paul, D.; Pathak, P.K.; Gupta, K.J. Phytoglobin-NO cycle and AOX pathway play a role in anaerobic germination and growth of deepwater rice. Plant. Cell Environ. 2021, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Hill, R.D. Nitrate, NO and haemoglobin in plant adaptation to hypoxia: An alternative to classic fermentation pathways. J. Exp. Bot. 2004, 55, 2473–2482. [Google Scholar] [CrossRef] [Green Version]
- Nghi, K.N.; Tondelli, A.; Valè, G.; Tagliani, A.; Marè, C.; Perata, P.; Pucciariello, C. Dissection of coleoptile elongation in japonica rice under submergence through integrated genome-wide association mapping and transcriptional analyses. Plant. Cell Environ. 2019, 42, 1832–1846. [Google Scholar] [CrossRef]
- Kawai, M.; Uchimiya, H. Coleoptile Senescence in Rice (Oryza sativa L.). Ann. Bot. 2000, 86, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Nghi, K.N.; Tagliani, A.; Mariotti, L.; Weits, D.A.; Perata, P.; Pucciariello, C. Auxin is required for the long coleoptile trait in japonica rice under submergence. New Phytol. 2021, 229, 85–93. [Google Scholar] [CrossRef]
- Guo, F.; Han, N.; Xie, Y.; Fang, K.; Yang, Y.; Zhu, M.; Wang, J.; Bian, H. The miR393a/target module regulates seed germination and seedling establishment under submergence in rice (Oryza sativa L.). Plant. Cell Environ. 2016, 39, 2288–2302. [Google Scholar] [CrossRef]
- Si-Ammour, A.; Windels, D.; Arn-Bouldoires, E.; Kutter, C.; Ailhas, J.; Meins, F.J.; Vazquez, F. miR393 and secondary siRNAs regulate expression of the TIR1/AFB2 auxin receptor clade and auxin-related development of Arabidopsis leaves. Plant Physiol. 2011, 157, 683–691. [Google Scholar] [CrossRef] [Green Version]
- Perata, P.; Guglielminetti, L.; Alpi, A. Anaerobic carbohydrate metabolism in wheat and barley, two anoxia-intolerant cereal seeds. J. Exp. Bot. 1996, 47, 999–1006. [Google Scholar] [CrossRef]
- Arduini, I.; Orlandi, O.; Ercoli, L.; Masoni, A. Submergence sensitivity of durum wheat, bread wheat and barley at the germination stage. Ital. J. Agron. 2016, 11, 100–106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, C. Comparisons of copy number, genomic structure, and conserved motifs for α-amylase genes from barley, rice, and wheat. Front. Plant Sci. 2017, 8, 1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, H.H.; Bailly, C.; Corbineau, F.; Leymarie, J. Induction of secondary dormancy by hypoxia in barley grains and its hormonal regulation. J. Exp. Bot. 2013, 64, 2017–2025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, M.E. Seed dormancy: An update on terminology, physiological genetics, and quantitative trait loci regulating germinability. Weed Sci. 2001, 49, 305–317. [Google Scholar] [CrossRef]
- Hilhorst, H. Definitions and hypothesis of seed dormancy. In Seed Development, Dormancy and Germination; Bradford, K.J., Nonogaki, H., Eds.; Oxford Blackwell Publishing: Oxford, UK, 2007; pp. 50–71. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L.; Gualano, N.; Leymarie, J.; Côme, D.; Corbineau, F. Hypoxia interferes with ABA metabolism and increases ABA sensitivity in embryos of dormant barley grains. J. Exp. Bot. 2006, 57, 1423–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benech-Arnold, R.L.; Giallorenzi, C.M.; Frank, J.; Rodriguez, V. Termination of hull-imposed dormancy in developing barley grains is correlated with changes in embryonic ABA levels and sensitivity. Seed Sci. Res. 1999, 9, 39–47. [Google Scholar] [CrossRef]
- Bradford, K.J.; Benech-Arnold, R.L.; Côme, D.; Corbineau, F. Quantifying the sensitivity of barley seed germination to oxygen, abscisic acid, and gibberellin using a population-based threshold model. J. Exp. Bot. 2008, 59, 335–347. [Google Scholar] [CrossRef]
- Lenoir, C.; Corbineau, F.; Côme, D. Barley (Hordeum vulgare) seed dormancy as related to glumella characteristics. Physiol. Plant. 1986, 68, 301–307. [Google Scholar] [CrossRef]
- Corbineau, F.S.; Lecat, D.C. Dormancy of three cultivars of oat seeds (Avena sativa L.). Seed Sci. Technol. 1986, 14, 725–735. [Google Scholar]
- Mendiondo, G.M.; Leymarie, J.; Farrant, J.M.; Corbineau, F.; Benech-Arnold, R.L. Differential expression of abscisic acid metabolism and signalling genes induced by seed-covering structures or hypoxia in barley (Hordeum vulgare L.) grains. Seed Sci. Res. 2010, 20, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Zafari, S.; Hebelstrup, K.H.; Igamberdiev, A.U. Transcriptional and metabolic changes associated with phytoglobin expression during germination of barley seeds. Int. J. Mol. Sci. 2020, 21, 2796. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Marsolais, F.; Bykova, N.V.; Igamberdiev, A.U. Nitric oxide and reactive oxygen species mediate metabolic changes in barley seed embryo during germination. Front. Plant Sci. 2016, 7, 138. [Google Scholar] [CrossRef] [Green Version]
- Bethke, P.C.; Gubler, F.; Jacobsen, J.V.; Jones, R.L. Dormancy of Arabidopsis seeds and barley grains can be broken by nitric oxide. Planta 2004, 219, 847–855. [Google Scholar] [CrossRef]
- Cochrane, D.W.; Shah, J.K.; Hebelstrup, K.H.; Igamberdiev, A.U. Expression of phytoglobin affects nitric oxide metabolism and energy state of barley plants exposed to anoxia. Plant Sci. 2017, 265, 124–130. [Google Scholar] [CrossRef]
- Ishibashi, Y.; Tawaratsumida, T.; Kondo, K.; Kasa, S.; Sakamoto, M.; Aoki, N.; Zheng, S.-H.; Yuasa, T.; Iwaya-Inoue, M. Reactive oxygen species are involved in gibberellin/abscisic acid signaling in barley aleurone cells. Plant Physiol. 2012, 158, 1705–1714. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, Y.; Kasa, S.; Sakamoto, M.; Aoki, N.; Kai, K.; Yuasa, T.; Hanada, A.; Yamaguchi, S.; Iwaya-Inoue, M. A Role for reactive oxygen species produced by NADPH oxidases in the embryo and aleurone cells in barley seed germination. PLoS ONE 2015, 10, e0143173. [Google Scholar] [CrossRef]
- De Sousa, C.A.F.; Sodek, L. Alanine metabolism and alanine aminotransferase activity in soybean (Glycine max) during hypoxia of the root system and subsequent return to normoxia. Environ. Exp. Bot. 2003, 50, 1–8. [Google Scholar] [CrossRef]
- Miyashita, Y.; Dolferus, R.; Ismond, K.P.; Good, A.G. Alanine aminotransferase catalyses the breakdown of alanine after hypoxia in Arabidopsis thaliana. Plant J. 2007, 49, 1108–1121. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, J.T.; Fröhlich, A.; Ramírez-Aguilar, S.J.; Schauer, N.; Fernie, A.R.; Erban, A.; Kopka, J.; Clark, J.; Langer, A.; Geigenberger, P. Transcript and metabolite profiling of the adaptive response to mild decreases in oxygen concentration in the roots of Arabidopsis plants. Ann. Bot. 2009, 103, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Rocha, M.; Sodek, L.; Licausi, F.; Hameed, M.W.; Dornelas, M.C.; van Dongen, J.T. Analysis of alanine aminotransferase in various organs of soybean (Glycine max) and in dependence of different nitrogen fertilisers during hypoxic stress. Amino Acids 2010, 39, 1043–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, M.; Licausi, F.; Araújo, W.L.; Nunes-Nesi, A.; Sodek, L.; Fernie, A.R.; van Dongen, J.T. Glycolysis and the tricarboxylic acid cycle are linked by alanine aminotransferase during hypoxia induced by waterlogging of Lotus japonicus. Plant Physiol. 2010, 152, 1501–1513. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Yamane, M.; Yamaji, N.; Kanamori, H.; Tagiri, A.; Schwerdt, J.G.; Fincher, G.B.; Matsumoto, T.; Takeda, K.; Komatsuda, T. Alanine aminotransferase controls seed dormancy in barley. Nat. Commun. 2016, 7, 11625. [Google Scholar] [CrossRef]
- Perata, P.; Geshi, N.; Yamaguchi, J.; Akazawa, T. Effect of anoxia on the induction of α-amylase in cereal seeds. Planta 1993, 191, 402–408. [Google Scholar] [CrossRef]
- Perata, P.; Pozueta-Romero, J.; Akazawa, T.; Yamaguchi, J. Effect of anoxia on starch breakdown in rice and wheat seeds. Planta. 1992, 188, 611–618. [Google Scholar] [CrossRef]
- Guglielminetti, L.; Busilacchi, H.A.; Perata, P.; Alpi, A. Carbohydrate-ethanol transition in cereal grains under anoxia. New Phytol. 2001, 151, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Yan, M.; Xue, C.; Xiong, Y.; Meng, X.; Li, B.; Shen, R.; Lan, P. Proteomic dissection of the similar and different responses of wheat to drought, salinity and submergence during seed germination. J. Proteom. 2020, 220, 103756. [Google Scholar] [CrossRef]
- Yan, M.; Zheng, L.; Li, B.; Shen, R.; Lan, P. Comparative proteomics reveals new insights into the endosperm responses to drought, salinity and submergence in germinating wheat seeds. Plant Mol. Biol. 2021, 105, 287–302. [Google Scholar] [CrossRef]
- Shen, C.; Yuan, J.; Qiao, H.; Wang, Z.; Liu, Y.; Ren, X.; Wang, F.; Liu, X.; Zhang, Y.; Chen, X.; et al. Transcriptomic and anatomic profiling reveal the germination process of different wheat varieties in response to waterlogging stress. BMC Genet. 2020, 21, 93. [Google Scholar] [CrossRef]
- Gibbs, D.J.; Conde, J.V.; Berckhan, S.; Prasad, G.; Mendiondo, G.M.; Holdsworth, M.J. Group VII ethylene response factors coordinate oxygen and nitric oxide signal transduction and stress responses in plants. Plant Physiol. 2015, 169, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Weits, D.A.; Giuntoli, B.; Kosmacz, M.; Parlanti, S.; Hubberten, H.-M.; Riegler, H.; Hoefgen, R.; Perata, P.; van Dongen, J.T.; Licausi, F. Plant cysteine oxidases control the oxygen-dependent branch of the N-end-rule pathway. Nat. Commun. 2014, 5, 3425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.D.; Klecker, M.; Hopkinson, R.J.; Weits, D.A.; Mueller, C.; Naumann, C.; O’Neill, R.; Wickens, J.; Yang, J.; Brooks-Bartlett, J.C.; et al. Plant cysteine oxidases are dioxygenases that directly enable arginyl transferase-catalysed arginylation of N-end rule targets. Nat. Commun. 2017, 8, 14690. [Google Scholar] [CrossRef]
- Licausi, F.; Kosmacz, M.; Weits, D.; Giuntoli, B.; Giorgi, F.M.; Voesenek, L.A.C.J.; Perata, P.; van Dongen, J.T. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature 2011, 479, 419–422. [Google Scholar] [CrossRef]
- Gibbs, D.J.; Lee, S.C.; Md Isa, N.; Gramuglia, S.; Fukao, T.; Bassel, G.W.; Correia, C.S.; Corbineau, F.; Theodoulou, F.L.; Bailey-Serres, J.; et al. Homeostatic response to hypoxia is regulated by the N-end rule pathway in plants. Nature 2011, 479, 415–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, D.J.; Md Isa, N.; Movahedi, M.; Lozano-Juste, J.; Mendiondo, G.M.; Berckhan, S.; Marín-de la Rosa, N.; Vicente Conde, J.; Sousa Correia, C.; Pearce, S.P.; et al. Nitric oxide sensing in plants is mediated by proteolytic control of group VII ERF transcription factors. Mol. Cell 2014, 53, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarban, R.; Vogler, M.; Wong, A.; Eppinger, J.; Al-Babili, S.; Gehring, C. Discovery of a nitric oxide-responsive protein in Arabidopsis thaliana. Molecules 2019, 24, 2691. [Google Scholar] [CrossRef] [Green Version]
- Holman, T.J.; Jones, P.D.; Russell, L.; Medhurst, A.; Úbeda Tomás, S.; Talloji, P.; Marquez, J.; Schmuths, H.; Tung, S.-A.; Taylor, I.; et al. The N-end rule pathway promotes seed germination and establishment through removal of ABA sensitivity in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 4549–4554. [Google Scholar] [CrossRef] [Green Version]
- Mendiondo, G.M.; Gibbs, D.J.; Szurman-Zubrzycka, M.; Korn, A.; Marquez, J.; Szarejko, I.; Maluszynski, M.; King, J.; Axcell, B.; Smart, K.; et al. Enhanced waterlogging tolerance in barley by manipulation of expression of the N-end rule pathway E3 ligase proteolysis 6. Plant Biotechnol. J. 2016, 14, 40–50. [Google Scholar] [CrossRef]
- Lin, C.C.; Chao, Y.T.; Chen, W.C.; Ho, H.Y.; Chou, M.Y.; Li, Y.R.; Wu, Y.L.; Yang, H.A.; Hsieh, H.; Lin, C.S.; et al. Regulatory cascade involving transcriptional and N-end rule pathways in rice under submergence. Proc. Natl. Acad. Sci. USA 2019, 116, 3300–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Molecular and Physiological Modification | Rice | Barley |
|---|---|---|
| Starch use | Increased expression and activity of α-amylase under anoxia [15,17] | No α-amylase expression or activity was detected under anoxia [15] |
| ABA-GA balance | Activation of a GA-independent signal under anoxia [26] | Expression of genes involved in GA inactivation and repression of genes involved in GA synthesis under hypoxia [47] |
| Pgb/NO cycle | In deepwater rice, Pgb/NO cycle contributes to ATP generation under anoxia [37] | The over-expression of Pgb1 supports germination under hypoxia [59] |
| N-degron pathway | - | HvPRT6 is involved in seed germination under hypoxia [82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Álvarez, E.M.; Pucciariello, C. Cereal Germination under Low Oxygen: Molecular Processes. Plants 2022, 11, 460. https://doi.org/10.3390/plants11030460
Gómez-Álvarez EM, Pucciariello C. Cereal Germination under Low Oxygen: Molecular Processes. Plants. 2022; 11(3):460. https://doi.org/10.3390/plants11030460
Chicago/Turabian StyleGómez-Álvarez, Eva María, and Chiara Pucciariello. 2022. "Cereal Germination under Low Oxygen: Molecular Processes" Plants 11, no. 3: 460. https://doi.org/10.3390/plants11030460