
Matching of Nitrogen Enhancement and Photosynthetic Efficiency by Arbuscular Mycorrhiza in Maize (Zea mays L.) in Relation to Organic Fertilizer Type


Abstract
:1. Introduction
2. Results
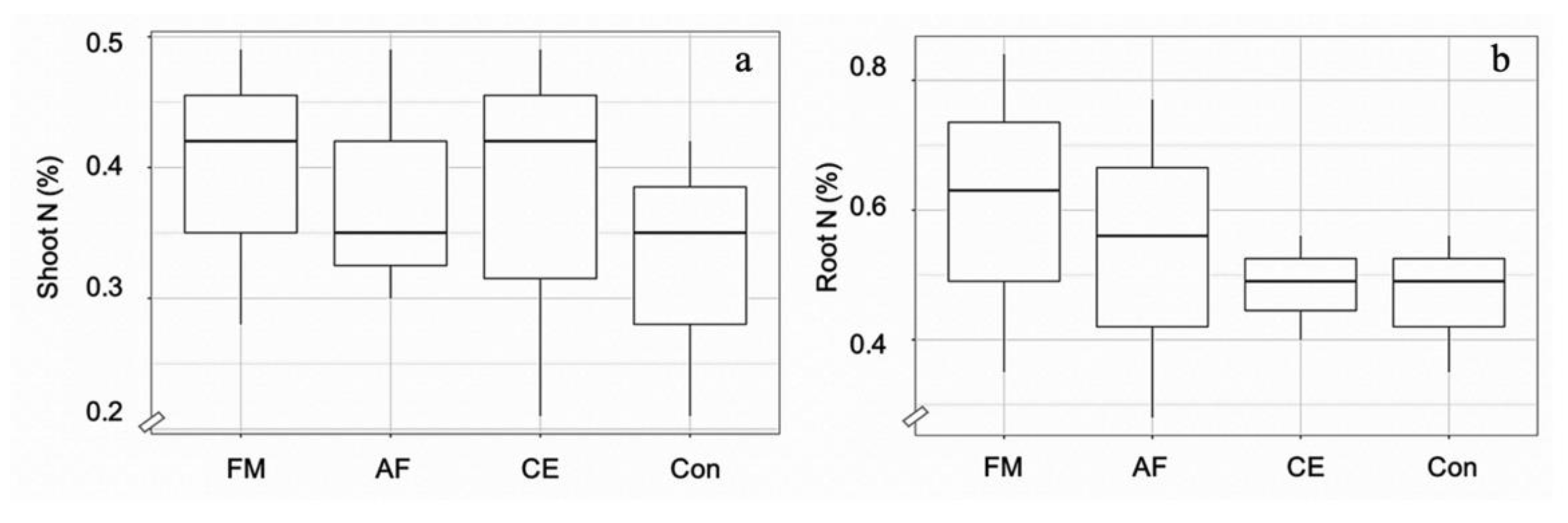
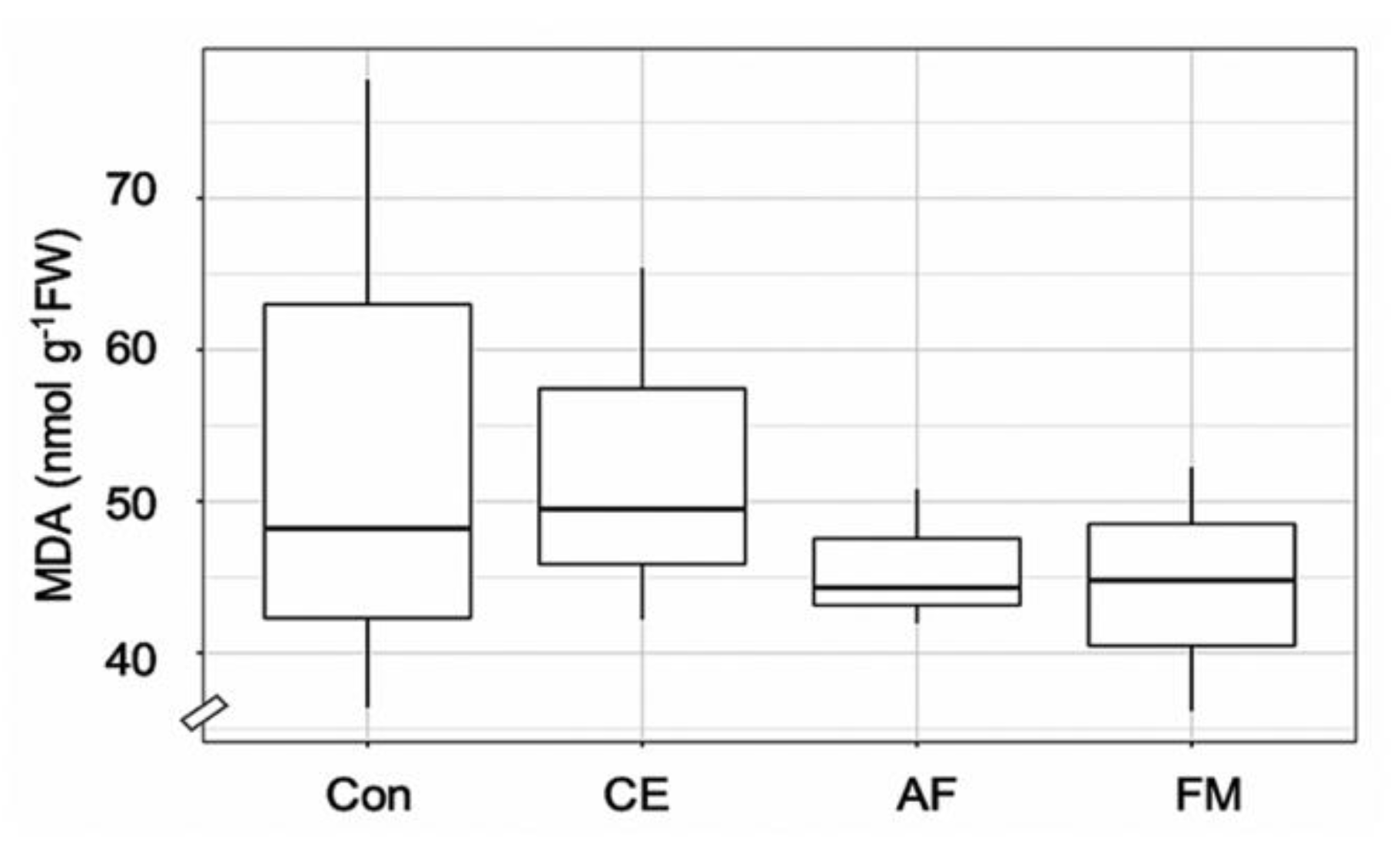
2.1. AMF Colonization, N Content, and Plant Adaptation
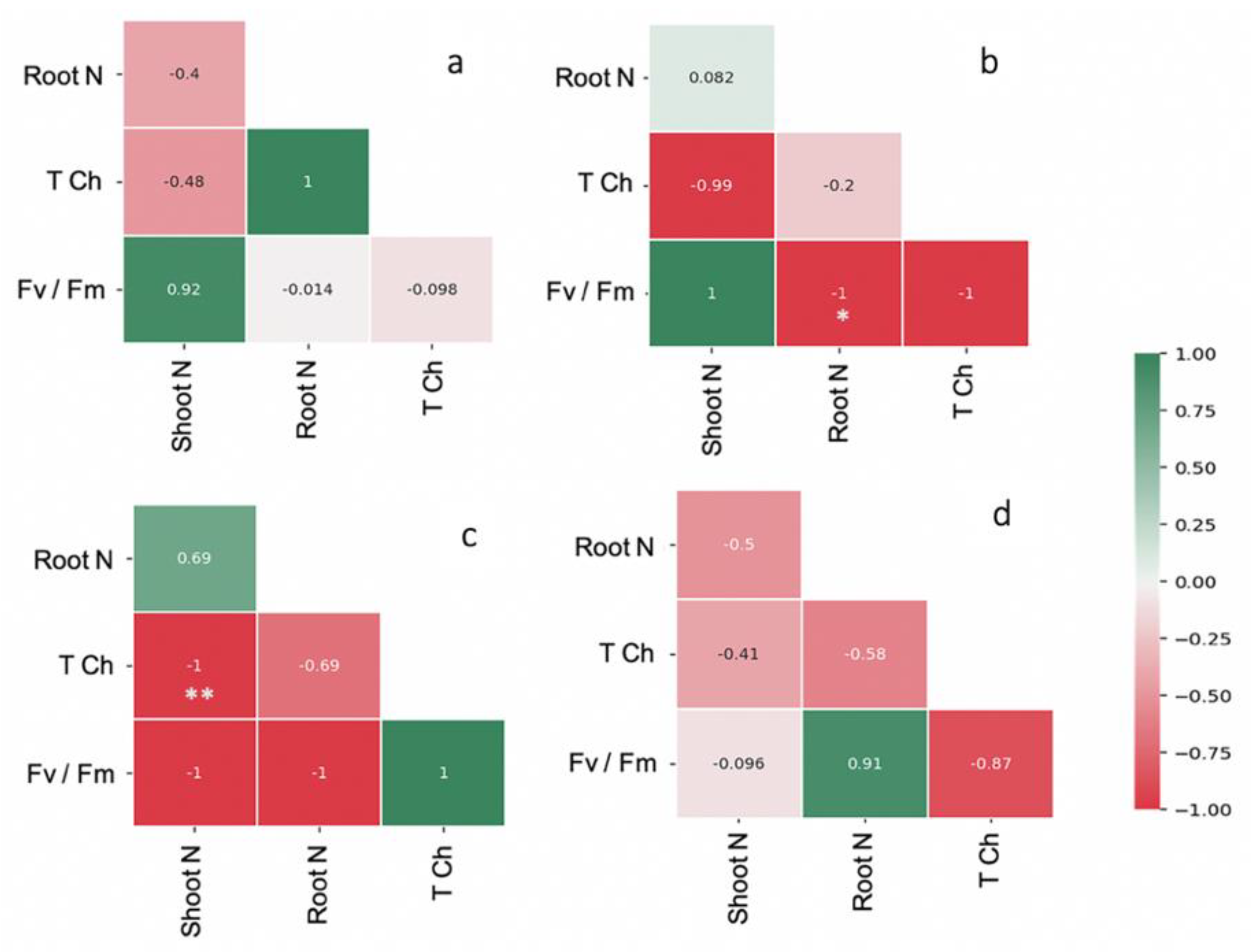
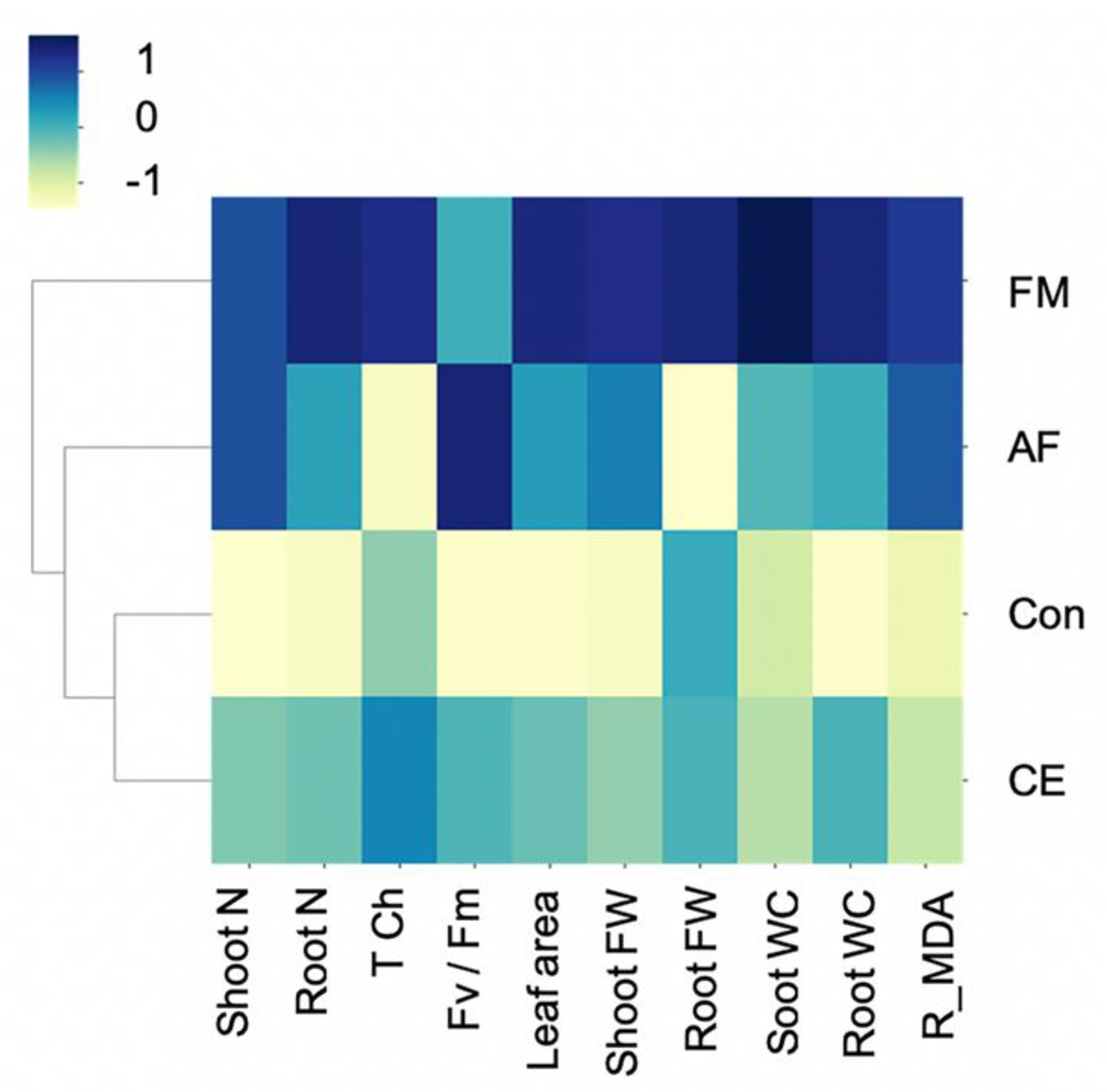
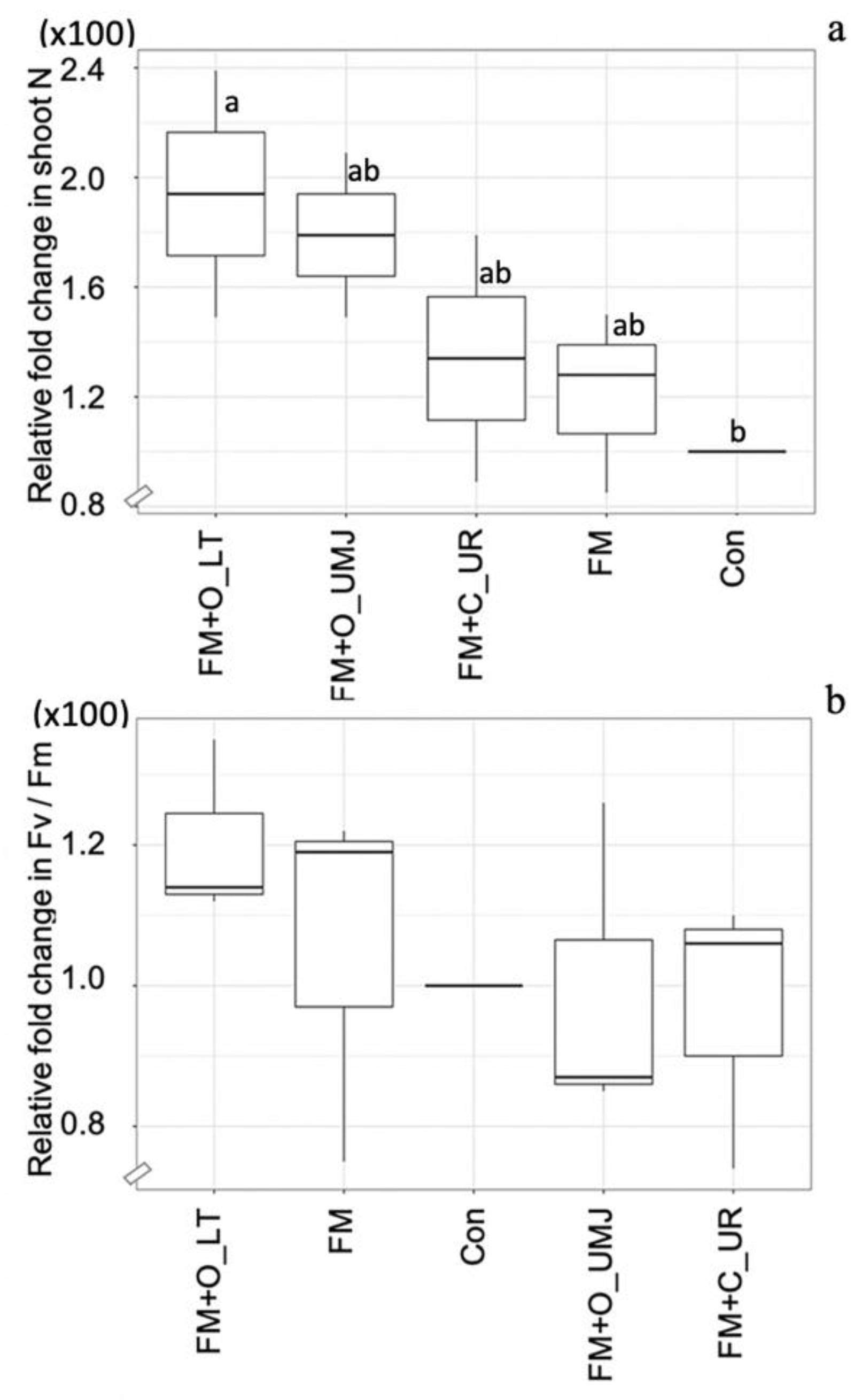
2.2. AMF Modulated N Improvement in Maize Plants
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. AMF Colonization Analysis
4.3. Total N Determination
4.4. Determination of Chlorophyll Content, Photosynthetic Efficiency, and Growth Character
4.5. Malondialdehyde Analysis
4.6. Experimental Layout and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mudau, N.F.; Soundy, P.; Du Toit, E.S. Plant growth and development of bush tea as affected by nitrogen, phosphorus, and potassium nutrition. HortScience 2005, 40, 1898–1901. [Google Scholar] [CrossRef]
- Ye, T.; Li, Y.; Zhang, J.; Hou, W.; Zhou, W.; Lu, J.; Xing, Y.; Li, X. Nitrogen, phosphorus, and potassium fertilization affects the flowering time of rice (Oryza sativa L.). Glob. Ecol. Conserv. 2019, 20, e00753. [Google Scholar] [CrossRef]
- Cao, P.; Lu, C.; Yu, Z. Historical nitrogen fertilizer use in agricultural ecosystems of the contiguous United States during 1850–2015: Application rate, timing, and fertilizer types. Earth Syst. Sci. Data 2018, 10, 969–984. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.M.; Messina, C.D.; Vyn, T.J. Simultaneous gains in grain yield and nitrogen efficiency over 70 years of maize genetic improvement. Sci. Rep. 2019, 9, 9095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, W.M.; Dibb, D.W.; Johnston, A.E.; Smyth, T.J. The contribution of commercial fertilizer nutrients to food production. Agron. J. 2005, 97, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Lu, D.; Zu, C.; Shen, J.; Wang, S.; Guo, Z.; Zhou, J.; Wang, H. One-time root-zone N fertilization increases maize yield, NUE and reduces soil N losses in lime concretion black soil. Sci. Rep. 2018, 8, 10258. [Google Scholar] [CrossRef]
- Jiang, C.; Lu, D.; Zu, C.; Zhou, J.; Wang, H. Root-zone fertilization improves crop yields and minimizes nitrogen loss in summer maize in China. Sci. Rep. 2018, 8, 15139. [Google Scholar] [CrossRef] [Green Version]
- Fu, W.; Wang, Y.; Ye, Y.; Zhen, S.; Zhou, B.; Wang, Y.; Hu, Y.; Zhao, Y.; Huang, Y. Grain yields and nitrogen use efficiencies in different types of stay-green maize in response to nitrogen fertilizer. Plants 2020, 9, 474. [Google Scholar] [CrossRef]
- Wang, X.X.; Wang, X.; Sun, Y.; Cheng, Y.; Liu, S.; Chen, X.; Feng, G.; Kuyper, T.W. Arbuscular mycorrhizal fungi negatively affect nitrogen acquisition and grain yield of maize in a N deficient soil. Front. Microbiol. 2018, 9, 418. [Google Scholar] [CrossRef] [Green Version]
- Berruti, A.; Lumini, E.; Balestrini, R.; Bianciotto, V. Arbuscular mycorrhizal fungi as natural biofertilizers: Let’s benefit from past successes. Front. Microbiol. 2016, 6, 1559. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Thokchom, S.D.; Kapoor, R. Arbuscular mycorrhiza improves photosynthesis and restores alteration in sugar metabolism in Triticum aestivum L. grown in arsenic contaminated soil. Front. Plant Sci. 2021, 12, 640379. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Wang, X.X.; Feng, G. Optimised nitrogen fertiliser management achieved higher diversity of arbuscular mycorrhiza fungi and high-yielding maize (Zea mays L.). Crop Pasture Sci. 2015, 66, 706–714. [Google Scholar] [CrossRef]
- Yooyongwech, S.; Phaukinsang, N.; Cha-um, S.; Supaibulwatana, K. Arbuscular mycorrhiza improved growth performance in Macadamia tetraphylla L. grown under water deficit stress involves soluble sugar and proline accumulation. Plant Growth Regul. 2013, 69, 285–293. [Google Scholar] [CrossRef]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Büking, H.; Lammers, P.J.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef]
- Ingraffia, R.; Amato, G.; Sosa-Hernández, M.A.; Frenda, A.S.; Rillig, M.C.; Giambalvo, D. Nitrogen type and availability drive mycorrhizal effects on wheat performance, nitrogen uptake and recovery, and production sustainability. Front. Plant Sci. 2020, 11, 760. [Google Scholar] [CrossRef]
- Huang, G.M.; Zou, Y.N.; Wu, Q.S.; Xu, Y.J.; Kuča, K. Mycorrhizal roles in plant growth, gas exchange, root morphology, and nutrient uptake of walnuts. Plant Soil Environ. 2020, 66, 295–302. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, E.B.; Egerton-Warburton, L.M.; Hart, M.M.; Klironomos, J.N.; Maherali, H.; Tedersoo, L. Arbuscular mycorrhizal fungi as mediators of ecosystem responses to nitrogen deposition: A trait-based predictive framework. J. Ecol. 2018, 106, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, H.J.; George, E. Effect of plant nitrogen status on the contribution of arbuscular mycorrhizal hyphae to plant nitrogen uptake. Physiol. Plant 1999, 105, 694–700. [Google Scholar] [CrossRef]
- Polcyn, W.; Paluch-Lubawa, E.; Lehmann, T.; Mikuła, R. Arbuscular mycorrhiza in highly fertilized maize cultures alleviates short-term drought effects but does not improve fodder yield and quality. Front. Plant Sci. 2019, 10, 496. [Google Scholar] [CrossRef]
- Arumugam, A.; Malolan, V.V.; Ponnusami, V. Contemporary pretreatment strategies for bioethanol production from corncobs: A comprehensive review. Waste Biomass Valori. 2021, 12, 577–612. [Google Scholar] [CrossRef]
- Ray, D.K.; Mueller, N.D.; West, P.C.; Foley, J.A. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.; Hou, P.; Wu, L.; Chen, X.; Cui, Z.; Zhang, F. Understanding production potentials and yield gaps in intensive maize production in China. Field Crops Res. 2013, 143, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Schulthess, U.; Timsina, J.; Herrera, J.M.; McDonald, A. Mapping field-scale yield gaps for maize: An example from Bangladesh. Field Crops Res. 2013, 143, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Blake, D.J.; Thiengburanathum, P.; Thiengburanathum, P.; Friend, R.M.; Doherty, B.; Thankappan, S. Looking at complex agri-food systems from an actor perspective: The case of Northern Thailand. Adv. Food Secur. Sustain. 2019, 4, 33–65. [Google Scholar] [CrossRef]
- Yap, V.Y.; Xaphokhame, P.; de Neergaard, A.; Bech Bruun, T. Barriers to agro-ecological intensification of smallholder upland farming systems in Lao PDR. Agronomy 2019, 9, 375. [Google Scholar] [CrossRef] [Green Version]
- Shi, S.M.; Chen, K.; Gao, Y.; Liu, B.; Yang, X.H.; Huang, X.Z.; Liu, G.X.; Zhu, L.Q.; He, X.H. Arbuscular mycorrhizal fungus species dependency governs better plant physiological characteristics and leaf quality of mulberry (Morus alba L.) seedlings. Front. Microbiol. 2016, 7, 1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bücking, H.; Kafle, A. Role of arbuscular mycorrhizal fungi in the nitrogen uptake of plants: Current knowledge and research gaps. Agronomy 2015, 4, 587–612. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhong, H.; Zhu, L.; Yuan, Y.; Xu, L.; Wang, G.G.; Zhai, L.; Yang, L.; Zhang, J. Arbuscular mycorrhizal fungi effectively enhance the growth of Gleditsia sinensis Lam. seedlings under greenhouse conditions. Forests 2019, 10, 567. [Google Scholar] [CrossRef] [Green Version]
- Fellbaum, C.R.; Gachomo, E.W.; Beesetty, Y.; Choudhari, S.; Strahan, G.D.; Pfeffer, P.E.; Kiersd, E.T.; Bückinga, H. Carbon availability triggers fungal nitrogen uptake and transport in arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2012, 109, 2666–2671. [Google Scholar] [CrossRef] [Green Version]
- Mauromicale, G.; Ierna, A.; Marchese, M. Chlorophyll fluorescence and chlorophyll content in field-grown potato as affected by nitrogen supply, genotype, and plant age. Photosynthetica 2006, 44, 76–82. [Google Scholar] [CrossRef]
- Song, X.; Wang, Y.; Lv, X. Responses of plant biomass, photosynthesis and lipid peroxidation to warming and precipitation change in two dominant species (Stipa grandis and Leymus chinensis) from North China Grasslands. Ecol. Evol. 2016, 6, 1871–1882. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and artifact. Plant Physiol. 2019, 1801, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodge, A.; Fitter, A.H. Substantial nitrogen acquisition by arbuscular mycorrhizal fungi from organic material has implications for N cycling. Proc. Natl. Acad. Sci. USA 2010, 107, 13754–13759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Liao, W.; Liu, C.; Wen, Z.; Kincaid, R.L.; Harrison, J.H.; Elliott, D.C.; Brown, M.D.; Solana, A.E.; Stevens, D.J. Value-Added Chemicals from Animal Manure; Final Technical Report, Contract DE-AC06–76RL01830; US Department of Energy: Washington, DC, USA, 2003.
- Jin, H.R.; Liu, J.; Liu, J.; Huang, X. Forms of nitrogen uptake, translocation, and transfer via arbuscular mycorrhizal fungi: A review. Sci. China Life Sci. 2012, 55, 474–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bungau, S.; Behl, T.; Aleya, L.; Bourgeade, P.; Aloui-Sossé, B.; Purza, A.L.; Abid, A.; Samuel, A.D. Expatiating the impact of anthropogenic aspects and climatic factors on long-term soil monitoring and management. Environ. Sci. Pollut. Res. 2021, 28, 30528–30550. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.; Zhang, Q.; Pan, J.; Jiang, S.; Liu, Y.; Bahadur, A.; Peng, Z.; Yang, Y.; Feng, H. Effect of arbuscular mycorrhizal fungi on soil enzyme activity is coupled with increased plant biomass. Eur. J. Soil Sci. 2019, 71, 84–92. [Google Scholar] [CrossRef]
- Samuel, A.D.; Bungau, S.; Tit, D.M.; Melinte, C.E.; Purza, L.; Badea, G.E. Effects of long-term application of organic and mineral fertilizers on soil enzymes. Rev. Chem. 2018, 69, 2608–2612. [Google Scholar] [CrossRef]
- Samuel, A.D.; Tit, D.M.; Melinte, C.E.; Iovan, C.; Purza, L.; Gitea, M.; Bungau, S. Enzymological and physicochemical evaluation of the effects of soil management practices. Rev. Chim. 2017, 68, 2243–2247. [Google Scholar] [CrossRef]
- Fang, F.; Wang, C.; Wu, F.; Tang, M.; Doughty, R. Arbuscular mycorrhizal fungi mitigate nitrogen leaching under poplar seedlings. Forests 2020, 11, 325. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Fang, F.; Wu, N.; Li, L.; Tang, M. Nitrate transporter gene expression and kinetics of nitrate uptake by Populus canadensis ‘Neva’ in relation to arbuscular mycorrhizal fungi and nitrogen availability. Front. Microbiol. 2020, 11, 176. [Google Scholar] [CrossRef] [Green Version]
- Diouf, A.; Ndiaye, M.; Fall-Ndiaye, M.; Diop, T. Maize crop N uptake from organic material of Gliricidia sepium coinoculated with rhizobium and arbuscular mycorrhizal fungus in sub-Saharian africa sandy soil. Am. J. Plant Sci. 2017, 8, 428–440. [Google Scholar] [CrossRef] [Green Version]
- Püschel, D.; Janoušková, M.; Voríšková, A.; Gryndlerová, H.; Vosátka, M.; Jansa, J. Arbuscular mycorrhiza stimulates biological nitrogen fixation in two Medicago spp. through improved phosphorus acquisition. Front. Plant Sci. 2017, 8, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitakdantham, R.; Suwanarit, A.; Suwanarit, P.; Leyval, C.; Sarobol, E.; Nopamornbodi, O.; Hoefner, W. Effects of arbuscular mycorrhizal fungal species (Glomus aggregatum and Acaulospora spinosa), nitrogen and phosphorus fertilizers on maize growth, root colonization and spore production. Thai J. Agric. Sci. 2004, 37, 173–180. [Google Scholar]
- Karasawa, T.; Takebe, M.; Kasahara, Y. Arbuscular mycorrhizal (AM) effects on maize growth and AM colonization of roots under various soil moisture conditions. Soil Sci. Plant Nutr. 2000, 46, 61–67. [Google Scholar] [CrossRef]
- Punyalue, A.; Jongjaidee, J.; Jamjod, S.; Rerkasem, B. Legume intercropping to reduce erosion, increase soil fertility and grain yield, and stop burning in highland maize production in Northern Thailand. CMU J. Nat. Sci. 2018, 17, 265–274. [Google Scholar] [CrossRef]
- Lang, C.A. Simple micro determination of Kjeldahl nitrogen in biological materials. Anal. Chem. 1958, 30, 1692–1694. [Google Scholar] [CrossRef]
- Shabala, S.N.; Shabala, S.I.; Martynenko, A.I.; Babourina, O.; Newman, I.A. Salinity effect on bioelectric activity growth, Na+ accumulation and chlorophyll fluorescence of maize leaves: A comparative survey and prospects for screening. Aust. J. Plant Physiol. 1998, 25, 609–616. [Google Scholar] [CrossRef]
- Jägerbrand, A.K.; Kudo, G. Short-term responses in maximum quantum yield of PSII (Fv/Fm) to ex-situ temperature treatment of populations of bryophytes originating from different sites in Hokkaido, Northern Japan. Plants 2016, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Zhou, Y.; Lam, H.M.; Zhang, J. Inhibition of photosynthesis and energy dissipation induced by water and high light stresses in rice. J. Exp. Bot. 2007, 58, 1207–1217. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | TCh (mg g−1 FW) | Fv/Fm | Leaf Area (cm2) | Shoot FW (g plant−1) | Root FW (g plant−1) | Shoot WC (%) | Root WC (%) |
|---|---|---|---|---|---|---|---|
| Con | 11.52 ± 2.05 | 0.61 ± 0.05 | 37.57 ± 8.12b | 1.30 ± 0.39b | 1.40 ± 0.14ab | 43.73 ± 3.24b | 42.17 ± 4.02b |
| FM | 14.27 ± 2.25 | 0.65 ± 0.09 | 55.62 ± 18.18a | 2.08 ± 0.59a | 1.76 ± 0.30a | 62.42 ± 4.24a | 52.74 ± 11.31a |
| AF | 10.01 ± 0.82 | 0.68 ± 0.02 | 49.19 ± 11.71ab | 1.88 ± 0.28ab | 0.87 ± 0.09b | 45.76 ± 0.39b | 48.30 ± 12.22ab |
| CE | 13.01 ± 1.78 | 0.64 ± 0.09 | 45.98 ± 5.69ab | 1.59 ± 0.24ab | 1.37 ± 0.17ab | 44.25 ± 7.33b | 47.95 ± 3.36ab |
| ANOVA | ns | ns | * | * | ** | ** | * |
| Study Approach with/without, Involving N and PR | Priority Effective AMF | Fertilizer Consideration | Plant Host | Reference |
|---|---|---|---|---|
| N and PR | Funneliformis mosseae | Type comparing (Plant/animal manure organic and chemical fertilizer) | Maize (Zea mays L.) | This study |
| N and PR | Glomus etunicatum, Diversispora spurca | ns 1 | Walnuts (Juglans regia L.) | [16] |
| N and PR (Under drought) | Rhizophagus irregularis | ns (Chemical dose) | Maize (Zea mays L.) | [19] |
| N and PR | Funneliformis mosseae, Acaulospora scrobiculata | ns | Mulberry (Morus alba L.) | [26] |
| N without PR | Mix of Rhizophagus irregularis and Funneliformis mosseae | Type comparing (Plant residue and mineral fertilizer) | Durum wheat (Triticum durum Desf.) | [15] |
| N without PR | Rhizophagus irregularis | ns (Chemical dose) | Poplar Populus×canadensis) | [40] |
| N without PR | Rhizophagus irregularis | ns | Poplar (Populus×canadensis) | [41] |
| N without PR | Rhizophagus aggregatum (plus Rhizobium) | ns (Plant organic fertilizer test) | Maize (Zea mays L.) | [42] |
| N without PR | Rhizophagus irregularis | ns | Legumes (Medicago spp.) | [43] |
| Without N, PR (Dry matter, yield) | Glomus aggregatum | ns (Chemical dose) | Maize (Zea mays L.) | [44] |
| Without N, PR (Growth parameters, phosphorus uptake) | Glomus spp. | ns | Maize (Zea mays L.) | [45] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yooyongwech, S.; Threeprom, W.; Tisarum, R.; Samphumphuang, T.; Chungloo, D.; Cha-um, S. Matching of Nitrogen Enhancement and Photosynthetic Efficiency by Arbuscular Mycorrhiza in Maize (Zea mays L.) in Relation to Organic Fertilizer Type. Plants 2022, 11, 369. https://doi.org/10.3390/plants11030369
Yooyongwech S, Threeprom W, Tisarum R, Samphumphuang T, Chungloo D, Cha-um S. Matching of Nitrogen Enhancement and Photosynthetic Efficiency by Arbuscular Mycorrhiza in Maize (Zea mays L.) in Relation to Organic Fertilizer Type. Plants. 2022; 11(3):369. https://doi.org/10.3390/plants11030369
Chicago/Turabian StyleYooyongwech, Suravoot, Waraporn Threeprom, Rujira Tisarum, Thapanee Samphumphuang, Daonapa Chungloo, and Suriyan Cha-um. 2022. "Matching of Nitrogen Enhancement and Photosynthetic Efficiency by Arbuscular Mycorrhiza in Maize (Zea mays L.) in Relation to Organic Fertilizer Type" Plants 11, no. 3: 369. https://doi.org/10.3390/plants11030369