
Potato (Solanum tuberosum L.) Plant Shoot and Root Changes under Abiotic Stresses—Yield Response


Abstract
:1. Introduction
2. Results
2.1. Significance of Tested Parameters
2.2. Changes in the Morphological Plant Parameters
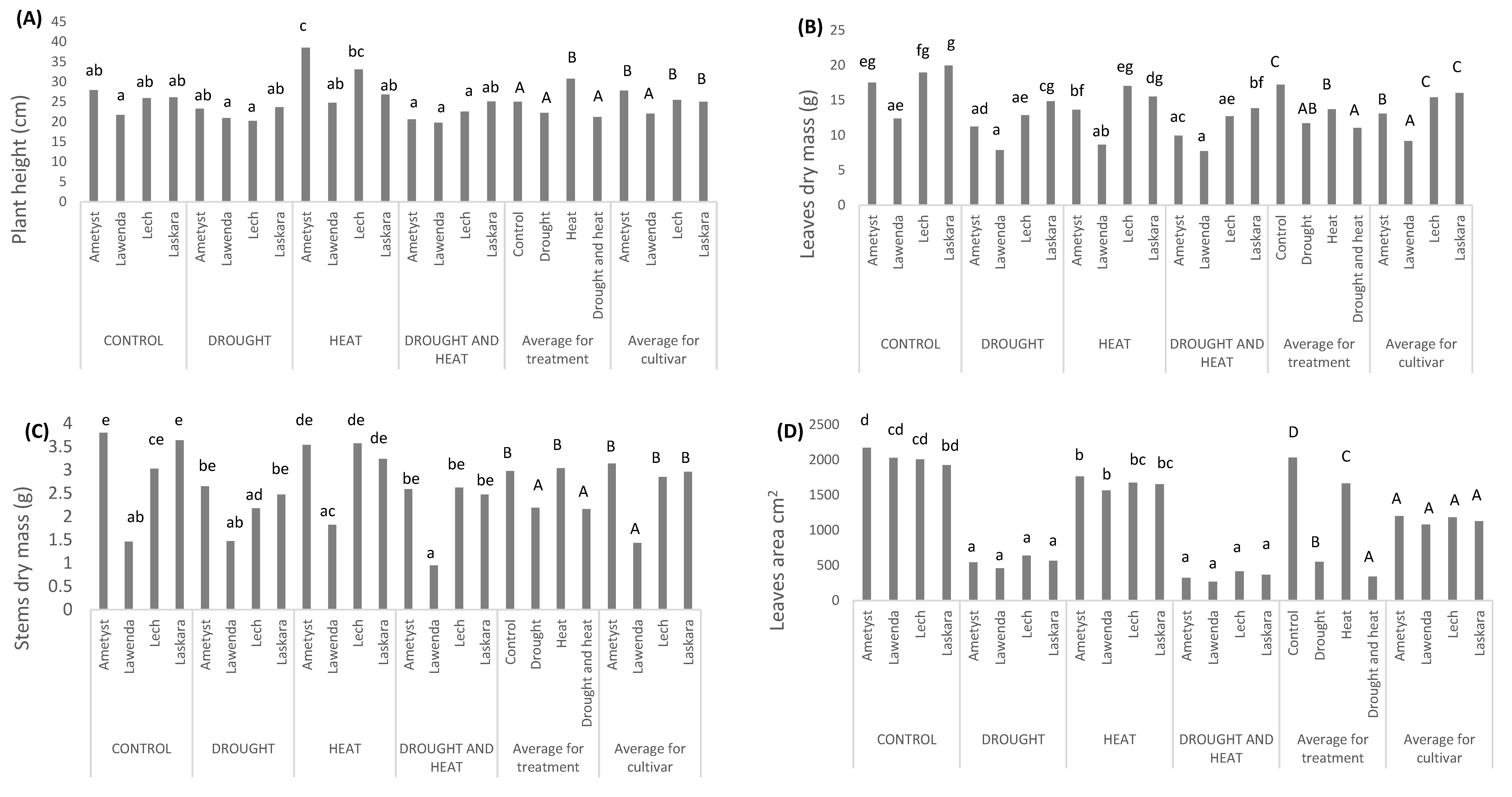
2.2.1. Plant Height Depending on the Applied Stress and Cultivar Used
2.2.2. Leaf Dry Mass Depending on the Applied Stress and Cultivar Used
2.2.3. Stem Dry Mass Depending on the Applied Stress and Cultivar Used
2.2.4. Leaf Area Depending on the Applied Stress and Cultivar Used
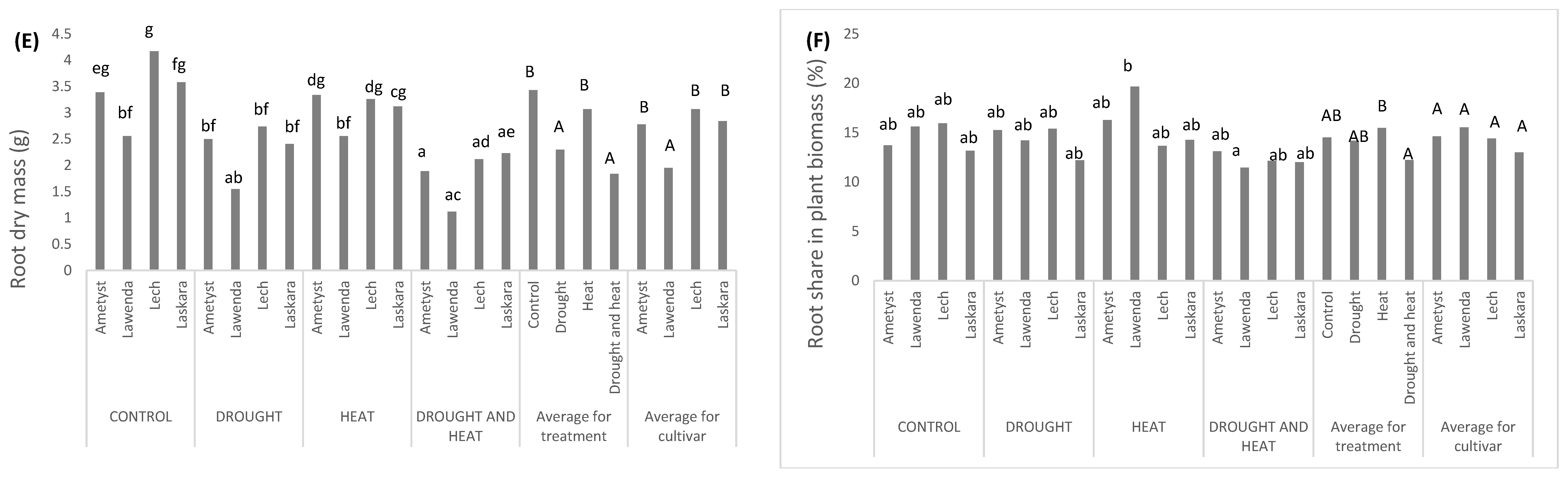
2.2.5. Root Dry Mass Depending on the Applied Stress and Cultivar Used
2.2.6. Root Share in Plant Biomass Depending on the Applied Stress and Cultivar Used
2.3. The Impact of Applied Stresses on the Tuber Yield
2.4. The Relationships between Selected Morphological Indicators of Plants and Yield Decrease under Stresses
3. Material and Methods
3.1. Plant Material
3.2. Tube Experiment
- control-optimal irrigation (70% of field water capacity) and optimal temperature (day/night temperature 22/18 °C);
- drought stress-remained without irrigation (40% of field water capacity) and optimal temperature (day/night 22/18 °C);
- high temperature stress-optimal irrigation (70% of field water capacity) and maintenance of elevated temperature (day/night temperature 38/25 °C);
- combined drought-high temperature stress-remained without irrigation (40% of field water capacity) and maintenance of elevated temperature (day/night temperature 38 °C/25 °C).
3.3. Pot Experiment concerning the Yield
4. Discussion
5. Main Conclusions
- Drought and high temperature, both separately and together, caused changes in plant morphology. Drought stress had a greater impact than high temperature stress. The biggest changes, however, took place when both stresses were applied simultaneously.
- Under all stresses, a decrease in tuber yield was found. The largest decrease was recorded in the case of both stresses applied simultaneously. The smallest decrease was visible in the case of high temperature stress.
- The greatest impact on the decrease in yield under the applied stresses was due to the root system size and its share in the entire biomass of the plant. This impact was higher in the case of drought stress than in heat stress.
- Our research confirms the importance of the root system size in the resistance of potato cultivars to abiotic stresses, and in particular to drought stress, and may be a valuable clue for potato growers.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Salinger, M.J.; Sivakumar, M.V.K.; Motha, R. Reducing vulnerability of agriculture and forestry to climate variability and change: Workshop summary and recommendations. Clim. Chang. 2005, 70, 341–362. [Google Scholar] [CrossRef]
- Cook, E.R.; Seager, R.; Cane, M.A.; Stahle, D.W. North American drought: Reconstructions, causes, and consequences. Earth-Sci. Rev. 2007, 81, 93–134. [Google Scholar] [CrossRef]
- Fischlin, A.; Midgley, G.F.; Price, J.T.; Leemans, R.; Gopal, B.; Turley, C.; Rounsevell, M.D.A.; Dude, O.P.; Tarazona, J.; Velichko, A.A. Ecosystems, their properties, goods, and services. In Climate Change: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Parry, M.L., Canziani, O.F., Palutikof, J.P., van der Linden, P.J., Hanson, C.E., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 211–272. [Google Scholar]
- Metz, B.; Davidson, O.; Bosch, P.; Dave, R.; Meyer, L. Mitigation of Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Holden, N.M.; Brereton, A.J.; Fealy, R.; Sweeney, J. Possible change in Irish climate and its impact on barley and potato yields. Agric. For. Meteorol. 2003, 116, 181–196. [Google Scholar] [CrossRef] [Green Version]
- George, T.S.; Taylor, M.A.; Dodd, I.C.; White, P.J. Climate change and consequences for potato production: A review of tolerance to emerging abiotic stress. Potato Res. 2018, 60, 239–268. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Verhagen, A. Climate change and its repercussions for the potato supply chain. Potato Res. 2008, 51, 223–237. [Google Scholar] [CrossRef]
- Hijmans, R.J. The effect of climate change on global potato production. Am. J. Potato Res. 2003, 80, 271–280. [Google Scholar] [CrossRef]
- Levy, D.; Veilleux, R.E. Adaptation of potato to high temperatures and salinity—A review. Am. J. Potato Res. 2007, 84, 487–506. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Jones, H.G.; Corlett, J.E. Current topics in drought physiology. J. Agric. Sci. 1992, 119, 291–296. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2011, 62, 869–882. [Google Scholar] [CrossRef]
- Grudzińska, M.; Boguszewska-Mańkowska, D.; Zarzyńska, K. Drought stress during the growing season: Changes in reducing sugars, starch content and respiration rate during storage of two potato cultivars differing in drought sensitivity. J. Agro. Crop Sci. 2022, 208, 609–620. [Google Scholar] [CrossRef]
- Evers, D.; Lefèvre, I.; Legay, S.; Lamoureux, D.; Hausman, J.F.; Rosales, R.O.G.; Marca, L.R.T.; Hoffmann, L.; Bonierbale, M.; Schafleitner, R. Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metabolite approach. J. Exp. Bot. 2010, 61, 2327–2343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mane, S.P.; Robinet, C.V.; Ulanov, A.; Schafleitner, R.; Tincopa, L.; Gaudin, A.; Nomberto, G.; Alvarado, C.; Solis, C.; Bolivar, L.A.; et al. Molecular and physiological adaptation to prolonged drought stress in the leaves of two Andean potato genotypes. Funct. Plant Biol. 2008, 35, 669–688. [Google Scholar] [CrossRef] [PubMed]
- Schafleitner, R.; Gutierrez, R.; Espino, R.; Gaudin, A.; Pérez, J.; Martínez, M.; Domínguez, A.; Tincopa, L.; Alvarado, C.; Numberto, G.; et al. Field screening for variation of drought tolerance in Solanum tuberosum L. by agronomical, physiological and genetic analysis. Potato Res. 2007, 50, 71–85. [Google Scholar] [CrossRef]
- Rykaczewska, K. The impact of high temperature during growing season on potato cultivars with different response to environmental stresses. Am. J. Plant Sci. 2013, 4, 2386–2393. [Google Scholar] [CrossRef] [Green Version]
- Lahlou, O.; Ouattar, S.; Ledent, J.F. The effect of drought and cultivar on growth parameters, yield, and yield components of potato. Agronomie 2003, 23, 257–268. [Google Scholar] [CrossRef]
- Handayani, T.; Watanabe, K. The combination of drought and heat stress has a greater effect on potato plants than single stresses. Plant Soil Environ. 2020, 66, 175–182. [Google Scholar] [CrossRef]
- Boguszewska-Mańkowska, D.; Zarzyńska, K.; Nosalewicz, A. Drought differentially affects root system size and architecture of potato cultivars with differing drought tolerance. Am. J. Pot. Res. 2020, 97, 54–62. [Google Scholar] [CrossRef]
- Nowacki, W. (Ed.) Characteristics of the National Register of Potato Cultivars. 2019. (In Polish). Available online: http://pw.ihar.edu.pl/blog/ht_project/charakterystyka-krajowego-rejestru-odmian-ziemniaka-wydanie-xxiii/ (accessed on 15 March 2019).
- Boguszewska-Mańkowska, D.; Pieczyński, M.; Wyrzykowska, A.; Kalaji, H.M.; Sieczko, L.; Szweykowska-Kulińska, Z.; Zagdańska, B. Divergent strategies displayed by potato (Solanum tuberosum L.) cultivars to cope with soil drought. J. Agron. Crop Sci. 2018, 204, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Monneveux, P.; Ramírez, D.A.; Pino, M.T. Drought tolerance in potato (S. tuberrosum L.): Can we learn from drought tolerance research in cereals? Plant Sci. 2013, 205, 76–86. [Google Scholar] [CrossRef]
- Iwama, K. Physiology of the potato: New insights into root system and repercussions for crop management. Potato Res. 2008, 51, 333–353. [Google Scholar] [CrossRef]
- Tuberosa, R. Phenotyping for drought tolerance of crops in the genomics era. Front. Physiol. 2012, 3, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anithakumari, A.M.; Nataraja, K.N.; Visser, R.G.F.; Van Der Linden, C.G. Genetic dissection of drought tolerance and recovery potential by quantitative trait locus mapping of a diploid potato population. Mol. Breed. 2012, 30, 1413–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albiski, F.; Najla, S.; Sanoubar, R.; Alkabani, N.; Murshed, R. In vitro screening of potato lines for drought tolerance. Physiol. Mol. Biol. Plants 2012, 18, 315–321. [Google Scholar] [CrossRef] [Green Version]
- Deblonde, P.M.K.; Ledent, J.F. Effects of moderate drought conditions on green leaf number, stem height, leaf length and tuber yield of potato cultivars. Eur. J. Agron. 2001, 14, 31–41. [Google Scholar] [CrossRef]
- Nasir, M.W.; Toth, Z. Response of Different Potato Genotypes to Drought Stress. Agriculture 2021, 11, 763. [Google Scholar] [CrossRef]
- Shah, N.H.; Paulsen, G.M. Interaction of drought and high temperature on photosynthesis and grain-filling of wheat. Plant Soil 2003, 257, 219–226. [Google Scholar] [CrossRef]
- Trenberth, K.E. The impact of climate change and variability on heavy precipitation, floods, and droughts. In Encyclopedia of Hydrological Sciences Hoboken; Anderson, M.G.M., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2008. [Google Scholar] [CrossRef] [Green Version]
- Andjelkovic, V. Introductory chapter: Climate changes and abiotic stress in plants. In Plant, Abiotic Stress, and Responses to Climate Change; Andjelkovic, V., Ed.; IntechOpen: Polje, Serbia, 2018; pp. 3–6. [Google Scholar] [CrossRef] [Green Version]
- Rykaczewska, K. The effect of high temperature occurring in subsequent stages of plant development on potato yield and tuber physiological defects. Am. J. Potato Res. 2015, 92, 339–349. [Google Scholar] [CrossRef]
- Wishart, J.; George, T.S.; Brown, L.K.; Ramsay, G.; Bradshaw, J.E.; White, P.J.; Gregory, P.J. Measuring variation in potato roots in both field and glasshouse: The search for useful yield predictors and a simple screen for root traits. Plant Soil 2013, 368, 231–249. [Google Scholar] [CrossRef]
- Wishart, J.; George, T.S.; Brown, L.K.; White, P.J.; Ramsay, G.; Jones, H.; Gregory, P.J. Field phenotyping of potato to assess root and shoot characteristics associated with drought tolerance. Plant Soil 2014, 378, 351–363. [Google Scholar] [CrossRef] [Green Version]
- Puértolas, J.; Ballester, C.; Elphinstone, E.D.; Dodd, I.C. Two potato (Solanum tuberosum) varieties differ in drought tolerance due to differences in root growth at depth. Funct. Plant Biol. 2014, 41, 1107–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bengough, A.G.; McKenzie, B.M.; Hallett, P.D.; Valentine, T.A. Root elongation, water stress, and mechanical impedance: A review of limiting stresses and beneficial root tip traits. J. Exp. Bot. 2011, 62, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahlou, O.; Ledent, J.F. Root mass and depth, stolons and roots formed on stolons in four cultivars of potato under water stress. Europ. J. Agron. 2005, 22, 159–173. [Google Scholar] [CrossRef]
- Zarzyńska, K.; Boguszewska-Mańkowska, D.; Nosalewicz, A. Differences in size and architecture of the potato cultivars root system and their tolerance to drought stress. Plant Soil Environ. 2017, 63, 159–164. [Google Scholar] [CrossRef] [Green Version]
- Jefferies, R.A. Cultivar responses to water stress in potato: Effects of shoot and roots. New Phytol. 1993, 123, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Vadez, V. Root hydraulics: The forgotten side of roots in drought adaptation. Field Crops Res. 2014, 165, 15–24. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Tested Stress | Sources of Variation | Plant Height (cm) | Leaves Dry Mass (g) | Stems Dry Mass (g) | Leaves Area (cm2) | Root Dry Mass (g) | Root Share in Plant Biomass (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| p-Value | Significance | p-Value | Significance | p-Value | Significance | p-Value | Significance | p-Value | Significance | p-Value | Significance | ||
| Drought | Cultivar | 0.4991 | - | 0.0000 | *** | 0.0000 | *** | 0.548 | - | 0.0091 | * | 0.3953 | - |
| Treatment | 0.1292 | - | 0.0000 | *** | 0.0000 | *** | 0.0000 | *** | 0.0000 | *** | 0.2521 | - | |
| Treatment × cultivar | 0.5820 | - | 0.784 | - | 0.0981 | - | 0.3681 | - | 0.7152 | - | 0.9790 | - | |
| Heat | Cultivar | 0.0026 | * | 0.000 | *** | 0.000 | *** | 0.694 | - | 0.0356 | * | 0.3791 | - |
| Treatment | 0.0033 | * | 0.000 | *** | 0.776 | - | 0.000 | *** | 0.0491 | * | 0.6412 | - | |
| Treatment × cultivar | 0.3701 | - | 0.802 | - | 0.328 | - | 0.883 | - | 0.6344 | - | 0.4412 | - | |
| Drought & Heat | Cultivar | 0.3231 | - | 0.0000 | *** | 0.0000 | *** | 0.6502 | - | 0.0085 | * | 0.5102 | - |
| Treatment | 0.1428 | - | 0.0000 | *** | 0.0003 | ** | 0.0000 | *** | 0.0000 | *** | 0.0466 | * | |
| Treatment × cultivar | 0.5971 | - | 0.6491 | - | 0.2502 | - | 0.3751 | - | 0.8293 | - | 0.7702 | - | |
| Drought Heat, Drought & Heat | Cultivar | 0.0000 | *** | 0.0000 | *** | 0.0000 | *** | 0.148 | - | 0.0000 | *** | 0.3471 | - |
| Treatment (Control, Drought, Heat, Drought & Heat) | 0.0010 | ** | 0.0000 | *** | 0.0000 | *** | 0.000 | *** | 0.0000 | *** | 0.0071 | * | |
| Treatment × cultivar | 0.0120 | * | 0.7721 | - | 0.0391 | * | 0.678 | - | 0.7271 | - | 0.1670 | - | |
| Cultivar/Treatment | Control | Drought | Decrease (Relative to Control) | Heat | Decrease (Relative to Control) | Drought and Heat | Decrease (Relative to Control) |
|---|---|---|---|---|---|---|---|
| Ametyst | 1467 c | 1184 a,b | 19.3 | 1263 b | 13.9 | 1120 b | 23.6 |
| Laskara | 1397 b,c | 1033 a,b | 26.0 | 1128 b | 19.2 | 955 a | 31.6 |
| Lawenda | 1433 c | 1027 a,b | 28.3 | 1184 b | 17.3 | 959 a | 33.1 |
| Lech | 1465 c | 1184 a,b | 19.2 | 1330 b,c | 9.2 | 1045 a,b | 28.6 |
| Mean | 1440 C | 1107 A,B | 23.2 | 1226 B | 14.9 | 1020 A | 29.2 |
| Tested Correlation | p | r | r2 |
|---|---|---|---|
| Yield decrease-plant height | 0.0393 | 0.0144 | 0.0002 |
| Yield decrease-leaves dry mass | 0.3190 | 0.2122 | 0.0450 |
| Yield decrease-stems dry mass | 0.7869 | −0.00582 | 0.0034 |
| Yield decrease-leaves area | 0.2090 | 0.2656 | 0.0430 |
| Yield decrease-root dry mass | 0.0006 | −0.6478 | 0.4190 |
| Yield decrease-root share in plant biomass | 0.0012 | −0.620 | 0.3850 |
| Tested Correlation | p | r | r2 |
|---|---|---|---|
| Yield decrease-plant height | 0.9466 | −0.0656 | 0.0043 |
| Yield decrease-leaves dry mass | 0.9750 | −0.0068 | 0.0000 |
| Yield decrease-stems mass | 0.4882 | 0.1487 | 0.0221 |
| Yield decrease-leaves area | 0.4450 | 0.1636 | 0.0268 |
| Yield decrease-root dry mass | 0.0734 | −0.0732 | 0.00054 |
| Yield decrease-root share in plant biomass | 0.6316 | −0.1031 | 0.0136 |
| Tested Correlation | p | r | r2 |
|---|---|---|---|
| Yield decrease-plant height | 0.0266 | 0.4521 | 0.2044 |
| Yield decrease-leaves dry mass | 0.1409 | 0.3096 | 0.0959 |
| Yield decrease-stem dry mass | 0.1415 | 0.3092 | 0.0956 |
| Yield decrease-leaves area | 0.0926 | 0.3511 | 0.1232 |
| Yield decrease-root dry mass | 0.0009 | −0.6350 | 0.4032 |
| Yield decrease-root share in plant biomass | 0.0001 | −0.6989 | 0.4885 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mańkowska, D.; Zarzyńska, K.; Wasilewska-Nascimento, B. Potato (Solanum tuberosum L.) Plant Shoot and Root Changes under Abiotic Stresses—Yield Response. Plants 2022, 11, 3568. https://doi.org/10.3390/plants11243568
Mańkowska D, Zarzyńska K, Wasilewska-Nascimento B. Potato (Solanum tuberosum L.) Plant Shoot and Root Changes under Abiotic Stresses—Yield Response. Plants. 2022; 11(24):3568. https://doi.org/10.3390/plants11243568
Chicago/Turabian StyleMańkowska, Dominika, Krystyna Zarzyńska, and Beata Wasilewska-Nascimento. 2022. "Potato (Solanum tuberosum L.) Plant Shoot and Root Changes under Abiotic Stresses—Yield Response" Plants 11, no. 24: 3568. https://doi.org/10.3390/plants11243568