
Rich in Phenolics—Strong Antioxidant Fruit? Comparative Study of 25 Strawberry Cultivars
 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Phytochemical Composition
2.2. Antiradical Activity
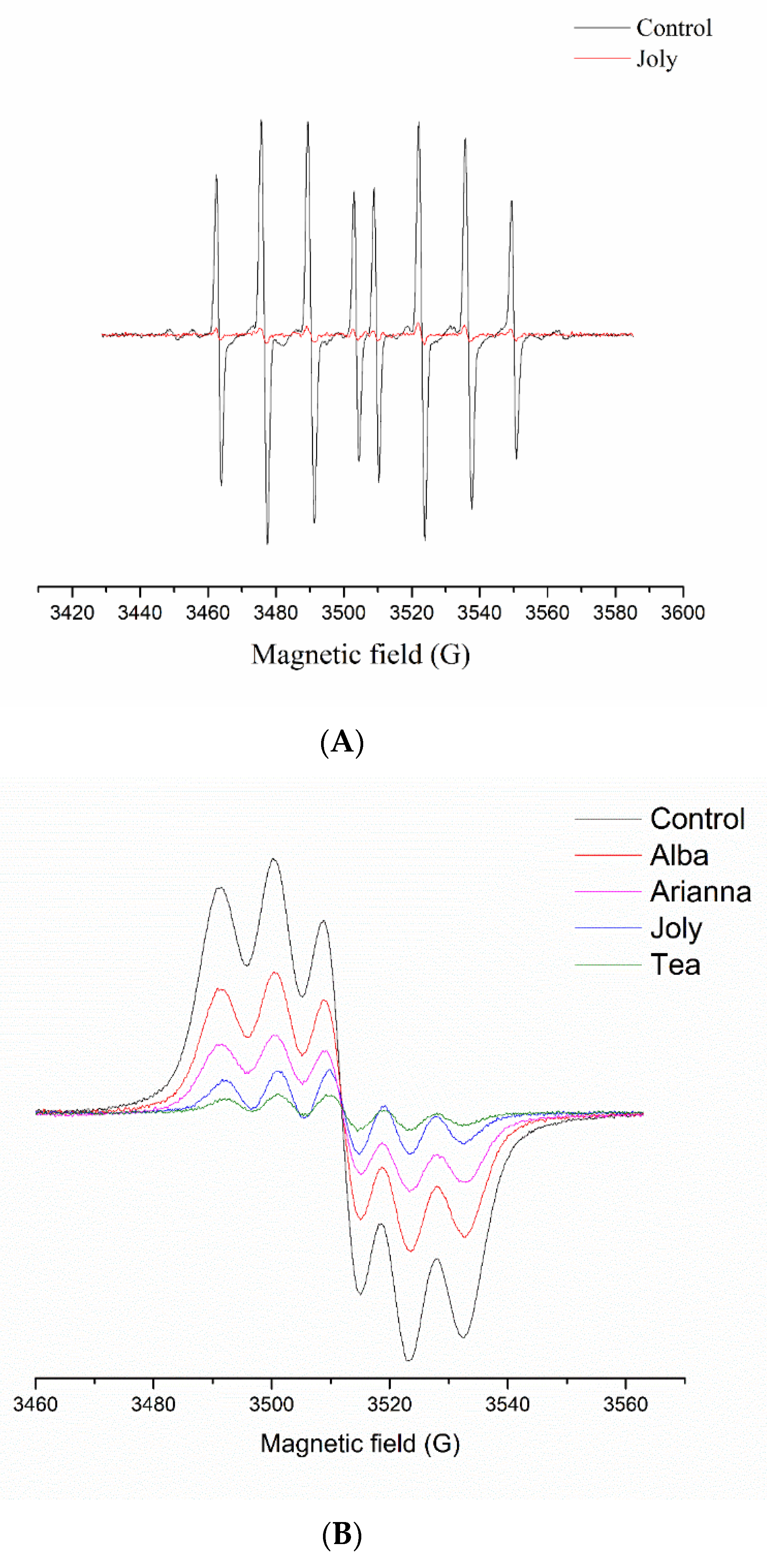
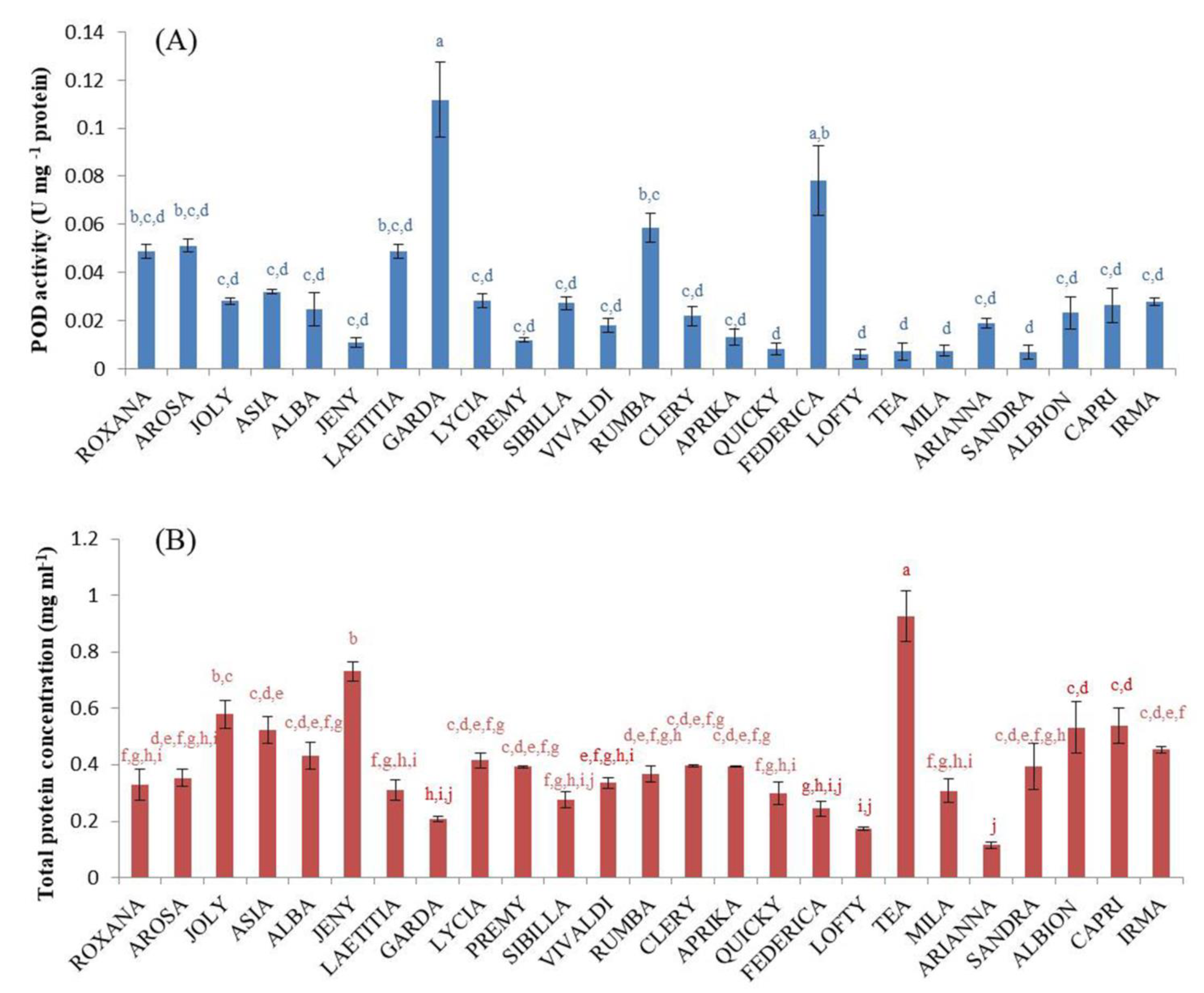
2.3. Peroxidase (POD) Activity
2.4. Polyphenols Profiles
3. Materials and Methods
3.1. Plant Materials
3.2. Sample Preparation
3.3. Spectrophotometric Measurements
3.4. Vitamin C
3.5. HPLC Analysis
3.6. Electron Paramagnetic Resonance (EPR) Spectroscopy
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cosme, F.; Pinto, T.; Aires, A.; Morais, M.C.; Bacelar, E.; Anjos, R.; Ferreira-Cardoso, J.; Oliveira, I.; Vilela, A.; Gonçalves, B. Red Fruits Composition and Their Health Benefits—A Review. Foods 2022, 11, 644. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Alvarez-Suarez, J.M.; Mazzoni, L.; Romandini, S.; Bompadre, S.; Diamanti, J.; Capocasa, F.; Mezzetti, B.; Quiles, J.L.; Ferreiro, M.S.; et al. The potential impact of strawberry on human health. Nat. Prod. Res. 2013, 27, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Aaby, K.; Mazur, S.; Nes, A.; Skrede, G. Phenolic compounds in strawberry (Fragaria x ananassa Duch.) fruits: Composition in 27 cultivars and changes during ripening. Food Chem. 2012, 132, 86–97. [Google Scholar] [CrossRef]
- Heinonenm, M.I.; Meyer, A.S.; Frankel, E.N. Antioxidant activity of berry phenolics on human low-density lipoprotein and liposome oxidation. J. Agric. Food Chem. 1998, 46, 4107–4112. [Google Scholar] [CrossRef]
- Beattie, J.; Crozier, A.; Duthie, G. Potential Health Benefits of Berries. Curr. Nutr. Food Sci. 2005, 1, 71–86. [Google Scholar] [CrossRef]
- Häkkinen, S.H.; Kärenlampi, S.O.; Heinonen, I.M.; Mykkänen, H.M.; Törrönen, A.R. Content of the flavonols quercetin, myricetin, and kaempferol in 25 edible berries. J. Agric. Food Chem. 1999, 47, 2274–2279. [Google Scholar] [PubMed]
- Di Vittori, L.; Mazzoni, L.; Battino, M.; Mezzetti, B. Pre-harvest factors influencing the quality of berries. Sci. Hortic. 2018, 233, 310–322. [Google Scholar] [CrossRef]
- Tomić, J.; Pešaković, M.; Karaklajić-Stajić, Ž.; Paunović, S.M.; Milinković, M.; Rilak, B. Quality Assessment of Strawberry Cultivars Grown in Western Serbia; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2021; pp. 955–962. [Google Scholar]
- Krüger, E.; Josuttis, M.; Nestby, R.; Toldam-Andersen, T.B.; Carlen, C.; Mezzetti, B. Influence of growing conditions at different latitudes of Europe on strawberry growth performance, yield and quality. J. Berry Res. 2012, 2, 143–157. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Cao, G.; Prior, R.L. Total antioxidant capacity of fruits. J. Agric. Food Chem. 1996, 44, 701–705. [Google Scholar] [CrossRef]
- Guo, C.; Yang, J.; Wei, J.; Li, Y.; Xu, J.; Jiang, Y. Antioxidant activities of peel, pulp and seed fractions of common fruits as determined by FRAP assay. Nutr. Res. 2003, 23, 1719–1726. [Google Scholar] [CrossRef]
- Sun, J.; Chu, Y.-F.; Wu, X.; Liu, R.H. Antioxidant and antiproliferative activities of common fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar] [CrossRef] [PubMed]
- Meyers, K.J.; Watkins, C.B.; Pritts, M.P.; Liu, R.H. Antioxidant and antiproliferative activities of strawberries. J. Agric. Food Chem. 2003, 51, 6887–6892. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Cao, G.; Prior, R.L. Oxygen radical absorbing capacity of anthocyanins. J. Agric. Food Chem. 1997, 45, 304–309. [Google Scholar] [CrossRef]
- Mazza, G.; Miniati, E. Anthocyanins in Fruits, Vegetables, and Grains; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Hong, V.; Wrolstad, R.E. Use of HPLC separation/photodiode array detection for characterization of anthocyanins. J. Agric. Food Chem. 1990, 38, 708–715. [Google Scholar] [CrossRef]
- Milosavljević, D.M.; Mutavdžić, D.R.; Radotić, K.; Milivojević, J.M.; Maksimović, V.M.; Dragišić Maksimović, J. Phenolic profiling of 12 strawberry cultivars using different spectroscopic methods. J. Agric. Food Chem. 2020, 68, 4346–4354. [Google Scholar] [CrossRef]
- Shirley, B.W. Flavonoid biosynthesis: ‘New’ functions for an ‘old’ pathway. Trends Plant Sci. 1996, 1, 377–382. [Google Scholar]
- Nile, S.H.; Park, S.W. Edible berries: Bioactive components and their effect on human health. Nutrition 2014, 30, 134–144. [Google Scholar] [CrossRef]
- Fernandes, V.C.; Domingues, V.F.; de Freitas, V.; Delerue-Matos, C.; Mateus, N. Strawberries from integrated pest management and organic farming: Phenolic composition and antioxidant properties. Food Chem. 2012, 134, 1926–1931. [Google Scholar] [CrossRef] [Green Version]
- Bors, W.; Saran, M. Radical scavenging by flavonoid antioxidants. Free. Radic. Res. Commun. 1987, 2, 289–294. [Google Scholar] [CrossRef]
- Das, D.K. [40] Naturally occurring flavonoids: Structure, chemistry, and high-performance liquid chromatography methods for separa-tion and characterization. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1994; pp. 410–420. [Google Scholar]
- Romeyer, F.M.; Macheix, J.J.; Goiffon, J.J.; Reminiac, C.C.; Sapis, J.C. Browning capacity of grapes. 3. Changes and importance of hydroxycinnamic acid-tartaric acid esters during development and maturation of the fruit. J. Agric. Food Chem. 1983, 31, 346–349. [Google Scholar] [CrossRef]
- Chisari, M.; Barbagallo, R.N.; Spagna, G. Characterization of polyphenol oxidase and peroxidase and influence on browning of cold stored strawberry fruit. J. Agric. Food Chem. 2007, 55, 3469–3476. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-X.; Schopfer, P. Hydroxyl-radical production in physiological reactions. A novel function of peroxidase. Eur. J. Biochem. 1999, 260, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Wesche-Ebeling, P.; Montgomery, M.W. Strawberry Polyphenoloxidase: Purification and Characterization. J. Food Sci. 1990, 55, 1315–1319. [Google Scholar] [CrossRef]
- Nowicka, A.; Kucharska, A.Z.; Sokół-Łętowska, A.; Fecka, I. Comparison of polyphenol content and antioxidant capacity of strawberry fruit from 90 cultivars of Fragaria× ananassa Duch. Food Chem. 2019, 270, 32–46. [Google Scholar] [CrossRef]
- Milosavljević, D.; Maksimović, V.; Milivojević, J.; Dragišić Maksimović, J. A Comparison of Major Taste- and Health-Related Compounds among Newly Released Italian Strawberry Cultivars; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2021; pp. 841–848. [Google Scholar]
- Urün, I.; Attar, S.H.; Sönmez, D.A.; Gündeşli, M.A.; Ercişli, S.; Kafkas, N.E.; Bandić, L.M.; Duralija, B. Comparison of polyphenol, sugar, organic acid, volatile compounds, and antioxidant capacity of commercially grown strawberry cultivars in Turkey. Plants 2021, 10, 1654. [Google Scholar] [CrossRef]
- Häkkinen, S.H.; Törrönen, A.R. Content of flavonols and selected phenolic acids in strawberries and Vaccinium species: Influence of cultivar, cultivation site and technique. Food Res. Int. 2000, 33, 517–524. [Google Scholar] [CrossRef]
- Mockevičiūtė, R.; Jurkonienė, S.; Gavelienė, V.; Jankovska-Bortkevič, E.; Šocik, B.; Armalytė, G.; Budrys, R. Effects Induced by the Agricultural Application of Probiotics on Antioxidant Potential of Strawberries. Plants 2022, 11, 831. [Google Scholar] [CrossRef]
- Klopotek, Y.; Otto, K.; Böhm, V. Processing Strawberries to Different Products Alters Contents of Vitamin C, Total Phenolics, Total Anthocyanins, and Antioxidant Capacity. J. Agric. Food Chem. 2005, 53, 5640–5646. [Google Scholar] [CrossRef]
- Pelayo, C.; Ebeler, S.; Kader, A. Postharvest life and flavor quality of three strawberry cultivars kept at 5°C in air or air+20 kPa CO2. Postharvest Biol. Technol. 2003, 27, 171–183. [Google Scholar] [CrossRef]
- Dragišić Maksimović, J.; Poledica, M.; Mutavdžić, D.; Mojović, M.; Radivojević, D.; Milivojević, J. Variation in nutritional quality and chemical composition of fresh strawberry fruit: Combined effect of cultivar and storage. Plant Foods Hum. Nutr. 2015, 70, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Kafkas, E.; Koşar, M.; Paydaş, S.; Kafkas, S.; Başer, K. Quality characteristics of strawberry genotypes at different maturation stages. Food Chem. 2007, 100, 1229–1236. [Google Scholar] [CrossRef]
- Milivojević, J.; Rakonjac, V.; Akšić, M.F.; Pristov, J.B.; Maksimović, V. Classification and fingerprinting of different berries based on biochemical profiling and antioxidant capacity. Pesqui. Agropecuária Bras. 2013, 48, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Walz, C.; Juenger, M.; Schad, M.; Kehr, J. Evidence for the presence and activity of a complete antioxidant defence system in mature sieve tubes. Plant J. 2002, 31, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Rice-Evans, A.C.; Diplock, A.T.; Symons, M.R. Techniques in free radical research. Lab. Tech. Biochem. Mol. Biol. 1991, 22, 1–278. [Google Scholar]
- Scandalios, J. Oxidative stress and defense mechanisms in plants: Introduction. Free. Radic. Biol. Med. 1997, 23, 471–472. [Google Scholar] [CrossRef]
- Osorio, C.; Carriazo, J.G.; Almanza, O. Antioxidant activity of corozo (Bactris guineensis) fruit by electron paramagnetic resonance (EPR) spectroscopy. Eur. Food Res. Technol. 2011, 233, 103–108. [Google Scholar] [CrossRef]
- Kostka, T.; Ostberg-Potthoff, J.J.; Briviba, K.; Matsugo, S.; Winterhalter, P.; Esatbeyoglu, T. Pomegranate (Punica granatum L.) extract and its anthocyanin and copigment fractions—Free radical scavenging activity and influence on cellular oxidative stress. Foods 2020, 9, 1617. [Google Scholar] [CrossRef] [PubMed]
- Brezová, V.; Šlebodová, A.; Staško, A. Coffee as a source of antioxidants: An EPR study. Food Chem. 2009, 114, 859–868. [Google Scholar] [CrossRef]
- Savić, A.G.; Mojović, M. Free radicals identification from the complex EPR signals by applying higher order statistics. Anal. Chem. 2012, 84, 3398–3402. [Google Scholar] [CrossRef]
- Bačíc, G.; Mojović, M. EPR spin trapping of oxygen radicals in plants: A methodological overview. Ann. New York Acad. Sci. 2005, 1048, 230–243. [Google Scholar] [CrossRef]
- Alvarez-Suarez, J.M.; Dekanski, D.; Ristić, S.; Radonjić, N.V.; Petronijević, N.D.; Giampieri, F.; Astolfi, P.; González-Paramás, A.M.; Santos-Buelga, C.; Tulipani, S.; et al. Strawberry polyphenols attenuate ethanol-induced gastric lesions in rats by activation of antioxidant enzymes and attenuation of MDA increase. PLoS ONE 2011, 6, e25878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moazzen, A.; Öztinen, N.; Ak-Sakalli, E.; Koşar, M. Structure-antiradical activity relationships of 25 natural antioxidant phenolic compounds from different classes. Heliyon 2022, 8, e10467. [Google Scholar] [CrossRef] [PubMed]
- López-Serrano, M.; Barceló, A. Peroxidase in unripe and processing-ripe strawberries. Food Chem. 1995, 52, 157–160. [Google Scholar] [CrossRef]
- López-Serrano, M.; Barceló, A.R. Histochemical localization and developmental expression of peroxidase and polyphenol oxidase in strawberries. J. Am. Soc. Hortic. Sci. 2001, 126, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Civello, P.M.; Martinez, G.A.; Chaves, A.R.; Anon, M.C. Peroxidase from strawberry fruit (Fragaria × ananassa Duch.): Partial purification and determination of some properties. J. Agric. Food Chem. 1995, 43, 2596–2601. [Google Scholar] [CrossRef]
- Kozłowska, M.; Fryder, K.; Wolko, B. Peroxidase involvement in the defense response of red raspberry to Didymella applanate (Niessl/Sacc.). Acta Physiol. Plant. 2001, 23, 303–310. [Google Scholar] [CrossRef]
- Menon, S.V.; Rao, T.R. Enzyme activities during the development and ripening of watermelon (Citrullus lanatus (Thunb.) Matsum. & Nakai) fruit. Int. J. Plant Dev. Biol. 2012, 6, 21–26. [Google Scholar]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vennat, B.; Bos, M.; Pourrat, A.; Bastide, P. Procyanidins from tormentil: Fractionation and study of the anti-radical activity towards superoxide anion. Biol. Pharm. Bull. 1994, 17, 1613–1615. [Google Scholar] [CrossRef] [PubMed]
- Crecente-Campo, J.; Nunes-Damaceno, M.; Romero-Rodríguez, M.; Vázquez-Odériz, M. Color, anthocyanin pigment, ascorbic acid and total phenolic compound determination in organic versus conventional strawberries (Fragaria ananassa Duch, cv Selva). J. Food Compos. Anal. 2012, 28, 23–30. [Google Scholar] [CrossRef]
- Yoshida, Y.; Tamura, H. Variation in Concentration and Composition of Anthocyanins among Strawberry Cultivars. J. Jpn. Soc. Hortic. Sci. 2005, 74, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Dragišić Maksimović, J.; Mojović, M.; Vučinić, Ž.; Maksimović, V. Spatial distribution of apoplastic antioxidative constituents in maize root. Physiol. Plant. 2021, 173, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Puhl, I.; Treutter, D. Ontogenetic variation of catechin biosynthesis as basis for infection and quiescence of Botrytis cinerea in developing strawberry fruits. J. Plant Dis. Prot. 2008, 115, 247–251. [Google Scholar] [CrossRef]
- Arts, I.C.W.; Van De Putte, B.; Hollman, P.C.H. Catechin Contents of Foods Commonly Consumed in The Netherlands. 1. Fruits, Vegetables, Staple Foods, and Processed Foods. J. Agric. Food Chem. 2000, 48, 1746–1751. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Hossain, H.; Hossain, M.; Reza, H.M.; Rahman, M.M.; Ullah, M.O. Hydroxycinnamic acid derivatives: A potential class of natural compounds for the management of lipid metabolism and obesity. Nutr. Metab. 2016, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- Kosar, M.; Kafkas, E.; Paydas, S.; Baser, K.H.C. Phenolic composition of strawberry genotypes at different maturation stages. J. Agric. Food Chem. 2004, 52, 1586–1589. [Google Scholar] [CrossRef]
- Williner, M.R.; Pirovani, M.E.; Güemes, D.R. Ellagic acid content in strawberries of different cultivars and ripening stages. J. Sci. Food Agric. 2003, 83, 842–845. [Google Scholar] [CrossRef]
- Muthukumaran, S.; Tranchant, C.; Shi, J.; Ye, X.; Xue, S.J. Ellagic acid in strawberry (Fragaria spp.): Biological, technological, stability, and human health aspects. Food Qual. Saf. 2017, 1, 227–252. [Google Scholar] [CrossRef] [Green Version]
- Tomić, J.; Pešaković, M.; Milivojević, J.; Karaklajić-Stajić, Ž. How to improve strawberry productivity, nutrients composition, and beneficial rhizosphere microflora by biofertilization and mineral fertilization? J. Plant Nutr. 2018, 41, 2009–2021. [Google Scholar] [CrossRef]
- Dragišić Maksimović, J.J.; Živanović, B.D. Quantification of the antioxidant activity in salt-stressed tissues. In Plant Salt Tolerance; Springer: Berlin/Heidelberg, Germany, 2012; pp. 237–250. [Google Scholar]
- Cheng, G.W.; Breen, P.J. Activity of phenylalanine ammonia-lyase (PAL) and concentrations of anthocyanins and phenolics in developing strawberry fruit. J. Am. Soc. Hortic. Sci. 1991, 116, 865–869. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hammerschmidt, R.; Nuckles, E.M.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Pantelidis, G.E.; Vasilakakis, M.; Manganaris, G.A.; Diamantidis, G. Antioxidant capacity, phenol, anthocyanin and ascorbic acid contents in raspberries, blackberries, red currants, gooseberries and Cornelian cherries. Food Chem. 2007, 102, 777–783. [Google Scholar] [CrossRef]
- Nakarada, Đ.; Pejin, B.; Tommonaro, G.; Mojović, M. Liposomal integration method for assessing antioxidative activity of water insoluble compounds towards biologically relevant free radicals: Example of avarol. J. Liposome Res. 2020, 30, 218–226. [Google Scholar] [CrossRef]
- Nakarada, Đ.J.; Marković, S.Z.; Popović, M.D.; Dimitrijević, M.S.; Rakić, A.A.; Mojović, M. Redox properties of grape wine skin extracts from the Šumadija region: An electron paramagnetic resonance study. Hosp. Pharmacol. Int. Multidiscip. J. 2021, 8, 1004–1013. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cultivars | TPC (mg eq GA g−1 FW) | TACY (mg eq Pg-3-G 100 g−1 FW) | Vit C (mg 100 g−1 FW) |
|---|---|---|---|
| Roxana | 0.49 ± 0.04 g,h | 15.89 ± 0.23 g,h | 52.80 ± 0.00 b |
| Arosa | 0.71 ± 0.02 e,f,g,h | 11.05 ± 0.16 h,i,j,k | 55.17 ± 0.98 a,b |
| Joly | 0.88 ± 0.09 b,c,d,e | 22.14 ± 0.68 b,c,d,e,f | 42.83 ± 1.27 d |
| Asia | 0.66 ± 0.00 d,e,f,g,h | 15.70 ± 1.62 g,h | 42.83 ± 1.27 d |
| Alba | 0.49 ± 0.03 h | 7.90 ± 1.93 j,k,l | 42.80 ± 1.04 d |
| Jeny | 0.50 ± 0.12 f,g,h | 3.08 ± 0.41 l | 39.82 ± 0.42 d |
| Laetitia | 1.07 ± 0.03 b,c | 15.83 ± 1.22 f,g,h | 56.32 ± 1.87 a |
| Garda | 0.54 ± 0.03 f,g,h | 6.79 ± 0.36 i,j,k,l | 54.57 ± 1.53 a,b |
| Lycia | 1.28 ± 0.08 a,b | 8.23 ± 0.16 i,j,k,l | 42.20 ± 0.85 d |
| Premy | 1.53 ± 0.25 a | 12.08 ± 2.48 h,i | 34.17 ± 1.65 e,f |
| Sibilla | 1.33 ± 0.15 a | 8.66 ± 0.64 i,j,k | 32.27 ± 0.49 f |
| Vivaldi | 1.37 ± 0.03 a | 24.92 ± 1.91 a,b,c | 46.70 ± 0.92 c |
| Rumba | 1.40 ± 0.10 a | 19.46 ± 3.25 d,e,f,g | 49.07 ± 1.20 c |
| Clery | 0.81 ± 0.05 c,d,e,f | 8.88 ± 0.51 i,j,k | 46.33 ± 1.27 c |
| Aprika | 1.41 ± 0.10 a | 11.44 ± 0.26 h,i,j | 34.90 ± 0.52 e,f |
| Quicky | 1.15 ± 0.06 b | 22.56 ± 0.60 b,c,d | 36.27 ± 0.67 e |
| Federica | 0.89 ± 0.11 b,c,d,e | 18.95 ± 0.78 c,d,e,f | 35.80 ± 0.52 e,f |
| Lofty | 0.93 ± 0.08 b,c,d | 23.81 ± 0.50 b,c,d | 33.93 ± 1.10 e,f |
| Tea | 0.76 ± 0.02 d,e,f,g | 18.47 ± 0.69 e,f,g | 35.20 ± 0.00 e,f |
| Mila | 0.77 ± 0.04 d,e,f,g | 20.97 ± 0.20 c,d,e | 34.60 ± 1.04 e,f |
| Arianna | 0.90 ± 0.03 b,c,d,e | 26.05 ± 0.53 a,b | 35.80 ± 0.52 e,f |
| Sandra | 0.87 ± 0.03 b,c,d,e | 30.60 ± 0.15 a | 36.57 ± 0.75 e |
| Albion | 0.71 ± 0.01 e,f,g,h | 11.18 ± 1.00 h,i,j | 48.10 ± 2.08 c |
| Capri | 0.69 ± 0.05 d,e,f,g,h | 5.86 ± 0.26 k,l | 41.04 ± 2.01 d |
| Irma | 0.46 ± 0.01 h | 9.11 ± 0.41 i,j,k | 42.23 ± 1.20 d |
| Cultivars | •OH | DPPH |
|---|---|---|
| Roxana | 93.85 | 54.16 |
| Arosa | 95.53 | 61.42 |
| Joly | 94.71 | 70.36 |
| Asia | 94.78 | 62.75 |
| Alba | 94.71 | 46.39 |
| Jeny | 94.90 | 61.20 |
| Laetitia | 94.82 | 68.32 |
| Garda | 95.31 | 47.71 |
| Lycia | 94.71 | 56.98 |
| Premy | 94.04 | 71.61 |
| Sibilla | 94.63 | 61.56 |
| Vivaldi | 95.27 | 65.11 |
| Rumba | 94.23 | 62.89 |
| Clery | 94.37 | 68.13 |
| Aprika | 93.63 | 63.95 |
| Quicky | 93.74 | 62.89 |
| Federica | 93.67 | 59.71 |
| Lofty | 94.67 | 66.79 |
| Tea | 94.97 | 78.88 |
| Mila | 94.34 | 53.00 |
| Arianna | 93.52 | 48.60 |
| Sandra | 94.04 | 63.98 |
| Albion | 94.71 | 59.06 |
| Capri | 94.49 | 58.63 |
| Irma | 94.00 | 58.77 |
| Cultivars | Pelargonidin-3-O-glucoside | Pelargonidin-3-O-acetylglucoside (P-3-G Equiv.) | Pelargonidin-3-O-rutinoside (P-3-G Equiv.) | Pelargonidin-3-O-malonylglucoside (P-3-G Equiv.) | Cyanidin-3-O-glucoside |
|---|---|---|---|---|---|
| Roxana | 264.27 ± 21.64 f,g,h | 5.20 ± 0.82 b | 23.80 ± 2.12 a,b | 1.90 ± 0.85 i,j | 3.20 ± 0.57 b,c,d,e,f,g,h |
| Arosa | 230.88 ± 36.62 d,e,f,g | 3.03 ± 0.00 c | 20.35 ± 9.83 c,d,e,f | 39.15 ± 0.21 c,d,e | 1.60 ± 0.28 h |
| Joly | 492.70 ± 12.09 a,b | n.d. | 17.60 ± 0.28 c,d,e | 156.58 ± 3.82 a | 2.55 ± 0.21 e,f,g,h |
| Asia | 487.08 ± 24.33 a | 8.30 ±0.66 a | 22.33 ± 1.63 b,c | 2.10 ± 0.21 i,j | 1.45 ± 0.21 h |
| Alba | 218.78 ± 8.63 h,i,j | 0.67 ± 0.14 e | 12.63 ± 1.81 e,f,g,h | 39.40 ± 1.27 b,c | 3.95 ± 0.64 b,c,d,e,f,g,h |
| Jeny | 81.14 ± 7.57 k | n.d. | 0.10 ± 0.00 l | 22.95 ± 0.21 e,f,g | 2.65 ± 0.92 e,f,g,h |
| Laetitia | 357.81 ± 27.65 b,c,d,e,f | 0.33 ± 0.06 e | 31.10 ± 1.98 a | 37.50 ± 0.21 b,c,d | 3.20 ± 0.28 d,e,f,g,h |
| Garda | 191.63 ± 14.07 j | 0.27 ± 0.00 e | 22.80 ± 0.14 b,c,d | 44.80 ± 3.18 b | 4.67 ± 0.67 b,c,d,e,f |
| Lycia | 206.35 ± 17.23 i,j | 0.40 ± 0.07 e | 10.69 ± 0.21 e,f,g,h | 38.90 ± 2.76 b,c | 5.42 ± 1.19 b,c,d,e,f,g |
| Premy | 190.57 ± 8.51 i,j | n.d. | 13.07 ± 0.40 e,f,g,h | 12.27 ± 0.49 f,g,h,i,j | 5.60 ± 0.20 b,c |
| Sibilla | 171.44 ± 7.13 j | 0.27 ± 0.06 e | 11.29 ± 0.98 e,f,g,h,i | 12.20 ± 0.14 f,g,h,i,j | 2.52 ± 0.79 f,g,h |
| Vivaldi | 301.44 ± 10.89 e,f,g,h | 0.50 ± 0.07 e | 10.83 ± 1.19 e,f,g,h,i,j | 24.53 ± 0.85 d,e,f | 4.95 ± 0.79 b,c,d,e |
| Rumba | 222.20 ± 36.43 h,i,j | 0.13 ± 0.06 e | 12.51 ± 0.79 e,f,g,h | 34.87 ± 3.75 b,c,d | 1.03 ± 0.00 h |
| Clery | 42.00 ± 6.25 l | n.d. | 3.36 ± 0.59 k | 3.00 ± 0.49 h,i,j | 6.16 ± 0.40 b |
| Aprika | 182.36 ± 1.98 i,j | n.d. | 15.12 ± 2.91 c,d,e,f,g | 12.37 ± 0.35 f,g,h,i,j | 6.58 ± 1.78 b |
| Quicky | 433.96 ± 8.91 a,b,c | 2.40 ± 0.42 c,d | 12.60 ± 2.12 e,f,g,h | 20.40 ± 1.30 e,f,g | 3.10 ± 0.21 c,d,e,f,g,h |
| Federica | 343.77 ± 34.15 c,d,e,f,g | 6.10 ± 0.21 b | 5.25 ± 1.91 j,k,l | 2.40 ± 0.85 i,j | 10.60 ± 1.91 a |
| Lofty | 301.47 ± 8.91 e,f,g,h | n.d. | 17.40 ± 1.27 c,d,e,f | 12.20 ± 1.13 f,g,h,i,j | 1.20 ± 0.21 h |
| Tea | 276.97 ± 3.82 g,h,i | 3.00 ± 0.42 c | 19.00 ± 1.91 c,d,e | n.d. | 3.40 ± 0.46 b,c,d,e,f,g,h |
| Mila | 334.77 ± 1.48 d,e,f,g | n.d. | 27.40 ± 0.85 a,b | 11.90 ± 0.28 f,g,h,i,j | 4.30 ± 0.46 b,c,d,e,f,g |
| Arianna | 424.76 ± 8.10 a,b,c,d | 0.70 ± 0.00 e | 8.25 ± 2.12 h,i,j,k,l | 15.17 ± 0.67 e,f,g,h,i | 2.40 ± 0.85 g,h |
| Sandra | 395.36 ± 7.65 b,c,d,e | n.d. | 4.50 ± 0.79 i,j,k,l | 16.96 ± 0.42 e,f,g,h | 5.90 ± 0.42 b,c |
| Albion | 257.62 ± 54.65 e,f,g,h | 0.90 ± 0.35 e | 23.20 ± 7.07 c,d,e,f | 54.89 ± 15.70 b | 9.50 ± 0.99 a |
| Capri | 199.03 ± 5.73 h,i,j | 1.80 ± 0.14 c,d | 12.50 ± 0.57 d,e,f,g,h | 7.20 ± 1.70 g,h,i,j | 3.80 ± 0.17 b,c,d,e,f,g,h |
| Irma | 202.43 ± 4.60 h,i,j | 1.75 ± 0.07 c,d | 9.75 ± 0.92 g,h,i,j,k | 1.60 ± 0.42 i,j | 4.20 ± 0.28 b,c,d,e |
| Cultivars | Ellagic Acid | Ellagic Acid Deoxyhexoside (EA Equiv.) | p-Coumaric Acid | p-Coumaroyl Glucose (CA Equiv.) |
|---|---|---|---|---|
| Roxana | 18.60 ± 0.71 c,d,e,f,g | 15.18 ± 0.85 e,f | 1.87 ± 0.40 h,i,j | 122.37 ± 11.54 d,e,f |
| Arosa | 13.90 ± 6.08 b,c,d,e,f | 21.35 ± 3.04 b,c,d | 2.87 ± 0.51 f,g,h | 96.23 ± 9.42 f,g |
| Joly | 33.40 ± 4.81 b,c,d | 34.23 ± 4.31 a | 1.55 ± 0.07 i,j | 182.97 ± 15.77 c |
| Asia | 30.60 ± 1.61 a,b,c | 21.51 ± 0.42 b,c,d | 4.27 ± 0.40 e,f | 302.17 ± 1.65 a |
| Alba | 13.33 ± 0.25 e,f,g | 7.29 ± 0.26 g,h | 4.25 ± 0.21 e,f | 135.35 ± 3.06 c,d,e |
| Jeny | 18.15 ± 2.33 b,c,d,e,f | 16.95 ± 0.14 d,e | 4.10 ± 0.71 e,f | 112.54 ± 20.72 e,f |
| Laetitia | 30.95 ± 6.72 a | 23.58 ± 0.92 b | 3.80 ± 0.71 f,g | 252.72 ± 28.25 b |
| Garda | 7.90 ± 0.44 g,h | 9.39 ± 0.42 g | 2.05 ± 0.21 h,i,j | 164.49 ± 3.77 c |
| Lycia | 13.73 ± 0.00 c,d,e,f,g | 17.13 ± 0.49 c,d,e | 3.53 ± 0.64 f,g,h | 135.35 ± 14.83 c,d,e |
| Premy | 10.58 ± 0.59 f,g | 6.86 ± 1.55 g,h | 6.57 ± 0.93 c,d,e | 67.59 ± 0.28 h,i,j |
| Sibilla | 10.61 ± 0.72 f,g | 3.40 ± 0.95 h | 6.17 ± 0.67 d,e | 65.94 ± 1.91 h,i,j |
| Vivaldi | 12.63 ± 0.59 e,f,g | 2.83 ± 0.55 h | 11.37 ± 0.78 a | 111.72 ± 2.83 e,f |
| Rumba | 14.48 ± 1.53 e,f,g | 2.96 ± 0.81 h | 11.37 ± 1.20 a | 63.33 ± 3.69 h,i,j |
| Clery | 31.23 ± 3.14 a,b | 3.20 ± 0.52 h | 1.20 ± 0.14 j | 75.03 ± 3.32 g,h,i |
| Aprika | 11.95 ± 1.18 e,f,g | 6.76 ± 1.34 g,h | 6.60 ± 0.07 c,d,e | 65.94 ± 2.76 h,i,j |
| Quicky | 14.45 ± 0.78 c,d,e,f,g | 3.00 ± 0.80 h | 8.53 ± 0.35 b | 55.79 ± 6.79 j,k |
| Federica | 5.55 ± 0.64 h | 1.70 ± 0.61 h | 7.50 ± 0.71 c,d | 35.76 ± 4.21 l |
| Lofty | 21.10 ± 2.12 d,e,f,g | 2.13 ± 0.25 h | 5.83 ± 0.23 d,e | 41.96 ± 2.80 k,l |
| Tea | 11.55 ± 0.07 e,f,g | 1.27 ± 0.15 h | 5.73 ± 0.40 d,e | 86.79 ± 1.41 g,h |
| Mila | 9.60 ± 1.48 g | 1.83 ± 0.06 h | 6.43 ± 0.21 c,d,e | 73.04 ± 1.34 g,h,i |
| Arianna | 9.90 ± 0.49 f,g | 2.13 ± 0.29 h | 8.73 ± 0.38 b | 74.34 ± 1.06 g,h,i |
| Sandra | 14.70 ± 2.12 e,f,g | 1.73 ± 0.06 h | 8.10 ± 0.14 b,c | 61.34 ± 0.92 i,j,k |
| Albion | 18.15 ± 0.07 c,d,e,f,g | 10.72 ± 0.21 f,g | 1.23 ± 0.29 j | 73.92 ± 10.36 g,h,i |
| Capri | 22.80 ± 3.11 b,c,d,e | 22.84 ± 0.07 b,c | 6.00 ± 0.85 d,e | 171.48 ± 11.30 c |
| Irma | 9.55 ± 2.76 f,g | 9.66 ± 1.06 g | 2.93 ± 0.06 f,g,h | 220.92 ± 20.48 b |
| Cultivars | Catechin | Quercetin-3-glucuronide (Querc Equiv.) | Kaempferol-3-acetylglucoside (Kaempf Equiv.) | Kaempferol-3-glucoside (Kaempf Equiv.) | Kaempferol-3-glucuronide (Kaempf Equiv.) | Kaempferol-3-coumaroyglucoside (Kaempf Equiv.) |
|---|---|---|---|---|---|---|
| Roxana | 43.56 ± 3.82 d,e,f,g | 5.95 ± 0.21 i,j | 1.15 ± 0.07 f,g | 6.34 ± 0.56 l | 3.32 ± 0.20 h | 20.83 ± 4.42 e,f |
| Arosa | 35.69 ± 3.85 g,h,i,j | 15.30 ± 3.68 g | 1.20 ± 0.17 f,g | 5.94 ± 1.56 l | 18.26 ± 1.93 f | 10.61 ± 0.28 g,h |
| Joly | 75.92 ± 1.62 a | 11.60 ± 1.31 h,i | 1.80 ± 0.14 e,f | 42.08 ± 0.25 e,f | 77.97 ± 4.98 c | 30.56 ± 1.32 d |
| Asia | 76.73 ± 3.78 a | 8.77 ± 0.47 h,i | 1.67 ± 1.07 e,f | 16.09 ± 0.49 h,i | 3.66 ± 0.40 h,i | 9.33 ± 0.23 h |
| Alba | 52.27 ± 6.35 c,d | 11.05 ± 1.06 h | 1.40 ± 0.30 f,g | 13.14 ± 0.10 i,j | 9.59 ± 0.09 g | 14.51 ± 2.70 g |
| Jeny | 22.10 ± 0.84 j | 40.25 ± 1.77 d | 1.25 ± 0.07 f,g | 20.50 ± 3.27 g,h | 14.53 ± 0.71 f,g | 21.48 ± 6.61 e,f |
| Laetitia | 66.56 ± 2.96 b | 23.50 ± 1.98 f | 1.57 ± 0.15 e,f | 28.58 ± 0.13 g | 15.83 ± 0.83 f,g | 20.97 ± 5.65 e,f |
| Garda | 27.44 ± 1.34 i,j | 11.95 ± 1.48 h | 1.80 ± 0.42 e,f | 3.95 ± 0.21 l | 8.22 ± 0.62 g,h | 9.76 ± 2.23 h |
| Lycia | 30.92 ± 1.59 h,i,j,k | 19.81 ± 2.37 f,g | 0.94 ± 0.07 g,h | 19.76 ± 2.85 g,h | 7.76 ± 0.56 g | 48.03 ± 2.39 c |
| Premy | 29.77 ± 0.76 h,i,j | 19.49 ± 2.96 f,g | 2.44 ± 0.28 e | 16.23 ± 2.88 h,i | 73.05 ± 0.95 c | 3.22 ± 0.22 i |
| Sibilla | 27.31 ± 1.53 i,j | 20.39 ± 1.80 f,g | 1.46 ± 0.49 f,g | 17.86 ± 0.21 h,i | 44.00 ± 2.88 e | 3.49 ± 0.20 i |
| Vivaldi | 50.72 ± 1.83 c,d | 6.12 ± 2.04 i,j | 1.36 ± 0.12 f,g | 19.22 ± 0.76 g,h | 67.81 ± 3.01 c,d | 3.92 ± 0.65 i |
| Rumba | 36.60 ± 4.37 g,h,i,j | 8.02 ± 1.02 h | 0.80 ± 0.11 h | 51.30 ± 5.25 e | 72.56 ± 3.76 c | 4.41 ± 0.22 i |
| Clery | 6.95 ± 0.22 k | 2.59 ± 0.49 j | 1.70 ± 0.40 e,f,g | 47.51 ± 1.52 e,f | 48.28 ± 2.10 d,e | 3.42 ± 0.24 i |
| Aprika | 28.98 ± 0.44 i,j | 15.90 ± 0.54 h | 0.94 ± 0.08 g,h | 12.95 ± 1.16 i,j | 64.78 ± 2.02 c | 4.12 ± 0.65 i |
| Quicky | 54.89 ± 3.96 c | 19.33 ± 0.84 f,g | 16.86 ± 5.21 b | 77.86 ± 0.39 c,d | 20.05 ± 3.37 f | 32.94 ± 9.10 d |
| Federica | 43.33 ± 3.24 d,e,f,g | 31.36 ± 5.51 e | 15.40 ± 2.13 b | 80.12 ± 17.55 b,c | 37.14 ± 0.10 e | 99.31 ± 7.37 b |
| Lofty | 37.60 ± 0.44 g,h,i,j | 73.74 ± 0.64 b | 20.38 ± 1.52 a | 48.09 ± 1.61 e,f | 60.16 ± 4.66 c,d | 27.10 ± 9.34 d,e |
| Tea | 48.11 ± 1.51 c,d.e | 76.29 ± 0.28 a | 13.86 ± 2.16 b,c | 63.36 ± 16.45 d,e | 81.31 ± 3.13 c | 98.94 ± 17.75 b |
| Mila | 39.61 ± 1.07 f,g,h,i | 50.73 ± 4.60 c | 10.29 ± 2.01 c,d | 127.09 ± 27.73 a | 294.50 ± 5.39 a | 108.57 ± 51.59 a |
| Arianna | 51.89 ± 0.31 c,d | 29.66 ± 0.07 e,f | 6.15 ± 0.27 d | 83.58 ± 2.48 b,c | 100.09 ± 14.54 b | 26.34 ± 12.21 d,e |
| Sandra | 31.33 ± 1.27 h,i,j,k | 34.00 ±1.06 e | 7.92 ± 0.50 d | 79.56 ± 37.99 b,c | 9.07 ± 1.47 g | 41.26 ± 13.03 c |
| Albion | 32.73 ± 5.85 h,i,j | 18.90 ± 0.92 f,g | 1.35 ± 0.07 f,g | 20.18 ± 1.94 g,h | 14.07 ± 3.46 f,g | 30.05 ± 6.22 d |
| Capri | 41.23 ± 2.41 e,f,g,h | 33.50 ± 0.42 e,f | 6.44 ± 3.04 d | 51.58 ± 0.47 e | 1.62 ± 0.43 j | 11.88 ± 2.37 g,h |
| Irma | 37.28 ± 1.17 g,h,i,j | 11.95 ± 0.78 h,i | 1.65 ± 0.07 e,f | 11.14 ± 0.58 j | 5.36 ± 0.29 h | 37.65 ± 5.09 c,d |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milosavljević, D.M.; Maksimović, V.M.; Milivojević, J.M.; Nakarada, Đ.J.; Mojović, M.D.; Dragišić Maksimović, J.J. Rich in Phenolics—Strong Antioxidant Fruit? Comparative Study of 25 Strawberry Cultivars. Plants 2022, 11, 3566. https://doi.org/10.3390/plants11243566
Milosavljević DM, Maksimović VM, Milivojević JM, Nakarada ĐJ, Mojović MD, Dragišić Maksimović JJ. Rich in Phenolics—Strong Antioxidant Fruit? Comparative Study of 25 Strawberry Cultivars. Plants. 2022; 11(24):3566. https://doi.org/10.3390/plants11243566
Chicago/Turabian StyleMilosavljević, Dragica M., Vuk M. Maksimović, Jasminka M. Milivojević, Đura J. Nakarada, Miloš D. Mojović, and Jelena J. Dragišić Maksimović. 2022. "Rich in Phenolics—Strong Antioxidant Fruit? Comparative Study of 25 Strawberry Cultivars" Plants 11, no. 24: 3566. https://doi.org/10.3390/plants11243566