
Impacts of Heat Stress around Flowering on Growth and Development Dynamic of Maize (Zea mays L.) Ear and Yield Formation
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
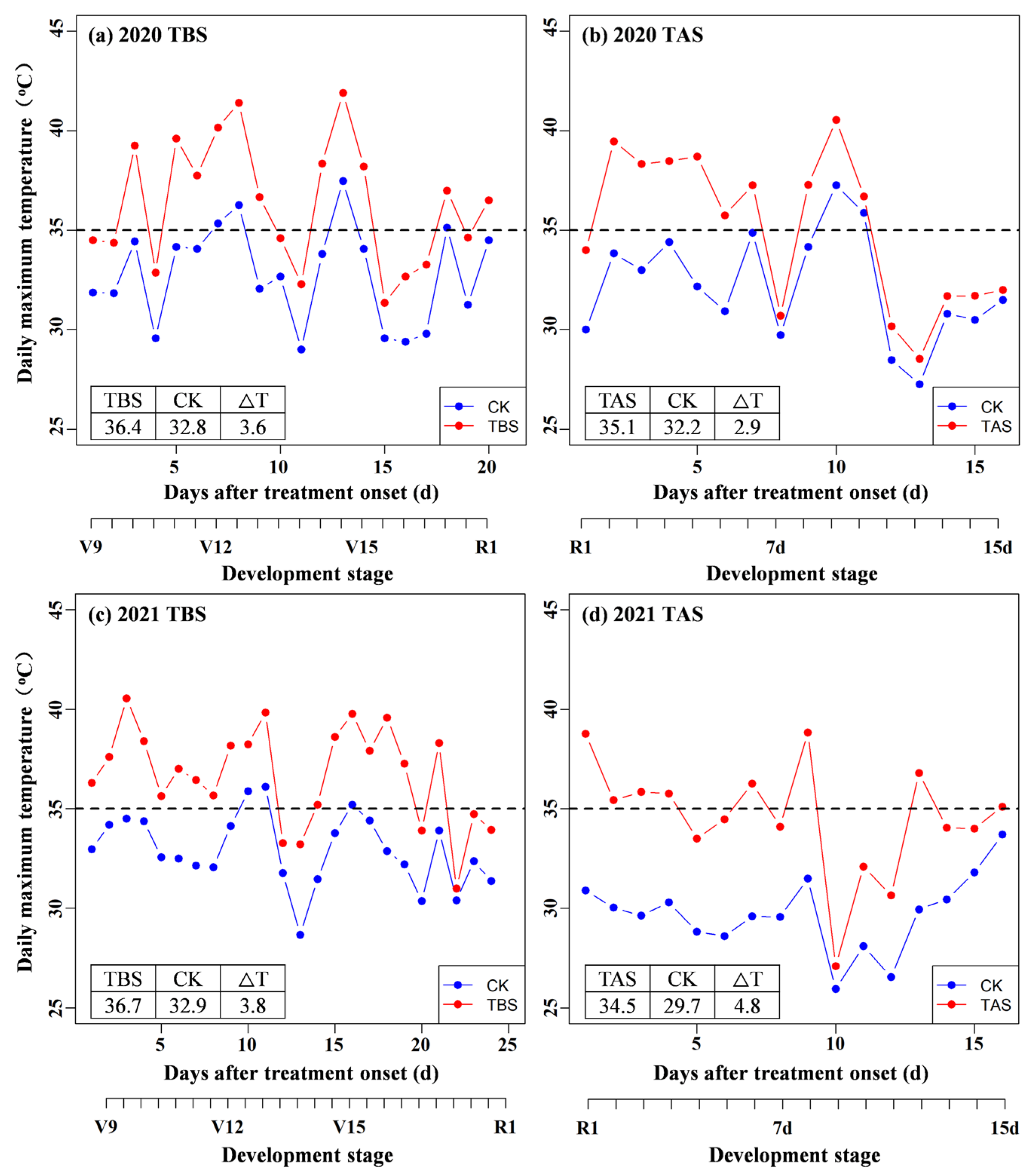
2.1. Crop Husbandry and Experimental Design
2.2. Sampling and Measurements
2.2.1. Ear Length, Ear Diameter, and Ear Dry Matter Weight
2.2.2. Floret Differentiation and Fertilization
2.2.3. Yield Components
2.2.4. Logistic Function and Feature Parameters
2.3. Statistical Analyses
3. Results
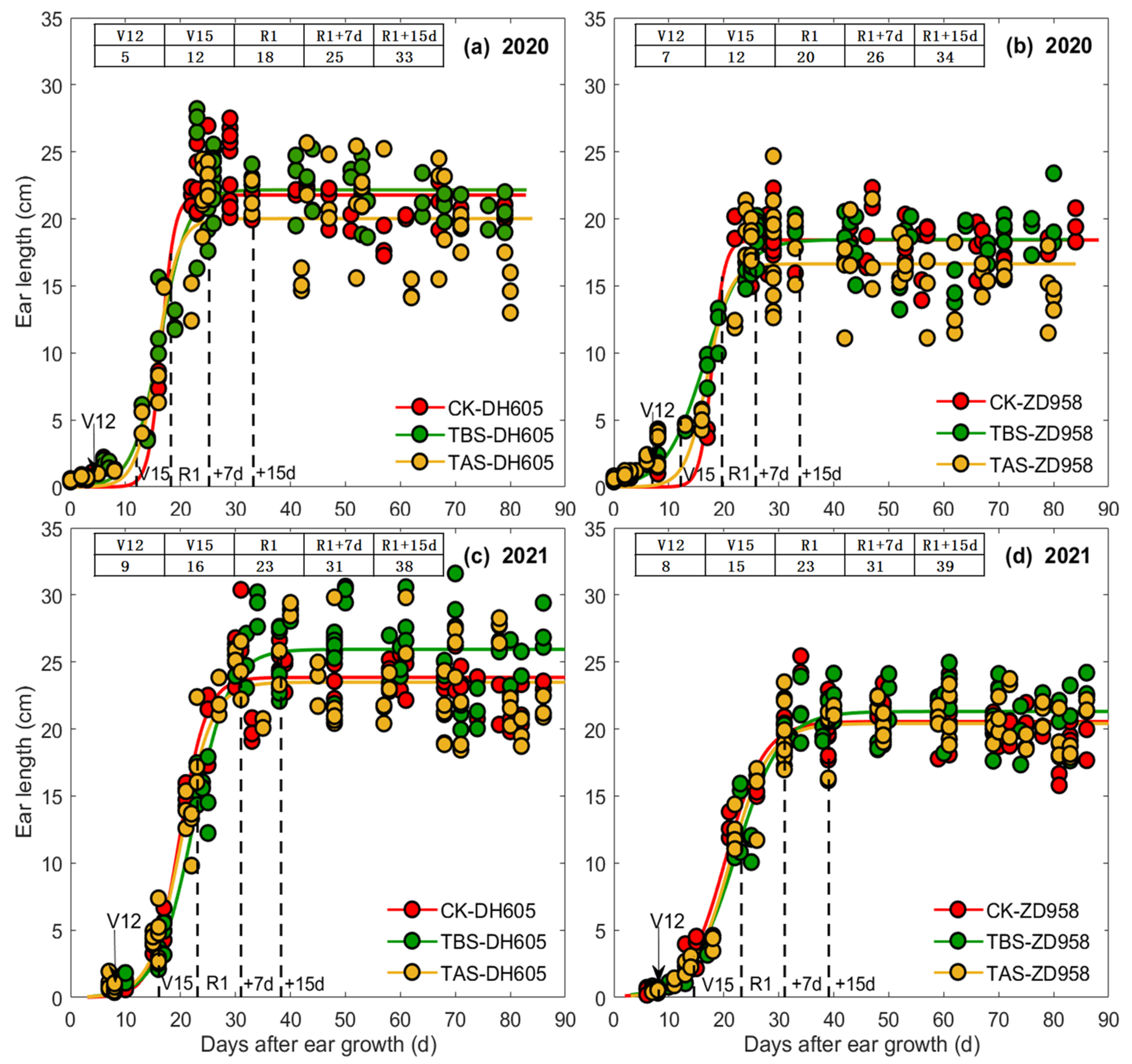
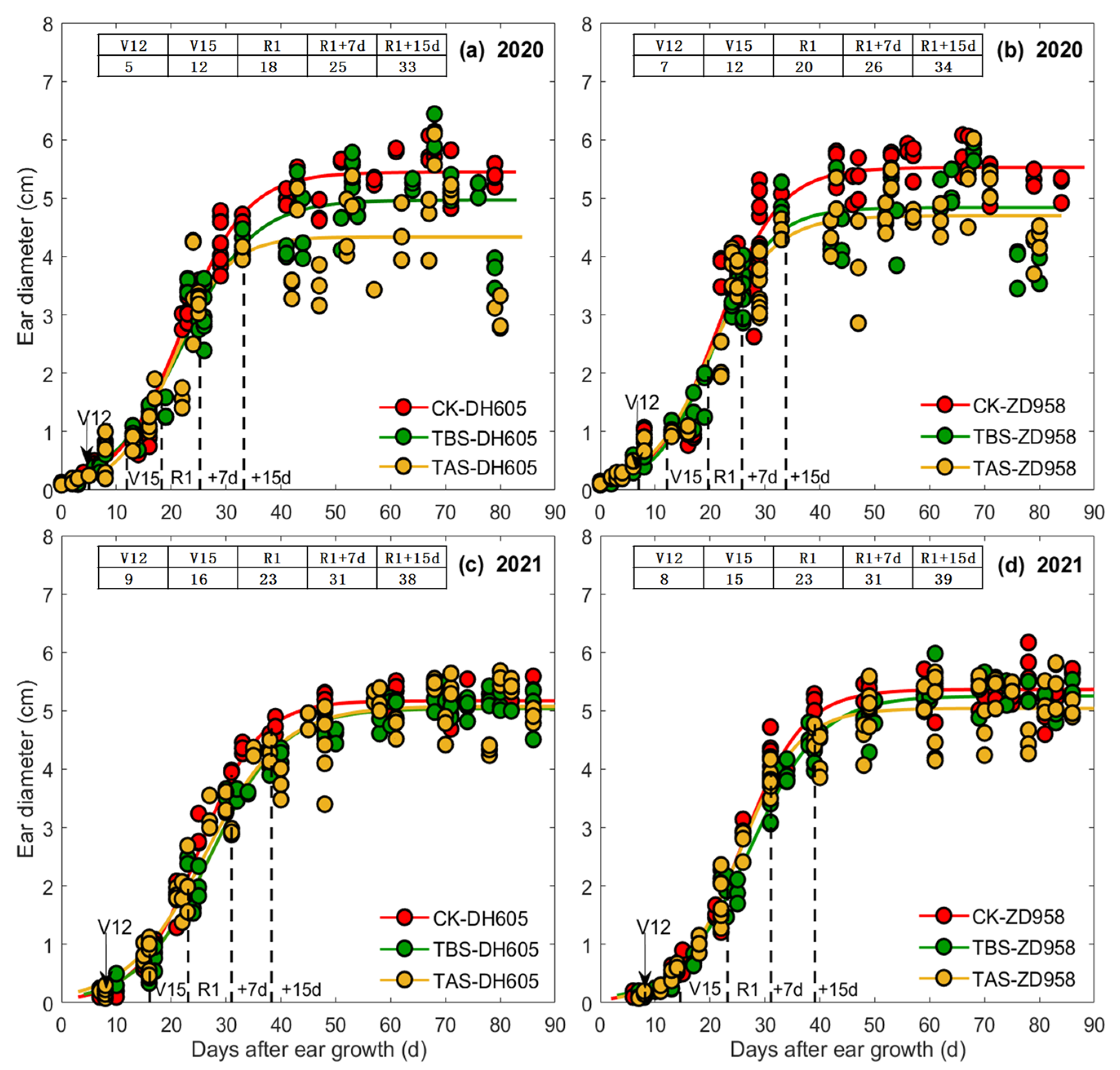
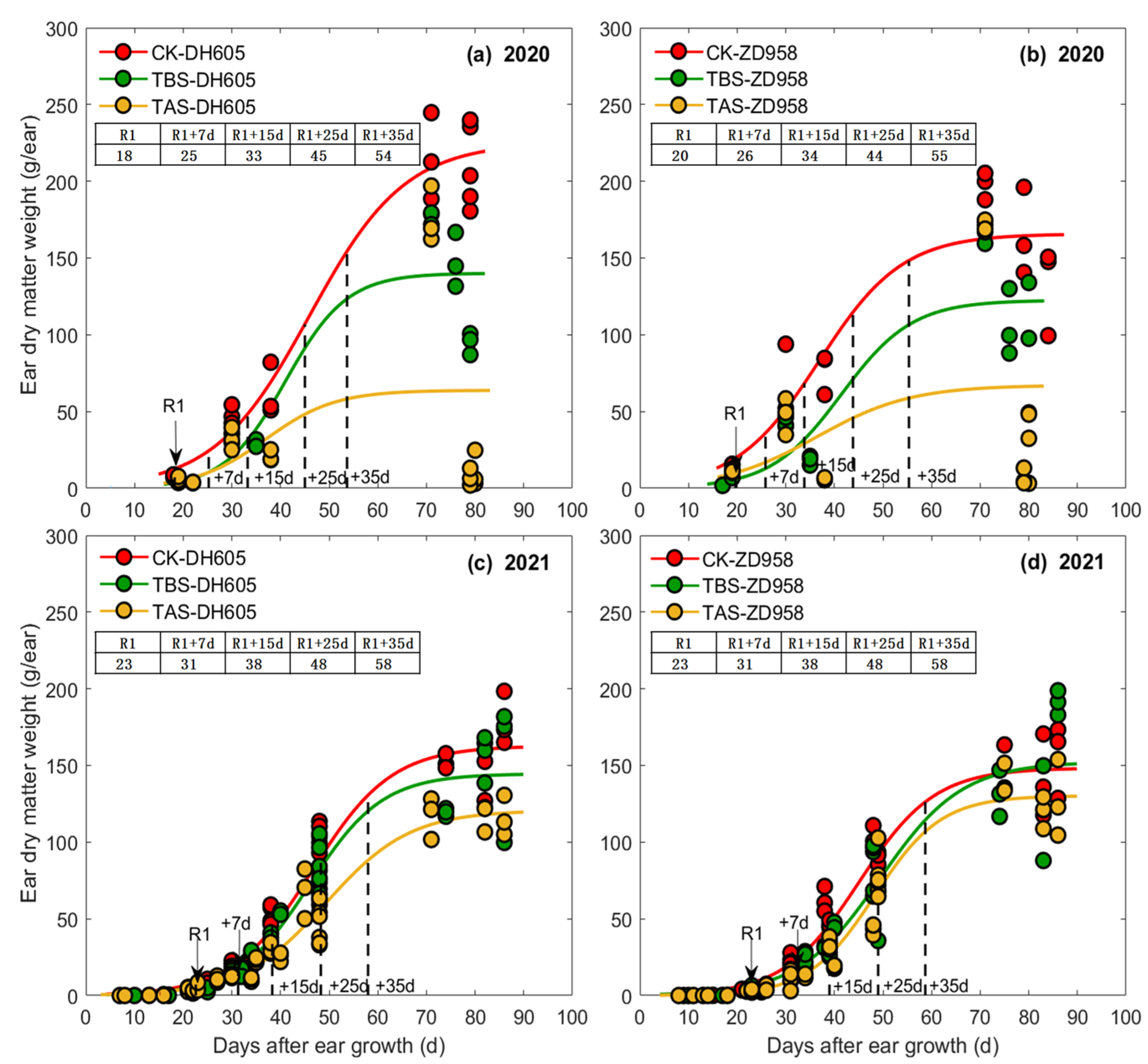
3.1. Effects of Heat Stress on Ear Length, Diameter, and Ear Dry Matter Weight
3.2. Effects of Heat Stress on Kernel Number, Kernel Weight, Yield Formation and Biomass Accumulation
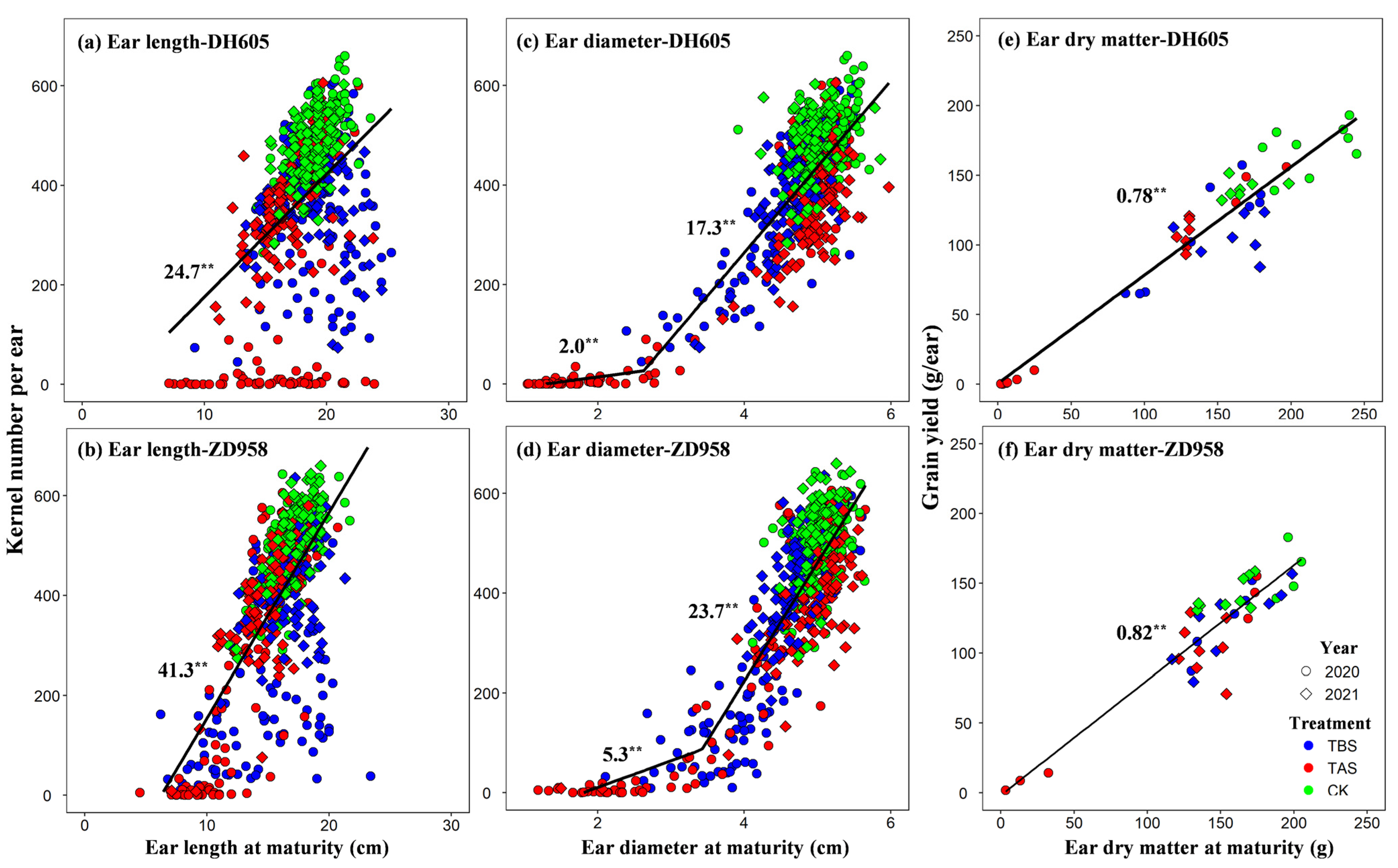
3.3. Relationship between Ear Morphological Traits and Yield Components
4. Discussion
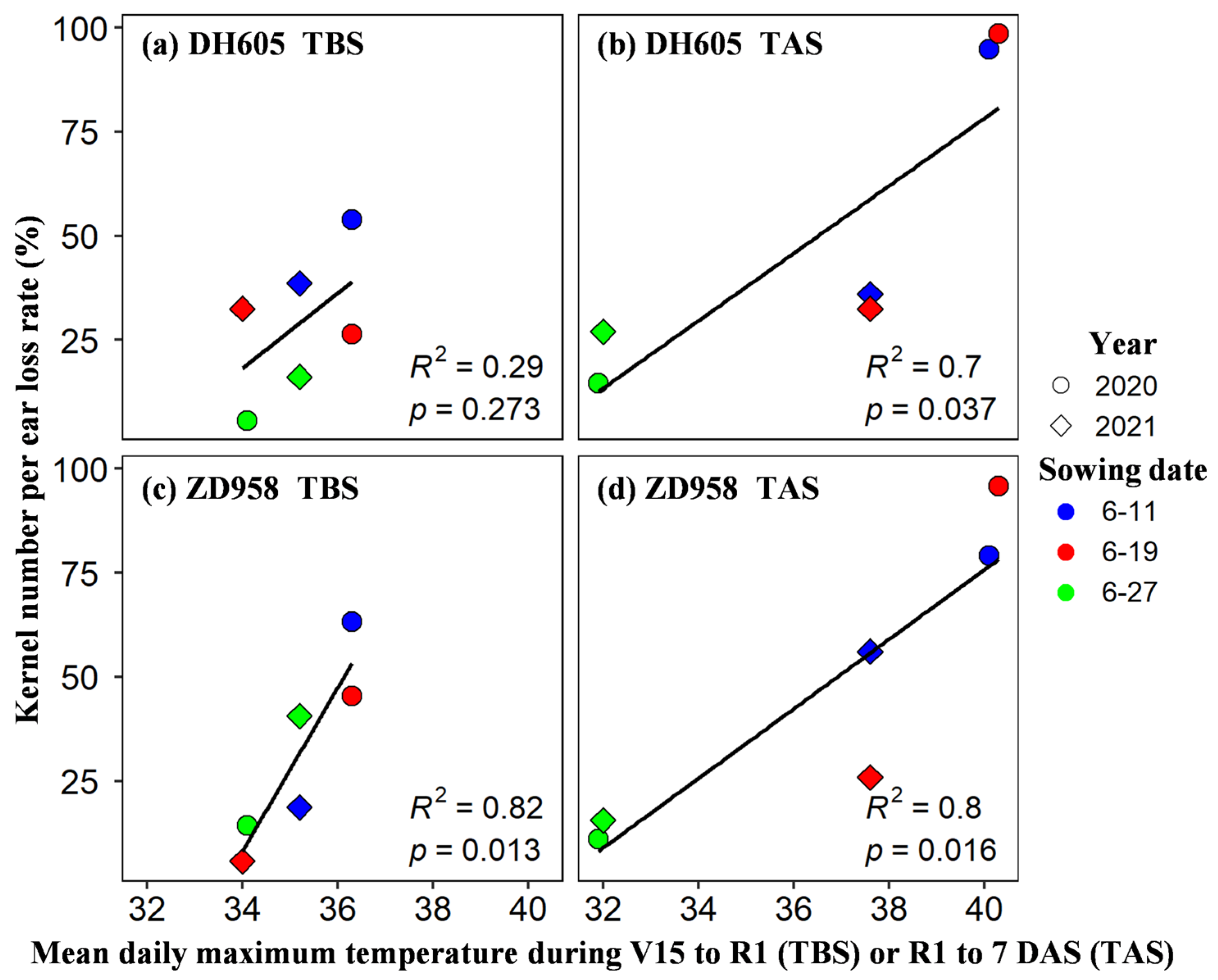
4.1. Effects of Different Levels of Heat Stress around Silking on KNP
4.2. Effects and Mechanism of Heat Stress around Silking on Ear Length, Ear Diameter and Ear Dry Matter
4.3. Effects and Mechanism of Heat Stress around Silking on Kernel Number
4.4. Effects and Mechanism of Heat Stress around Silking on Kernel Weight
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akram, M.; Wahid, A.; Abrar, M.; Manan, A.; Naeem, S.; Zahid, M.; Gilani, M.; Paudyal, R.; Gong, H.; Ran, J. Comparative study of six maize (Zea mays L.) cultivars concerning cadmium uptake, partitioning and tolerance. Appl. Ecol. Environ. Res. 2021, 19, 2305–2331. [Google Scholar] [CrossRef]
- Rouf Shah, T.; Prasad, K.; Kumar, P. Maize—A potential source of human nutrition and health: A review. Cogent Food Agric. 2016, 2, 1166995. [Google Scholar] [CrossRef]
- Liu, M.; Sheng, D.; Liu, X.; Wang, Y.; Hou, X.; Wang, Y.; Wang, P.; Guan, L.; Dong, X.; Huang, S. Dissecting heat tolerance and yield stability in maize from greenhouse and field experiments. J. Agron. Crop Sci. 2022, 208, 348–361. [Google Scholar] [CrossRef]
- Huang, M.; Wang, J.; Wang, B.; Li Liu, D.; Yu, Q.; He, D.; Wang, N.; Pan, X. Optimizing sowing window and cultivar choice can boost China’s maize yield under 1.5 °C and 2 °C global warming. Environ. Res. Lett. 2020, 15, 024015. [Google Scholar] [CrossRef]
- Jodage, K.; Kuchanur, P.; Zaidi, P.; Patil, A.; Seetharam, K.; Vinayan, M.; Arunkumar, B. Association and path analysis for grain yield and its attributing traits under heat stress condition in tropical maize (Zea mays L.). Electron. J. Plant Breed. 2017, 8, 336–341. [Google Scholar] [CrossRef]
- Cicchino, M.; Edreira, J.I.R.; Otegui, M.E. Heat stress during late vegetative growth of maize: Effects on phenology and assessment of optimum temperature. Crop Sci. 2010, 50, 1431–1437. [Google Scholar] [CrossRef]
- Gourdji, S.M.; Mathews, K.L.; Reynolds, M.; Crossa, J.; Lobell, D.B. An assessment of wheat yield sensitivity and breeding gains in hot environments. Proc. Biol. Sci. 2013, 280, 20122190. [Google Scholar] [CrossRef]
- Lizaso, J.; Ruiz-Ramos, M.; Rodriguez, L.; Gabaldon-Leal, C.; Oliveira, J.; Lorite, I.; Sánchez, D.; García, E.; Rodríguez, A. Impact of high temperatures in maize: Phenology and yield components. Field Crop. Res. 2018, 216, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Tian, B.; Zhu, J.; Nie, Y.; Xu, C.; Meng, Q.; Wang, P. Mitigating heat and chilling stress by adjusting the sowing date of maize in the North China Plain. J. Agron. Crop Sci. 2019, 205, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Tucker, S.L.; Dohleman, F.G.; Grapov, D.; Flagel, L.; Yang, S.; Wegener, K.M.; Kosola, K.; Swarup, S.; Rapp, R.A.; Bedair, M. Evaluating maize phenotypic variance, heritability, and yield relationships at multiple biological scales across agronomically relevant environments. Plant Cell Environ. 2020, 43, 880–902. [Google Scholar] [CrossRef]
- Mo, E.; Zhao, R.; Li, Y.; Wang, J.; Liu, F.; Du, Y. Study on growth dynamic of waxy corn ear based on Logistic model. Chin. Agric. Sci. Bull. 2012, 28, 49–52. (In Chinese) [Google Scholar]
- Cicchino, M.; Edreira, J.; Uribelarrea, M.; Otegui, M. Heat stress in field-grown maize: Response of physiological determinants of grain yield. Crop Sci. 2010, 50, 1438–1448. [Google Scholar] [CrossRef]
- Neiff, N.; Trachsel, S.; Valentinuz, O.R.; Balbi, C.N.; Andrade, F.H. High temperatures around flowering in maize: Effects on photosynthesis and grain yield in three genotypes. Crop Sci. 2016, 56, 2702–2712. [Google Scholar] [CrossRef]
- Li, N.; Hao, K.; Sha, B.; Liang, G. Study on growth dynamics of maize ear after anthesis based on image processing. Chin. Agric. Sci. Bull. 2014, 30, 234–240. (In Chinese) [Google Scholar]
- Zhao, L.; Li, C.; Liu, T.; Wang, X.; Seng, S. Effect of high temperature during flowering on photosynthetic characteristics and grain yield and quality of different genotypes of maize (Zea mays L.). Sci. Agric. Sin. 2012, 45, 4947–4958. (In Chinese) [Google Scholar]
- Wang, H.Q.; Liu, P.; Zhang, J.W.; Zhao, B.; Ren, B.Z. Endogenous hormones inhibit differentiation of young ears in maize (Zea mays L.) under heat stress. Front. Plant Sci. 2020, 11, 1655–1669. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Yu, K.; Li, H.; Jia, S.; Yang, Q.; Zhao, X.; Zhao, Y.; Liu, T. The effect of elevating temperature on the growth and development of reproductive organs and yield of summer maize. J. Integr. Agric. 2021, 20, 1783–1795. [Google Scholar] [CrossRef]
- Edreira, J.R.; Carpici, E.B.; Sammarro, D.; Otegui, M. Heat stress effects around flowering on kernel set of temperate and tropical maize hybrids. Field Crop. Res. 2011, 123, 62–73. [Google Scholar] [CrossRef]
- Shao, J.; Liu, P.; Zhao, B.; Zhang, J.; Zhao, X.; Ren, B. Combined effects of high temperature and waterlogging on yield and stem development of summer maize. Crop J. 2022; in press. [Google Scholar] [CrossRef]
- Niu, S.; Du, X.; Wei, D.; Liu, S.; Tang, Q.; Bian, D.; Zhang, Y.; Cui, Y.; Gao, Z. Heat stress after pollination reduces kernel number in maize by insufficient assimilates. Front. Genet. 2021, 12, 728166. [Google Scholar] [CrossRef]
- Wang, Y.; Sheng, D.; Zhang, P.; Dong, X.; Yan, Y.; Hou, X.; Wang, P.; Huang, S. High temperature sensitivity of kernel formation in different short periods around silking in maize. Environ. Exp. Bot. 2020, 183, 104343. [Google Scholar] [CrossRef]
- Mu, X.; Ma, Z.; Zhang, L.; Fu, J.; Liu, T.; Ding, Y.; Xia, L.; Zhang, F.; Zhang, J.; Qi, Q. Responses of photosynthetic fluorescence characteristics, pollination, and yield components of maize cultivars to high temperature during flowering. Chin. J. Eco-Agric. 2022, 30, 57–71. (In Chinese) [Google Scholar]
- Wang, Y.; Liu, X.; Hou, X.; Sheng, D.; Dong, X.; Gao, Y.; Wang, P.; Huang, S. Maximum lethal temperature for flowering and seed set in maize with contrasting male and female flower sensitivities. J. Agron. Crop Sci. 2021, 207, 679–689. [Google Scholar] [CrossRef]
- Wang, Y.; Tao, H.; Tian, B.; Sheng, D.; Xu, C.; Zhou, H.; Huang, S.; Wang, P. Flowering dynamics, pollen, and pistil contribution to grain yield in response to high temperature during maize flowering. Environ. Exp. Bot. 2019, 158, 80–88. [Google Scholar] [CrossRef]
- Alam, M.A.; Seetharam, K.; Zaidi, P.H.; Dinesh, A.; Vinayan, M.T.; Nath, U.K. Dissecting heat stress tolerance in tropical maize (Zea mays L.). Field Crop. Res. 2017, 204, 110–119. [Google Scholar] [CrossRef]
- Guo, J.; Gu, X.; Lu, W.; Lu, D. Multiomics analysis of kernel development in response to short-term heat stress at the grain formation stage in waxy maize. J. Exp. Bot. 2021, 72, 6291–6304. [Google Scholar] [CrossRef]
- Liu, X.; Yu, Y.; Huang, S.; Xu, C.; Wang, X.; Gao, J.; Meng, Q.; Wang, P. The impact of drought and heat stress at flowering on maize kernel filling: Insights from the field and laboratory. Agric. For. Meteorol. 2022, 312, 108733. [Google Scholar] [CrossRef]
- Huang, M.; Wang, J.; Wang, B.; Li Liu, D.; Feng, P.; Yu, Q.; Pan, X.; Waters, C. Assessing maize potential to mitigate the adverse effects of future rising temperature and heat stress in China. Agric. For. Meteorol. 2021, 311, 108673. [Google Scholar] [CrossRef]
- Edreira, J.I.R.; Mayer, L.I.; Otegui, M.E. Heat stress in temperate and tropical maize hybrids: Kernel growth, water relations and assimilate availability for grain filling. Field Crop. Res. 2014, 166, 162–172. [Google Scholar] [CrossRef]
- Yin, S.; Li, P.; Xu, Y.; Liu, J.; Yang, T.; Wei, J.; Xu, S.; Yu, J.; Fang, H.; Xue, L. Genetic and genomic analysis of the seed-filling process in maize based on a logistic model. Heredity 2020, 124, 122–134. [Google Scholar] [CrossRef]
- Suwa, R.; Hakata, H.; Hara, H.; El-Shemy, H.A.; Adu-Gyamfi, J.J.; Nguyen, N.T.; Kanai, S.; Lightfoot, D.A.; Mohapatra, P.K.; Fujita, K. High temperature effects on photosynthate partitioning and sugar metabolism during ear expansion in maize (Zea mays L.) genotypes. Plant Physiol. Biochem. 2010, 48, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Vanaja, M.; Sathish, P.; Kumar, G.V.; Razzaq, A.; Vagheera, P.; Lakshmi, N.J.; Yadav, S.; Sarkar, B.; Maheswari, M. Elevated temperature and moisture deficit stress impact on phenology, physiology and yield responses of hybrid maize. J. Agrometeorol. 2017, 19, 295–300. [Google Scholar] [CrossRef]
- Fang, Q.; Zhang, X.; Chen, S.; Shao, L.; Sun, H.; Yan, Z. Selecting traits to reduce seasonal yield variation of summer maize in the North China Plain. Agron. J. 2019, 111, 343–353. [Google Scholar] [CrossRef]
- Qiao, J.; Li, P.; Zhang, M.; Li, C.; Zhu, W.; Dai, S.; Huang, L.; Yu, T.; Guo, G.; Niu, J. Effect of high temperature on yield and quality of different summer maize varieties at flowering stage. J. Henan Agric. Sci. 2019, 48, 11–18. [Google Scholar]
- Yan, P.; Chen, Y.; Sui, P.; Vogel, A.; Zhang, X. Effect of maize plant morphology on the formation of apical kernels at different sowing dates and under different plant densities. Field Crop. Res. 2018, 223, 83–92. [Google Scholar] [CrossRef]
- Liu, M.; Dong, X.; Zhang, Y.; Gu, M.; Yu, Y.; Xie, H.; Yang, H.; Yu, X.; Huang, S. Heat stress on maize with contrasting genetic background: Differences in flowering and yield formation. Agric. For. Meteorol. 2022, 319, 108934. [Google Scholar] [CrossRef]
- Cicchino, M.A.; Rattalino Edreira, J.I.; Otegui, M.E. Maize physiological responses to heat stress and hormonal plant growth regulators related to ethylene metabolism. Crop Sci. 2013, 53, 2135–2146. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Cultivar | Treatment | Ear Length (EL) | Ear Diameter (ED) | Ear Dry Matter (EDM) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a | b | c | t1 (d) | t2 (d) | t2 − t1 (d) | vmean (cm d−1) | a | b | c | t1 (d) | t2 (d) | t2 − t1 (d) | vmean (cm d−1) | a | b | c | t1 (d) | t2 (d) | t2 − t1 (d) | vmean (g ear−1 d−1) | |||
| 2020 | DH605 | TBS | 22.2 | 6.4 | 0.39 | 13 | 20 | 7 | 1.9 | 5.0 | 3.4 | 0.16 | 13 | 29 | 16 | 0.18 | 140.1 | 6.5 | 0.16 | 32 | 49 | 17 | 4.9 |
| TAS | 20.0 | 8.2 | 0.52 | 13 | 18 | 5 | 2.3 | 4.3 | 4.1 | 0.20 | 14 | 27 | 13 | 0.19 | 63.7 | 5.2 | 0.14 | 28 | 47 | 19 | 2.0 | ||
| CK | 21.8 | 14.0 | 0.84 | 15 | 18 | 3 | 4.0 | 5.4 | 3.7 | 0.17 | 14 | 30 | 16 | 0.20 | 165.7 | 4.3 | 0.11 | 27 | 51 | 24 | 4.0 | ||
| ZD958 | TBS | 18.5 | 4.6 | 0.28 | 12 | 21 | 9 | 1.1 | 4.8 | 3.9 | 0.19 | 14 | 27 | 13 | 0.21 | 122.5 | 5.7 | 0.14 | 31 | 50 | 19 | 3.8 | |
| TAS | 16.6 | 8.5 | 0.50 | 14 | 20 | 6 | 1.6 | 4.7 | 3.5 | 0.18 | 12 | 27 | 15 | 0.18 | 67.1 | 3.8 | 0.10 | 25 | 51 | 26 | 1.5 | ||
| CK | 18.4 | 16.0 | 0.89 | 16 | 19 | 3 | 3.6 | 5.5 | 3.8 | 0.18 | 14 | 28 | 14 | 0.23 | 225.2 | 4.7 | 0.10 | 34 | 60 | 26 | 4.9 | ||
| 2021 | DH605 | TBS | 25.9 | 6.9 | 0.31 | 18 | 27 | 8 | 1.8 | 5.0 | 4.1 | 0.15 | 19 | 36 | 18 | 0.16 | 144.6 | 6.1 | 0.13 | 37 | 57 | 20 | 4.1 |
| TAS | 23.5 | 7.2 | 0.35 | 17 | 24 | 8 | 1.8 | 5.0 | 4.6 | 0.18 | 18 | 33 | 15 | 0.20 | 86.6 | 6.5 | 0.14 | 37 | 56 | 19 | 2.7 | ||
| CK | 23.9 | 8.2 | 0.41 | 17 | 23 | 6 | 2.1 | 5.2 | 4.5 | 0.18 | 18 | 32 | 15 | 0.21 | 148.3 | 5.8 | 0.13 | 34 | 55 | 21 | 4.2 | ||
| ZD958 | TBS | 21.3 | 5.4 | 0.24 | 17 | 28 | 11 | 1.1 | 5.2 | 4.2 | 0.15 | 19 | 37 | 18 | 0.17 | 152.3 | 5.9 | 0.12 | 38 | 60 | 22 | 4.0 | |
| TAS | 20.4 | 6.0 | 0.28 | 17 | 26 | 9 | 1.3 | 5.1 | 3.7 | 0.14 | 17 | 36 | 19 | 0.16 | 120.7 | 5.7 | 0.11 | 40 | 64 | 24 | 2.9 | ||
| CK | 20.6 | 5.8 | 0.28 | 16 | 25 | 9 | 1.3 | 5.4 | 4.6 | 0.18 | 18 | 33 | 15 | 0.21 | 167.2 | 5.7 | 0.12 | 37 | 58 | 21 | 4.4 | ||
| Cultivar | Year | Treatment | Total Florets | Floret Fertilization Rate (%) | Seed Setting Rate (%) | Kernel Number Per Ear | Kernel-Row Number | Kernel Number Per Row | Kernel Number Loss Rate (%) | 1000-Grain Weight (g) | Grain Yield (g) | Plant dry Matter (g) | Ear dry Matter (g) | The Ratio of Dry Matter Partitioning to the Ear |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2020 | DH605 | TBS | 752 a | 68.7 b | 47.4 a | 370 b | 16.0 a | 24.8 b | 27.2 | 309 ab | 116 b | 252.0 b | 139.7 b | 0.55 b |
| TAS | 756 a | 42.7 c | 24.9 b | 156 c | 11.7 b | 15.8 c | 69.3 | 289 b | 53 c | 214.4 b | 64.9 c | 0.23 c | ||
| CK | 795 a | 90.9 a | 65.0 a | 508 a | 17.2 a | 31.2 a | / | 331 a | 180 a | 339.3 a | 215.0 a | 0.63 a | ||
| ZD958 | TBS | 581 a | 69.5 b | 63.8 a | 365 b | 14.1 ab | 26.7 b | 26.3 | 306 a | 117 b | 258.1 ab | 121.8 b | 0.46 b | |
| TAS | 606 a | 55.5 b | 39.8 b | 243 c | 12.0 b | 21.2 c | 50.9 | 286 b | 62 c | 226.2 b | 64.9 c | 0.25 c | ||
| CK | 628 a | 90.5 a | 78.4 a | 495 a | 14.8 a | 33.8 a | / | 319 a | 179 a | 291.5 a | 181.5 a | 0.62 a | ||
| 2021 | DH605 | TBS | 746 a | 76.7 b | 48.3 b | 372 b | 15.5 a | 26.6 b | 22.0 | 297 a | 131 b | 212.3 b | 140.1 b | 0.66 a |
| TAS | 761 a | 75.5 b | 47.7 b | 364 b | 15.6 a | 26.2 b | 23.7 | 321 a | 119 b | 196.6 b | 126.7 b | 0.65 a | ||
| CK | 767 a | 93.9 a | 62.0 a | 477 a | 15.9 a | 32.0 a | / | 310 a | 163 a | 244.2 a | 163.3 a | 0.67 a | ||
| ZD958 | TBS | 631 a | 83.8 b | 66.8 a | 426 b | 14.8 a | 30.8 b | 17.1 | 284 a | 136 b | 218.0 ab | 133.6 b | 0.61 a | |
| TAS | 702 a | 82.1 b | 56.0 b | 391 c | 14.7 a | 28.4 b | 23.9 | 286 a | 126 b | 205.5 b | 129.1 b | 0.63 a | ||
| CK | 693 a | 91.6 a | 68.2 a | 498 a | 15.2 a | 34.3 a | / | 275 a | 158 a | 236.7 a | 155.6 a | 0.66 a | ||
| Source of variation | Year | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ||
| Cultivar | ** | ** | ** | ** | NS | NS | ** | NS | NS | ** | NS | |||
| Sowing Date | * | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | |||
| Treatment | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | |||
| Year × Cultivar | ** | NS | NS | ** | NS | ** | ** | NS | NS | ** | NS | |||
| Year × Sowing date | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | |||
| Year × Treatment | NS | ** | ** | ** | ** | ** | ** | ** | ** | ** | ** | |||
| Cultivar × Sowing date | NS | NS | ** | NS | NS | ** | ** | * | NS | NS | NS | |||
| Cultivar × Treatment | NS | NS | * | * | NS | NS | ** | NS | NS | ** | NS | |||
| Sowing date × Treatment | NS | ** | ** | ** | NS | ** | ** | ** | ** | ** | ** | |||
| Year × Cultivar × Sowing date | ** | * | ** | ** | NS | * | ** | NS | NS | NS | NS | |||
| Year × Cultivar × Treatment | NS | NS | ** | NS | NS | NS | ** | NS | NS | NS | NS | |||
| Year × Sowing date × Treatment | NS | ** | ** | ** | ** | * | ** | ** | ** | ** | ** | |||
| Cultivar × Sowing date × Treatment | NS | NS | ** | * | NS | NS | ** | ** | * | * | NS | |||
| Year × Cultivar × Sowing date × Treatment | * | NS | ** | ** | * | NS | ** | ** | NS | NS | NS | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Liu, Q.; Ming, B.; Shang, W.; Zhao, X.; Wang, X.; Wang, J.; Zhang, J.; Luo, Z.; Liao, Y. Impacts of Heat Stress around Flowering on Growth and Development Dynamic of Maize (Zea mays L.) Ear and Yield Formation. Plants 2022, 11, 3515. https://doi.org/10.3390/plants11243515
Wang N, Liu Q, Ming B, Shang W, Zhao X, Wang X, Wang J, Zhang J, Luo Z, Liao Y. Impacts of Heat Stress around Flowering on Growth and Development Dynamic of Maize (Zea mays L.) Ear and Yield Formation. Plants. 2022; 11(24):3515. https://doi.org/10.3390/plants11243515
Chicago/Turabian StyleWang, Na, Qi Liu, Bo Ming, Wenxin Shang, Xuefeng Zhao, Xuqing Wang, Jing Wang, Junlong Zhang, Zhongkui Luo, and Yong Liao. 2022. "Impacts of Heat Stress around Flowering on Growth and Development Dynamic of Maize (Zea mays L.) Ear and Yield Formation" Plants 11, no. 24: 3515. https://doi.org/10.3390/plants11243515