
Biochemical Analyses of Bioactive Extracts from Plants Native to Lampedusa, Sicily Minor Island
 , , ,
, , ,  ,
,  , and
, and
Abstract
:1. Introduction
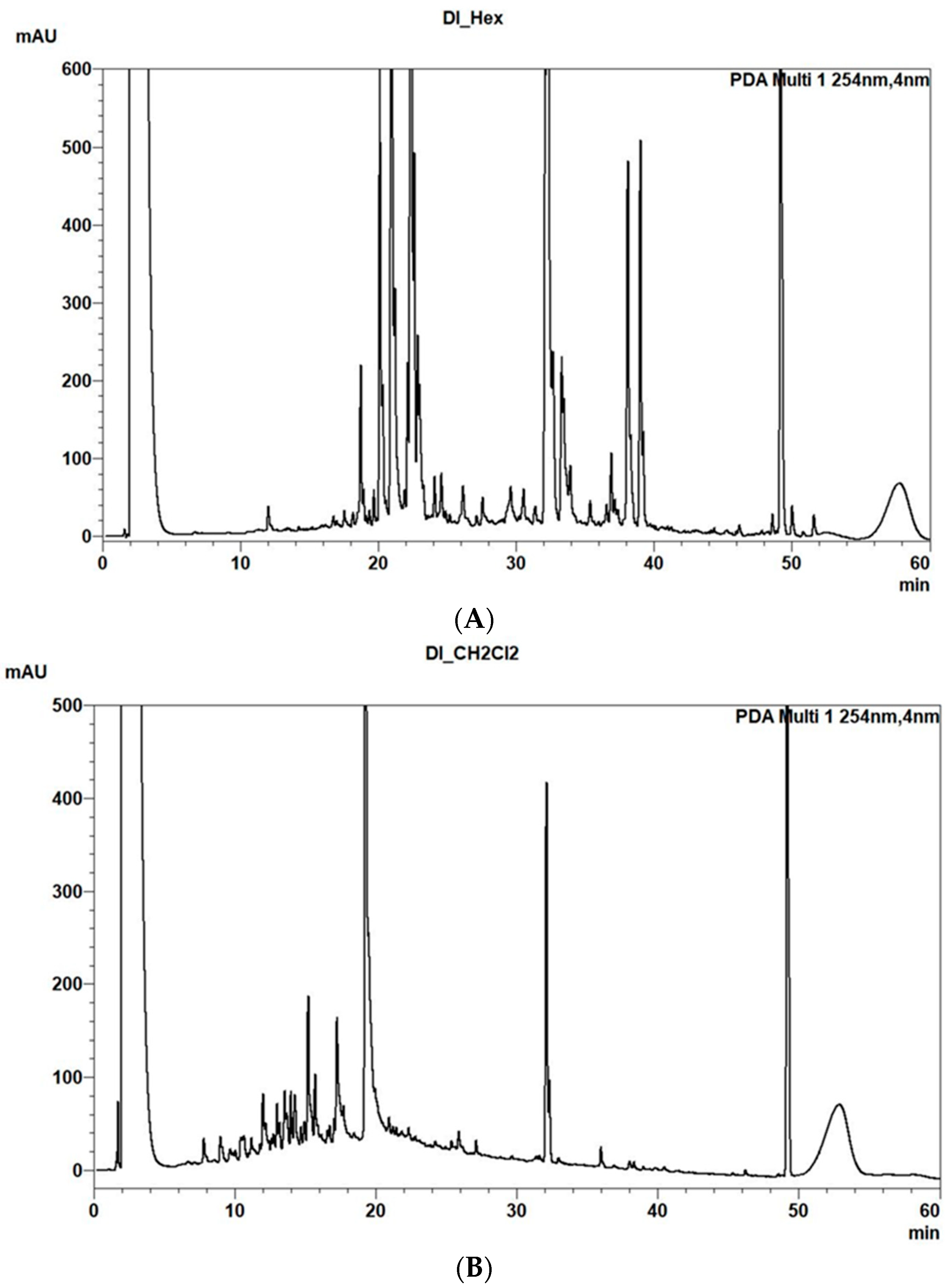
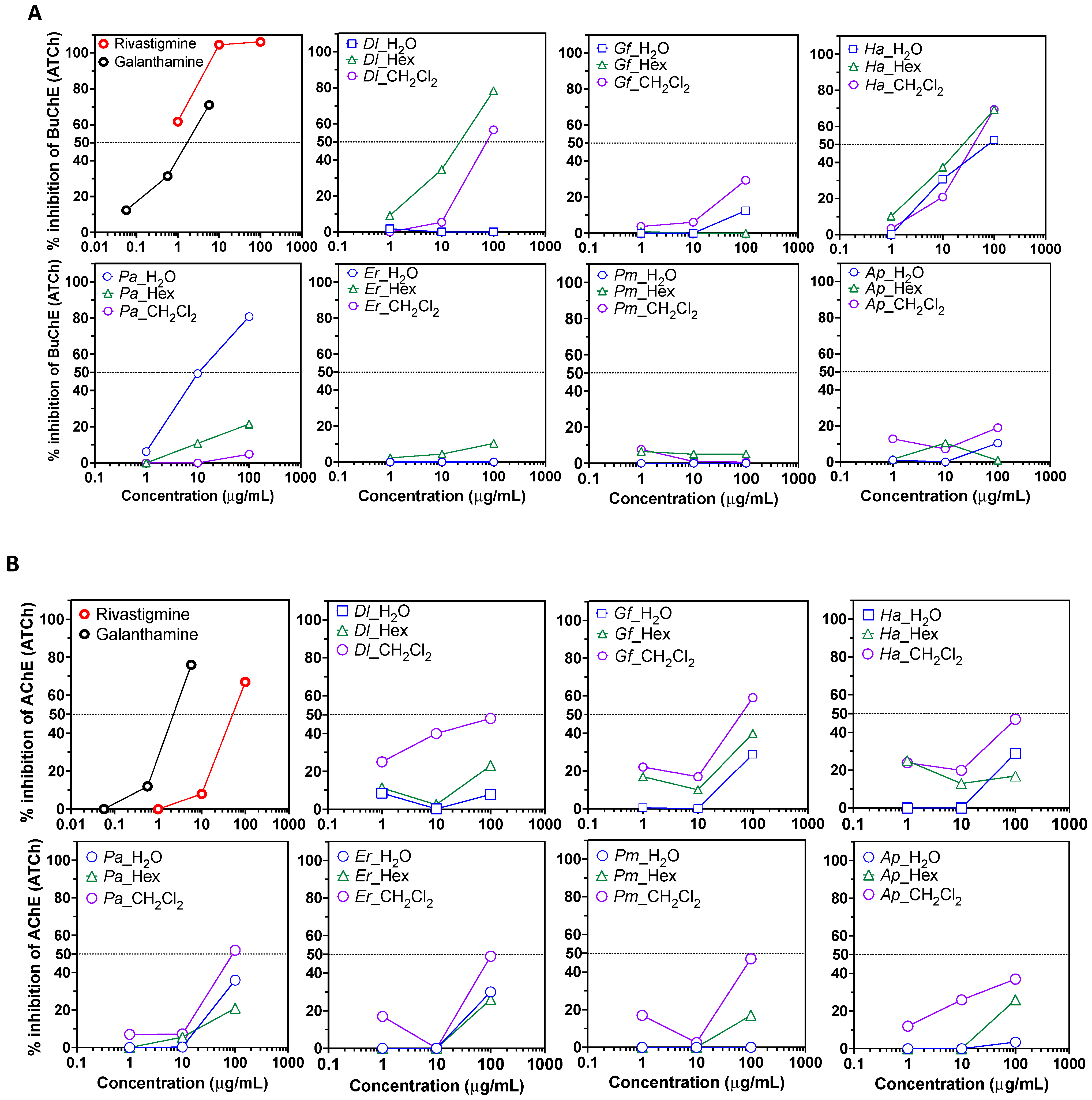
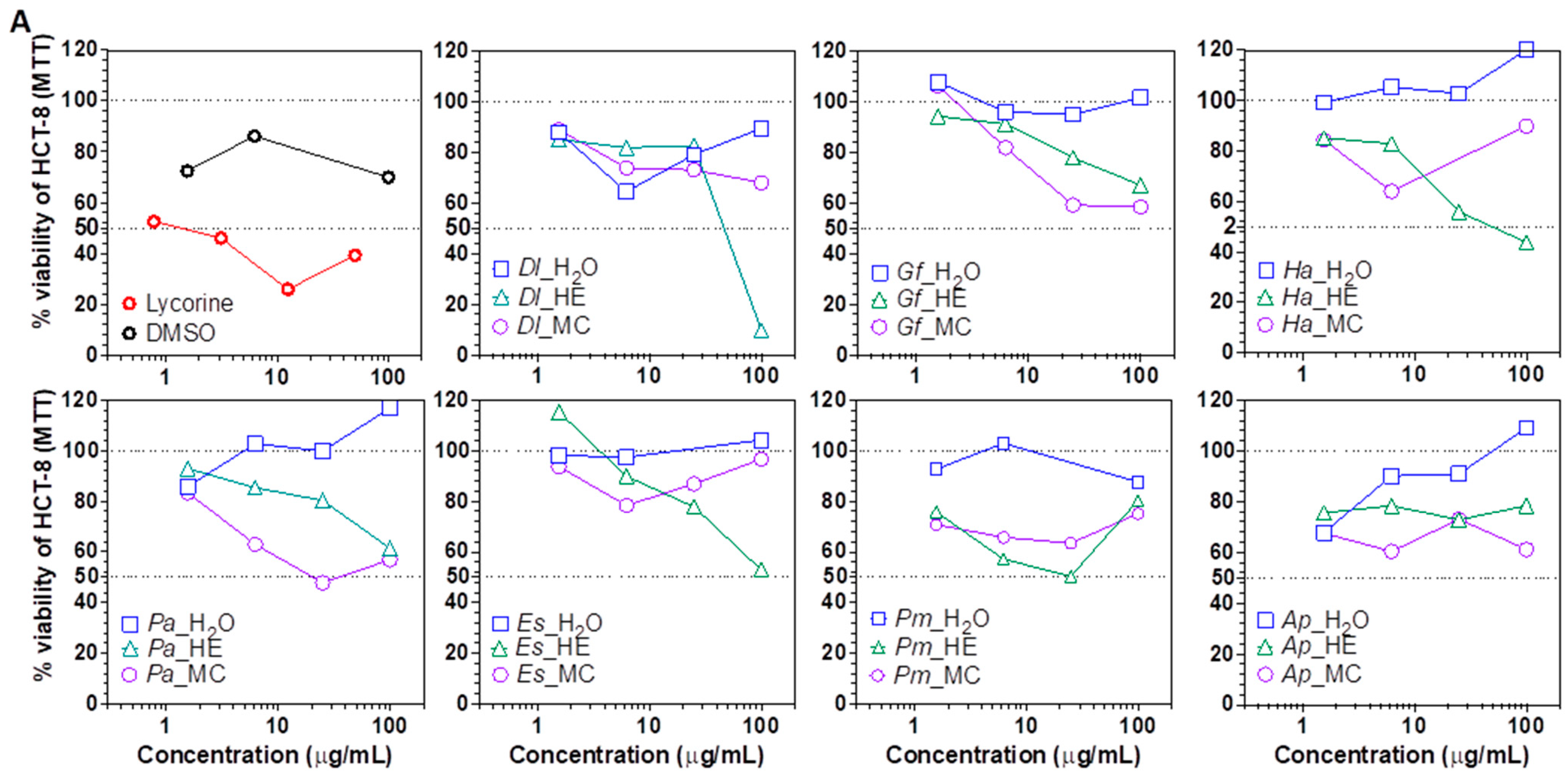
2. Results and Discussion
3. Materials and Methods
3.1. General Procedures
3.2. Plant Material
3.3. General Procedure for the Extraction of Plant Metabolites
3.4. HPLC-MS Analyses of Plant Organic Extracts
3.4.1. Samples Preparation
3.4.2. UHPLC-DAD-MS Analyses
3.4.3. Chemical Reagents
3.5. Biological Assays
3.5.1. Cytotoxic Assays
3.5.2. Antiviral Assays
Dengue Virus Infection
3.5.3. Anti-Cholinesterase Assays
3.5.4. Statistical Analyses
3.5.5. Phytotoxicity Bioassays
3.5.6. Bioassays on Tomato Seeds
3.5.7. Bioassays on Phelipanche ramosa Seeds
3.5.8. Bioassays on Cress Seeds
3.5.9. Leaf Puncture Assay
3.5.10. Antifungal Bioassays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2000, 83, 770–803. [Google Scholar] [CrossRef] [Green Version]
- Tringali, C. Bioactive Compounds from Natural Sources: Isolation, Characterization and Biological Properties, 1st ed.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 2nd ed; Wiley: Chichester, UK, 2002. [Google Scholar]
- Osbourn, A.E.; Lanzotti, V. Plant-Derived Natural Products; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Ortiz, A.; Sansinenea, E. Recent advancements for microorganisms and their natural compounds useful in agriculture. Appl. Microbiol. Biotechnol. 2021, 105, 891–897. [Google Scholar] [CrossRef]
- Gosavi, S.; Subramanian, M.; Reddy, R.; Shet, B.L. A study of prescription pattern of neutraceuticals, knowledge of the patients and cost in a tertiary care hospital. J. Clin. Diagn. Res. 2016, 10, FC01. [Google Scholar] [CrossRef]
- Masi, M.; Di Lecce, R.; Cimmino, A.; Evidente, A. Advances in the chemical and biological characterization of Amaryllidaceae alkaloids and natural analogues isolated in the last decade. Molecules 2020, 25, 5621. [Google Scholar] [CrossRef]
- Evidente, A.; Masi, M. Natural bioactive cinnamoyltyramine alkylamides and co-metabolites. Biomolecules 2021, 11, 1765. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Roberto, L.; Catania, M.R.; Chiaviello, A.; De Natale, A.; Roscetto, E.; Pinto, G.; Pollio, A.; Ingenito, A.; Palumbo, G. Screening and scoring of antimicrobial and biological activities of italian vulnerary plants against major oral pathogenic bacteria. Evid.-Based Complement. Altern. Med. 2013, 316, 280. [Google Scholar] [CrossRef] [Green Version]
- Schrader, K.K.; Avolio, F.; Andolfi, A.; Cimmino, A.; Evidente, A. Ungeremine and its hemisynthesized analogues as bactericides against Flavobacterium columnare. J. Agric. Food Chem. 2013, 61, 1179–1183. [Google Scholar] [CrossRef] [Green Version]
- Oo, O.; Pa, S.; Ps, F. Antimicrobial and antiprotozoal activities of twenty-four Nigerian medicinal plant extracts. S. Afr. J. Bot. 2018, 117, 240–246. [Google Scholar]
- Pollio, A.; Zarrelli, A.; Romanucci, V.; Di Mauro, A.; Barra, F.; Pinto, G.; Crescenzi, E.; Romulo, A.; Ea, Z.; Rondevaldova, J.; et al. Screening of in vitro antimicrobial activity of plants used in traditional Indonesian medicine. Pharm. Biol. 2018, 56, 287–293. [Google Scholar]
- Ribeiro, I.C.D.O.; Mariano, E.G.A.; Careli, R.T.; Morais-Costa, F.; De Sant’Anna, F.M.; Pinto, M.S.; De Souza, M.R.; Duarte, E.R. Plants of the Cerrado with antimicrobial effects against Staphylococcus spp. and Escherichia coli from cattle. BMC Vet. Res. 2018, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Valerio, F.; Masi, M.; Cimmino, A.; Moeini, S.A.; Lavermicocca, P.; Evidente, A. Antimould microbial and plant metabolites with potential use in intelligent food packaging. Nat. Prod. Res. 2018, 32, 1605–1610. [Google Scholar] [CrossRef]
- Avolio, F.; Rimando, A.M.; Cimmino, A.; Andolfi, A.; Jain, S.; Tekwani, B.L.; Evidente, A. Inuloxins A-D and derivatives as antileishmanial agents: Structure-activity relationship study. J. Antibiot. 2014, 67, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Cimmino, A.; Tabanca, N.; Becnel, J.J.; Bloomquist, J.R.; Evidente, A. A survey of bacterial, fungal and plant metabolites against Aedes aegypti (Diptera: Culicidae), the vector of yellow and dengue fevers and Zika virus. Open Chem. 2017, 15, 156–166. [Google Scholar] [CrossRef]
- Masi, M.; Di Lecce, R.; Mérindol, N.; Girard, M.P.; Berthoux, L.; Desgagné-Penix, I.; Calabrò, V.; Evidente, A. Cytotoxicity and antiviral properties of alkaloids isolated from Pancratium maritimum. Toxins 2022, 14, 262. [Google Scholar] [CrossRef] [PubMed]
- Gueribis, F.; Zermane, N.; Khalfi-Habess, O.; Siafa, A.; Cimmino, A.; Boari, A.; Evidente, A. Bioefficacy of compounds from Dittrichia viscosa (Asteraceae) as protectant of chickpea seeds against the cowpea seed beetle Callosobruchus maculatus (Coleoptera: Chrysomelidae). J. Plant Dis. Prot. 2019, 126, 437–446. [Google Scholar] [CrossRef]
- Freda, F.; Masi, M.; Kashefi, J.; Cristofaro, M.; Musmeci, S.; Evidente, A. Acaricidal activity of the plant sesquiterpenoids α-costic acid and inuloxin A against the cattle ectoparasitic tick, Rhipicephalus (Boophilus) annulatus. Int. J. Acarol. 2020, 46, 409–413. [Google Scholar] [CrossRef]
- Cimmino, A.; Freda, F.; Santoro, E.; Superchi, S.; Evidente, A.; Cristofaro, M.; Masi, M. α-Costic acid, a plant sesquiterpene with acaricidal activity against Varroa destructor parasitizing the honey bee. Nat. Prod. Res. 2021, 35, 1428–1435. [Google Scholar] [CrossRef]
- Ganassi, S.; Masi, M.; Grazioso, P.; Evidente, A.; De Cristofaro, A. Activity of some plant and fungal metabolites towards Aedes albopictus (Diptera, Culicidae). Toxins 2021, 13, 285. [Google Scholar] [CrossRef]
- Shi, B.B.; Kongkiatpaiboon, S.; Chen, G.; Schinnerl, J.; Cai, X.H. Nematocidal alkaloids from the roots of Stemona mairei (H. Lév.) K. Krause and identification of their pharmacophoric moiety. Bioorg. Chem. 2022, 14, 106239. [Google Scholar] [CrossRef]
- Lamoral-Theys, D.; Decaestecker, C.; Mathieu, V.; Dubois, J.; Kornienko, A.; Kiss, R.; Pottier, L. Lycorine and its derivatives for anticancer drug design. Mini-Rev. Med. Chem. 2010, 10, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Van Goietsenoven, G.; Hutton, J.; Becker, J.P.; Lallemand, B.; Robert, F.; Lefranc, F.; Pirker, C.; Vandebussche, G.; Van Antwerpen, P.; Evidente, A.; et al. Targeting of eEF1A with Amaryllidaceae isocarbostyrils as a strategy to combat melanomas. FASEB J. 2010, 24, 4575–4584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cimmino, A.; Mathieu, V.; Evidente, M.; Ferderin, M.; Banuls, L.M.Y.; Masi, M.; De Carvalho, A.; Kiss, R.; Evidente, A. Glanduliferins A and B, two new glucosylated steroids from Impatiens glandulifera, with in vitro growth inhibitory activity in human cancer cells. Fitoterapia 2016, 109, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Gunawardana, S.; van Rensburg, M.J.; James, P.C.; Mochel, J.G.; Heliso, P.S.; Albalawib, A.S.; Cimmino, A.; van Otterloc, W.A.L.; Kornienko, A.; et al. Alkaloids isolated from Haemanthus humilis Jacq., an indigenous South African Amaryllidaceae: Anticancer activity of coccinine and montanine. S. Afr. J. Bot. 2019, 126, 277–281. [Google Scholar] [CrossRef]
- Ka, S.; Masi, M.; Merindol, N.; Di Lecce, R.; Plourde, M.B.; Seck, M.; Górecki, M.; Pescitelli, G.; Desgagne-Penix, I.; Evidente, A. Gigantelline, gigantellinine and gigancrinine, cherylline-and crinine-type alkaloids isolated from Crinum jagus with anti-acetylcholinesterase activity. Phytochemistry 2020, 175, 112390. [Google Scholar] [CrossRef] [PubMed]
- Karakoyun, Ç.; Bozkurt, B.; Çoban, G.; Masi, M.; Cimmino, A.; Evidente, A.; Somer, N.U. A comprehensive study on Narcissus tazetta subsp. tazetta L.: Chemo-profiling, isolation, anticholinesterase activity and molecular docking of Amaryllidaceae alkaloids. S. Afr. J. Bot. 2020, 130, 148–154. [Google Scholar]
- Andolfi, A.; Zermane, N.; Cimmino, A.; Avolio, F.; Boari, A.; Vurro, M.; Evidente, A. Inuloxins A-D, phytotoxic bi-and tri-cyclic sesquiterpene lactones produced by Inula viscosa: Potential for broomrapes and field dodder management. Phytochemistry 2013, 86, 112–120. [Google Scholar] [CrossRef]
- Sangermano, F.; Masi, M.; Kumar, A.; Peravali, R.; Tuzi, A.; Cimmino, A.; Vallone, D.; Giamundo, G.; Conte, I.; Evidente, A.; et al. In vitro and in vivo toxicity evaluation of natural products with potential applications as biopesticides. Toxins 2021, 13, 805. [Google Scholar] [CrossRef]
- Moeini, A.; Masi, M.; Zonno, M.C.; Boari, A.; Cimmino, A.; Tarallo, O.; Vuro, M.; Evidente, A. Encapsulation of inuloxin A, a plant germacrane sesquiterpene with potential herbicidal activity, in β-cyclodextrins. Org. Biomol. Chem. 2019, 17, 2508–2515. [Google Scholar] [CrossRef]
- Serino, N.; Boari, A.; Santagata, G.; Masi, M.; Malinconico, M.; Evidente, A.; Vurro, M. Biodegradable polymers as carriers for tuning the release and improve the herbicidal effectiveness of Dittrichia viscosa plant organic extracts. Pest Manag. Sci. 2021, 77, 646–658. [Google Scholar] [CrossRef]
- Ferrazzano, G.F.; Cantile, T.; Roberto, L.; Ingenito, A.; Catania, M.R.; Roscetto, E.; Palumbo, G.; Zarrelli, A.; Pollio, A. Determination of the in vitro and in vivo antimicrobial activity on salivary Streptococci and Lactobacilli and chemical characterisation of the phenolic content of a Plantago lanceolata infusion. BioMed Res. Int. 2015, 2015, 286817. [Google Scholar] [CrossRef] [Green Version]
- Bocquet, L.; Sahpaz, S.; Bonneau, N.; Beaufay, C.; Mahieux, S.; Samaillie, J.; Roumy, V.; Jacquin, J.; Bordage, S.; Hennebelle, T.; et al. Phenolic compounds from Humulus lupulus as natural antimicrobial products: New weapons in the fight against methicillin resistant Staphylococcus aureus, Leishmania mexicana and Trypanosoma brucei strains. Molecules 2019, 24, 1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naz, R.; Roberts, T.H.; Bano, A.; Nosheen, A.; Yasmin, H.; Hassan, M.N.; Keyani, R.; Ullah, S.; Khan, W.; Anwar, Z. GC-MS analysis, antimicrobial, antioxidant, antilipoxygenase and cytotoxic activities of Jacaranda mimosifolia methanol leaf extracts and fractions. PLoS ONE 2020, 15, e0236319. [Google Scholar] [CrossRef] [PubMed]
- Napoli, E.; Siracusa, L.; Ruberto, G.; Carrubba, A.; Lazzara, S.; Speciale, A.; Cimino, F.; Saija, A.; Cristani, M. Phytochemical profiles, phototoxic and antioxidant properties of eleven Hypericum species–A comparative study. Phytochemistry 2018, 152, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Chaouche, T.M.; Haddouchi, F.; Ksouri, R.; Medini, F.; El-Haci, I.A.; Boucherit, Z.; SEkkal, F.Z.; Atik-Bekara, F. Antioxidant potential of hydro-methanolic extract of Prasium majus L: An in vitro study. Pak. J. Biol. Sci. 2013, 16, 1318–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammami, S.; Jannet, H.B.; Mighri, Z.; Nuzillard, J.M. Iridoid and flavonoid glycosides from the aerial part of Prasium majus growing in Tunisia. J. Soc. Chem. Tunisie 2003, 5, 17–24. [Google Scholar]
- Basta, A.; Tzakou, O.; Couladis, M.; Yannitsaros, A. Essential oil composition of Prasium majus L. from Greece. Flavour Fragr. J. 2007, 22, 347–349. [Google Scholar] [CrossRef]
- Kabbash, A.; Shoeib, N. Chemical and biological investigation of some secondary metabolites in Atriplex halimus growing in Egypt. Nat. Prod. Commun. 2012, 7, 1465–1468. [Google Scholar] [CrossRef] [Green Version]
- Nejma, A.B.; Nguir, A.; Jannet, H.B.; Daïch, A.; Othman, M.; Lawson, A.M. New septanoside and 20-hydroxyecdysone septanoside derivative from Atriplex halimus roots with preliminary biological activities. Bioorg. Med. Chem. Lett. 2015, 25, 1665–1670. [Google Scholar] [CrossRef]
- Dey, S.; Samanta, G.C.; Jayaraman, N. Advancements in synthetic and structural studies of septanoside sugars. Recent Trends Carbohydr. Chem. 2020, 1, 217–251. [Google Scholar]
- Dghim, F.; Zouari, F.; Boukhris, M.; Neffati, M. Composition chimique et activite antioxydante d’un arbuste des zones arides: Periploca angustifolia labill (apocynacees). J. Soc. Chim. Tunisie 2013, 15, 163–173. [Google Scholar]
- Dghim, F.; Zouari, F.; Boukhris, M.; Neffati, M. Chemical composition of root bark of Periploca angustifolia growing wild in Saharian Tunisia. J. Essent. Oil Bear. Plants 2013, 16, 338–345. [Google Scholar]
- Athmouni, K.; Belhaj, D.; Mkadmini Hammi, K.; El Feki, A.; Ayadi, H. Phenolic compounds analysis, antioxidant, and hepatoprotective effects of Periploca angustifolia extract on cadmium-induced oxidative damage in HepG2 cell line and rats. Arch. Physiol. Biochem. 2018, 124, 261–274. [Google Scholar] [CrossRef]
- Athmouni, K.; Belhaj, D.; Ayadi, H.; Chawech, R.; Jarraya, R.; El Feki, A. Characterization of polysaccharides isolated from Periploca angustifolia and its antioxidant activity and renoprotective potential against cadmium induced toxicity in HEK293 cells and rat kidney. Int. J. Biol. Macromol. 2019, 125, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.C.; Yu, J.T.; Wang, H.F.; Tan, M.S.; Meng, X.F.; Wang, C.; Jiang, T.; Zhu, X.C.; Tan, L. Efficacy and safety of donepezil, galantamine, rivastigmine, and memantine for the treatment of Alzheimer’s disease: A systematic review and meta-analysis. J. Alzheimer’s Dis. 2014, 41, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Tao, H.; Mei, J.; Tang, X. The anticancer effects of 2-methoxyestradiol on human huh7 cells in vitro and in vivo. Biochem. Biophys. Res. Commun. 2019, 512, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Wang, S.; Liang, W.; Wang, W.; Wang, H.; Zhao, M.; Liu, X. Salvianolic acid B reverses multidrug resistance in HCT-8/VCR human colorectal cancer cells by increasing ROS levels. Mol. Med. Rep. 2017, 15, 724–730. [Google Scholar] [CrossRef] [Green Version]
- Arafa, A.M.; Mohamed, M.E.; Eldahmy, S.I. The aerial parts of yellow horn poppy (Glaucium flavum Cr.) growing in Egypt: Isoquinoline alkaloids and biological activities. J. Pharm. Sci. Res. 2016, 8, 323. [Google Scholar]
- Wu, F.; Zhao, S.; Yu, B.; Chen, Y.M.; Wang, W.; Song, Z.G.; Hu, Y.; Tao, Z.W.; Tian, J.H.; Pei, Y.Y.; et al. A new coronavirus associated with human respiratory disease in China. Nature 2020, 578, 265–269. [Google Scholar] [CrossRef]
- Chala, B.; Hamde, F. Emerging and re-emerging vector-borne infectious diseases and the challenges for control: A review. Public Health Front. 2021, 9, 715759. [Google Scholar] [CrossRef]
- Zou, G.; Puig-Basagoiti, F.; Zhang, B.; Qing, M.; Chen, L.; Pankiewicz, K.W.; Felczak, K.; Yuan, Z.; Shi, P.Y. A single-amino acid substitution in West Nile virus 2K peptide between NS4A and NS4B confers resistance to lycorine, a flavivirus inhibitor. Virology 2009, 384, 242–252. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Niu, J.; Wang, C.; Huang, B.; Wang, W.; Zhu, N.; Deng, Y.; Wang, H.; Ye, F.; Cen, S.; et al. High-Throughput screening and identification of potent broad-spectrum inhibitors of coronaviruses. J. Virol. 2019, 93, e00023-19. [Google Scholar] [CrossRef] [Green Version]
- Stojanovic, G.; Ethordevic, A.; Smelcerovic, A. Do other Hypericum species have medical potential as St. John’s wort (Hypericum perforatum)? Curr. Med. Chem. 2013, 20, 2273–2295. [Google Scholar] [CrossRef] [PubMed]
- Zanella, L.; Vianello, F. Functional food from endangered ecosystems: Atriplex halimus as a case study. Foods 2020, 9, 1533. [Google Scholar] [CrossRef]
- Matos, A.D.R.; Caetano, B.C.; de Almeida Filho, J.L.; Martins, J.; de Oliveira, M.G.P.; Sousa, T.D.C.; Horta, M.A.P.; Siqueira, M.M.; Fernandez, J.H. Identification of hypericin as a candidate repurposed therapeutic agent for cOVID-19 and its potential anti-SARS-CoV-2 activity. Front. Microbiol. 2022, 13, 828984. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, F.F.; Anhlan, D.; Schofbanker, M.; Schreiber, A.; Classen, N.; Hensel, A.; Hempel, G.; Scholz, W.; Kuhn, J.; Hrincius, E.R.; et al. Hypericum perforatum and its ingredients hypericin and pseudohypericin demonstrate an antiviral activity against SARS-CoV-2. Pharmaceuticals 2022, 15, 530. [Google Scholar] [CrossRef]
- Sommier, S. Le isole Pelagie—Lampedusa, Linosa e Lampione—E la Loro Flora; Stabilimento Pellas, Luigi Chiti Successore: Firenze, Italy, 1908. [Google Scholar]
- Di Martino, A. Flora and vegetation. In Biogeography of the Pelagie Islands; Zavattari, E., Ed.; Reports of the National Academy of the XL: Rome, Italy, 1958; Series IV; Volume XI, pp. 163–261. [Google Scholar]
- Corti, C.; Lo Cascio, P.; Massetti, M.; Corti, C.; Pasta, S. Storia naturale delle Isole Pelagie; Società Editrice L’Epos: Palermo, Italy, 2002; pp. 151–192. [Google Scholar]
- Lamoral-Theys, D.; Andolfi, A.; Van Goietsenoven, G.; Cimmino, A.; Le Calve, B.; Wauthoz, N.; Megalizzi, V.; Gras, T.; Bruyere, C.; Dubois, J.; et al. Lycorine, the main phenanthridine Amaryllidaceae alkaloid, exhibits significant antitumor activity in cancer cells that display resistance to proapoptotic stimuli: An investigation of structure-activity relationship and mechanistic insight. J. Med. Chem. 2009, 52, 6244–6256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischl, W.; Bartenschlager, R. High-throughput screening using dengue virus reporter genomes. Methods Mol. Biol. 2013, 1030, 205–219. [Google Scholar]
- Ka, S.; Merindol, N.; Sow, A.A.; Singh, A.; Landelouci, K.; Plourde, M.B.; Pépin, G.; Masi, M.; Di Lecce, R.; Evidente, A.; et al. Amaryllidaceae alkaloid cherylline inhibits the replication of Dengue and Zika viruses. Antimicrob. Agents Chemother. 2021, 65, e0039821. [Google Scholar] [CrossRef] [PubMed]
- Chatel-Chaix, L.; Bartenschlager, R. Dengue virus- and hepatitis C virus-induced replication and assembly compartments: The enemy inside—Caught in the web. J. Virol. 2014, 88, 5907–5911. [Google Scholar] [PubMed]
- Gabrielsen, B.; Monath, T.P.; Huggins, J.W.; Kefauver, D.F.; Pettit, G.R.; Groszek, G.; Hollingshead, M.; Kirsi, J.J.; Shannon, W.M.; Schubert, E.M.; et al. Antiviral (RNA) activity of selected Amaryllidaceae isoquinoline constituents and synthesis of related substances. J. Nat. Produt. 1992, 55, 1569–1581. [Google Scholar] [CrossRef]
- Summa, V.; Petrocchi, A.; Bonelli, F.; Crescenzi, B.; Donghi, M.; Ferrara, M.; Fiore, F.; Gardelli, C.; Gonzalez Paz, O.; Hazuda, D.J.; et al. Discovery of raltegravir, a potent, selective orally bioavailable HIV-integrase inhibitor for the treatment of HIV-AIDS infection. J. Med. Chem. 2008, 51, 5843–5855. [Google Scholar] [CrossRef] [PubMed]
- Mamun, A.A.; Pidany, F.; Hulcova, D.; Marikova, J.; Kucera, T.; Schmidt, M.; Catapano, M.C.; Hrabinova, M.; Jun, D.; Muckova, L.; et al. Amaryllidaceae alkaloids of norbelladine-type as inspiration for development of highly selective butyrylcholinesterase inhibitors: Synthesis, biological activity evaluation, and docking studies. Int. J. Molecul. Sci. 2021, 22, 8308. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Boari, A.; Evidente, A.; Vurro, M. Metabolites inhibiting germination of Orobanche ramosa seeds produced by Myrothecium verrucaria and Fusarium compactum. J. Agric. Food Chem. 2005, 53, 1598–1603. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Botanical Family | Weight (mg) of 1 HE Extract | Weight (mg) of MC Extract | N articles Found | N Articles Found with “Metabolite” as Keyword |
|---|---|---|---|---|---|
| Atriplex halimus (Ap) | Amaranthaceae | 14.74 | 40.68 | 401 | 4 |
| Daucus lopadusanus (Dl) | Apiaceae | 153.20 | 84.50 | 1 | 0 |
| Echinops spinosus (Es) | Asteraceae | 26.72 | 119.63 | 40 | 0 |
| Glaucium flavum (Gf) | Papaveraceae | 203.57 | 2765.62 | 221 | 0 |
| Hypericum aegypticum (Ha) | Hypericaceae | 77.92 | 173.96 | 9 | 0 |
| Periploca angustifolia (Pa) | Apocynaceae | 12.96 | 89.28 | 22 | 0 |
| Prasium majus (Pm) | Lamiaceae | 71.25 | 264.23 | 28 | 3 |
| Tomato | Cress | Broomrape | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rootlet Length (cm) | Germinated Seeds (n.) | Rootlet Length (cm) | Germinated Seeds (%) | |||||||||
| WR 1 | MC | HE | WR | MC | HE | WR | MC | HE | WR | MC | HE | |
| Daucus lopadusanus | 1.8 | 2.0 | 3.1 | 7 | 4.5 | 8.5 | 1.30 | 1.06 | 1.42 | 78.5 | 56 | 46.5 |
| Glaucium flavum | 2.4 | 2.4 | 2.8 | 8 | 6.5 | 8 | 1.10 | 1.21 | 1.10 | 67.5 | 37 | 68 |
| Hypericum aegypticum | 2.9 | 1.6 | 3.2 | 7.5 | 6.5 | 5 | 2.03 | 1.25 | 1.13 | 80 | 44.5 | 59 |
| Periploca angustifolia | 2.5 | 1.3 | 2.0 | 8 | 5 | 6 | 0.88 | 1.03 | 0.94 | 65 | 44 | 59.5 |
| Echinops ritro | 2.4 | 0.6 | 2.8 | 5.5 | 1 | 9.5 | 0.58 | 0.35 | 1.07 | 63.5 | 34 | 53 |
| Prasium majus | 2.4 | 0.9 | 3.0 | 7.5 | 7.5 | 5.5 | 1.55 | 0.82 | 1.15 | 73 | 47 | 66 |
| Atriplex halimus | 1.6 | 1.1 | 2.5 | 7 | 3 | 6.5 | 0.79 | 0.74 | 1.13 | 52.5 | 54 | 64.5 |
| Control | 3.0 | 7.5 | 2.56 | 70 | ||||||||
| CA 1 | CT | HE | SN | SO | SH | PO | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WR 2 | MC | HE | WR | MC | HE | WR | MC | HE | WR | MC | HE | WR | MC | HE | WR | MC | HE | WR | MC | HE | |
| Daucus lopadusanus | - 3 | + | + | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | + | + |
| Glaucium flavum | - | - | + | - | - | - | - | + | - | - | - | - | - | + | - | - | + | + | - | + | + |
| Hypericum aegypticum | - | + | - | - | - | - | - | - | - | - | - | - | - | + | + | - | + | + | - | + | + |
| Periploca angustifolia | - | + | - | - | - | - | + | - | - | - | - | - | - | - | - | - | - | - | - | + | + |
| Echinops ritro | - | + | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | + |
| Prasium majus | - | - | - | - | - | - | - | - | - | - | - | - | - | - | + | - | - | + | - | - | + |
| Atriplex halimus | - | - | + | - | - | - | - | + | - | - | - | - | - | - | + | - | - | - | - | - | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Lecce, R.; Mérindol, N.; Pérez, M.G.; Karimzadegan, V.; Berthoux, L.; Boari, A.; Zidorn, C.; Vurro, M.; Surico, G.; Desgagné-Penix, I.; et al. Biochemical Analyses of Bioactive Extracts from Plants Native to Lampedusa, Sicily Minor Island. Plants 2022, 11, 3447. https://doi.org/10.3390/plants11243447
Di Lecce R, Mérindol N, Pérez MG, Karimzadegan V, Berthoux L, Boari A, Zidorn C, Vurro M, Surico G, Desgagné-Penix I, et al. Biochemical Analyses of Bioactive Extracts from Plants Native to Lampedusa, Sicily Minor Island. Plants. 2022; 11(24):3447. https://doi.org/10.3390/plants11243447
Chicago/Turabian StyleDi Lecce, Roberta, Natacha Mérindol, Mayra Galarza Pérez, Vahid Karimzadegan, Lionel Berthoux, Angela Boari, Christian Zidorn, Maurizio Vurro, Giuseppe Surico, Isabel Desgagné-Penix, and et al. 2022. "Biochemical Analyses of Bioactive Extracts from Plants Native to Lampedusa, Sicily Minor Island" Plants 11, no. 24: 3447. https://doi.org/10.3390/plants11243447