
Phytochemicals and Their Correlation with Molecular Data in Micromeria and Clinopodium (Lamiaceae) Taxa

 , , , , , , , and
, , , , , , , and
Abstract
:1. Introduction
2. Results
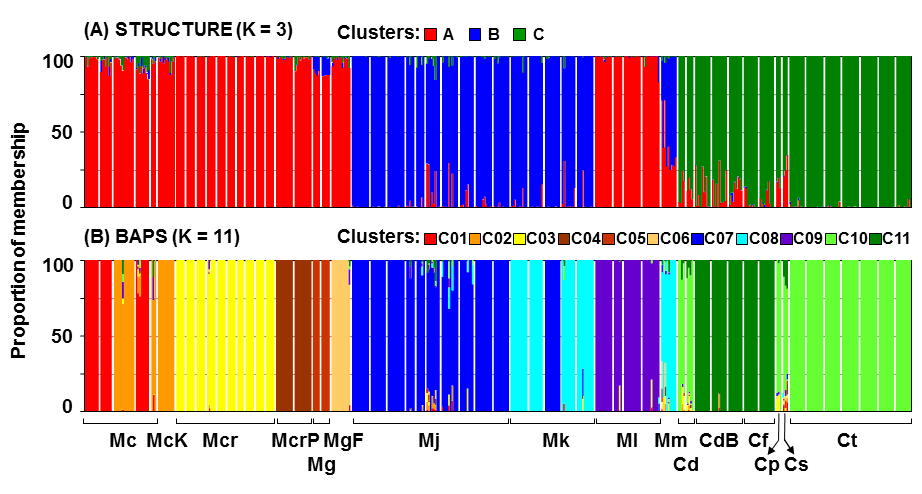
2.1. AFLP Analysis
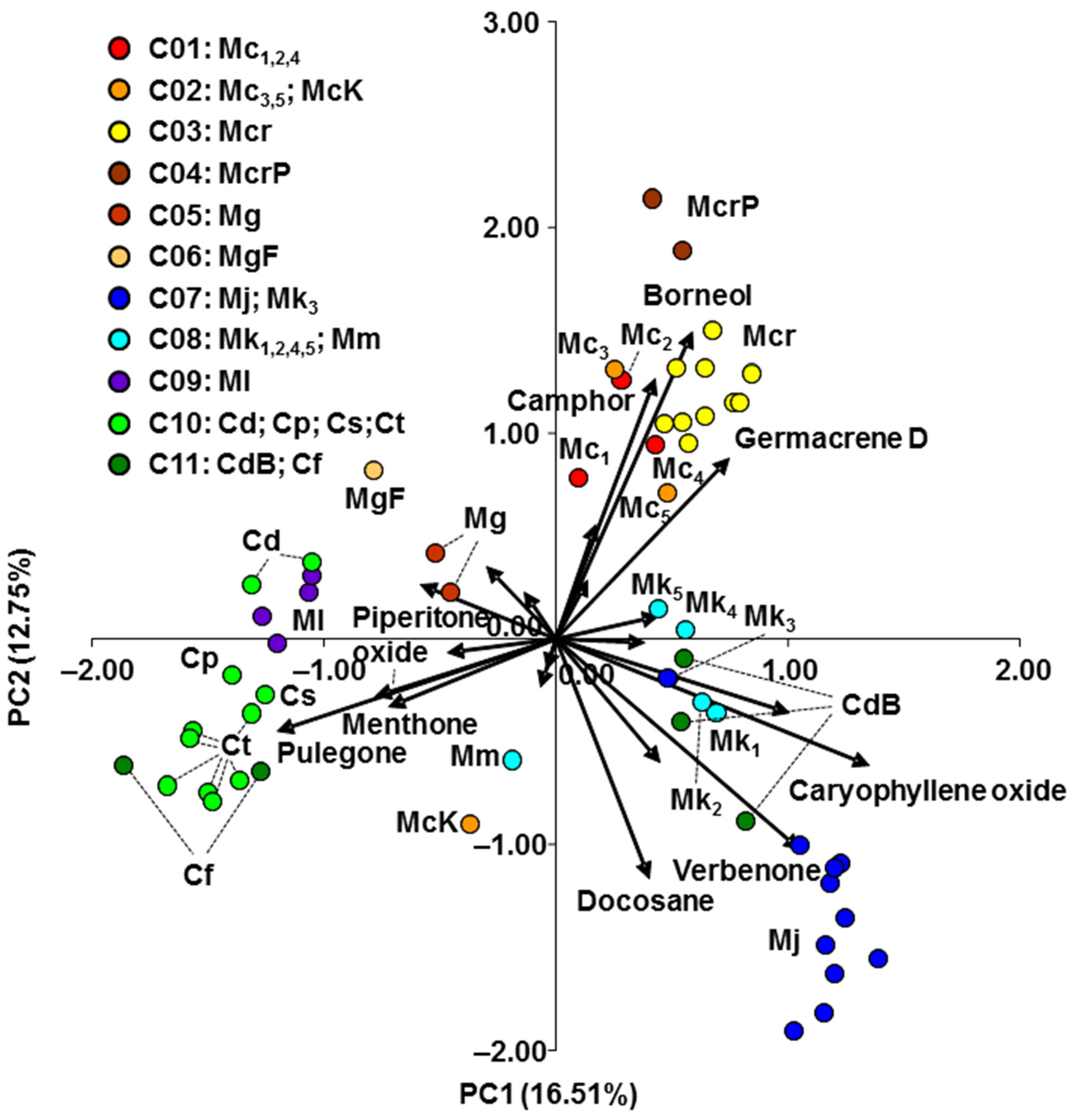
2.2. Essential Oil Analysis
2.3. Mantel Test
3. Discussion
4. Materials and Methods
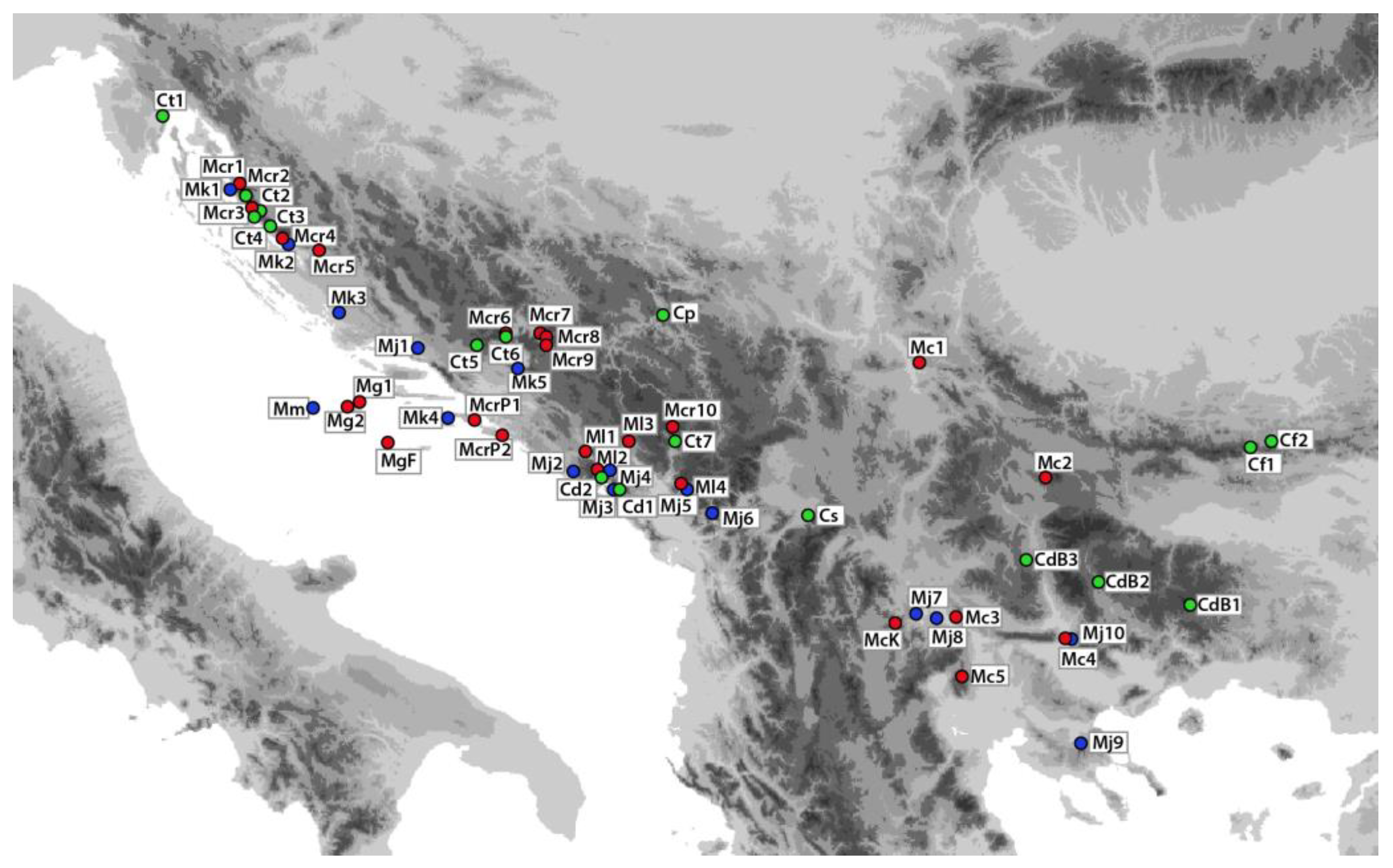
4.1. Plant Material
4.2. Molecular Analysis
4.2.1. DNA Isolation
4.2.2. AFLP Data Analysis
Within-Population Diversity
Population Differentiation and Structure
4.3. Gas Chromatography and Mass Spectrometry (GC-MS) Analyses
4.4. Principal Component Analysis
4.5. Mantel Test
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bräuchler, C.; Ryding, O.; Heubl, G. The genus Micromeria (Lamiaceae), a synoptical update. Willdenowia 2008, 38, 363–410. [Google Scholar] [CrossRef] [Green Version]
- Erhardt, W.; Götz, E.; Bödeker, N.; Seybold, S. Zander—Handwörterbuch der Pflanzennamen, 19th ed.; Eugen Ulmer GmbH und Co.: Stuttgart, Germany, 2014; p. 576. [Google Scholar]
- Bräuchler, C.; Meimberg, H.; Abele, T.; Heubl, G. Polyphyly of the genus Micromeria (Lamiaceae)—evidence from cpDNA sequence data. Taxon 2005, 54, 639–650. [Google Scholar] [CrossRef]
- Chater, A.O.; Guinea, E. Micromeria Benth. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1972; Volume 3, pp. 167–170. [Google Scholar]
- Briquet, J. Labiatae. In Die natürlichen Pflanzenfamilien; Engler, A., Prantl, K., Eds.; Wilhelm Engelmann: Leipzig, Germany, 1897; Volume 4, pp. 183–380. [Google Scholar]
- Killick, D.J.B. South African species of Satureia. Bothalia 1961, 7, 435–437. [Google Scholar] [CrossRef] [Green Version]
- Greuter, W.; Burdet, H.M.; Long, D. Satureja. In Med-Checklist; Conservatoire et Jardin Botaniques de la ville de Genève: Geneva, Switzerland, 1986; Volume 3, pp. 323–341. [Google Scholar]
- Bentham, G. Labiatae. In Prodromus Systematis Universalis Regni Vegetabilis; Candolle, A.P., Ed.; Treuttel et Würtz: Paris, France, 1848; Volume 12, pp. 212–226. [Google Scholar]
- Pérez de Paz, P.L. Revisión del Género Micromeria Bentham (Lamiaceae—Stachyoideae) en la Región Macaronésica; Instituto de Estudios Canarios: La Laguna de Tenerife, Spain, 1978; 306p. [Google Scholar]
- Doroszenko, A. Taxonomic Studies on the Satureja Complex (Labiatae). Ph.D. Thesis, Edinburgh University, Edinburgh, UK, 1986. [Google Scholar]
- Valverde, R.M. Sinopsis y distribución del género Micromeria Bentham. Bot. Complut. 1993, 18, 157–168. [Google Scholar]
- Morales, R.; Luque, M.N. El género Calamintha Mill. (Labiatae) en la Península Ibérica e Islas Baleares. Anal. Jard. Bot. Madr. 1997, 55, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Harley, R.M.; Atkins, S.; Budantzev, A.L.; Cantino, P.D.; Conn, B.J.; Grayer, R.; Harley, M.M.; De Kok, R.; Krestovskaja, T.; Morales, R.; et al. Labiatae. In The Families and Genera of Vascular Plants, 1st ed.; Kubitzki, K., Ed.; Springer: Berlin, Germany, 2004; Volume 7, pp. 167–282. [Google Scholar]
- Boissier, E. Flora Orientalis; Apud H. Georg, Biblioplam: Genevae, Switzerland, 1879; Volume 4, pp. 568–575. [Google Scholar]
- Bräuchler, C.; Meimberg, H.; Heubl, G. New names in Old World Clinopodium—The transfer of the species of Micromeria sect. Pseudomelissa to Clinopodium. Taxon 2006, 55, 977–981. [Google Scholar] [CrossRef]
- Kremer, D.; Stabentheiner, E.; Bogunić, F.; Ballian, D.; Eleftheriadou, E.; Stešević, D.; Matevski, V.; Ranđelović, V.; Ivanova, D.; Ruščić, M.; et al. Micromorphological traits of Balcanic Micromeria and closely related Clinopodium species (Lamiaceae). Plants 2021, 10, 1666. [Google Scholar] [CrossRef] [PubMed]
- Diklić, N. Micromeria Bentham. In Flora SR Srbije; Josifović, M., Ed.; Academie Serbe des Sciences et des Arts: Belgrade, Serbia, 1974; Volume 6, pp. 458–462. (In Serbian) [Google Scholar]
- Šilić, Č. Monography of genera Satureja L., Calamintha Miller, Micromeria Bentham, Acinos Miller and Clinopodium L. In Flora of Yugoslavia; Zemaljski Muzej BiH: Sarajevo, Bosnia and Herzegovina, 1979; pp. 172–262. (In Bosnian) [Google Scholar]
- Ančev, M. Micromeria Benth. In Flora Reipublicae Popularis Bulgaricae; Velčev, V., Ed.; Academiae Scientiarum Bulgaricae: Sofia, Bulgaria, 1989; Volume 9, pp. 356–362. [Google Scholar]
- Delipavlov, D.; Cheshmedzhiev, I. Key to the Plants in Bulgaria; Academic Publishing House of Agricultural University: Plovdiv, Bulgaria, 2003. (In Bulgarian) [Google Scholar]
- Petrova, A.; Vladimirov, V. Red list of Bulgarian vascular plants. Phytol. Balc. 2009, 15, 63–94. [Google Scholar]
- Anonymous. Red list of endangered wild species and subspecies of plants, animals and fungi. Off. Gaz. Fed. Bosnia Herzeg. 2014, 7, 111–116. (In Bosnian) [Google Scholar]
- Nikolić, T. Flora Croatica, 1st ed.; Alfa: Zagreb, Croatia, 2019; Volume 4, pp. 408–409. [Google Scholar]
- Šavikin, K.P.; Menković, N.R.; Zdunić, G.M.; Tasić, S.R.; Ristić, M.S.; Stević, T.R.; Dajić-Stevanović, Z.P. Chemical composition and antimicrobial activity of the essential oils of Micromeria thymifolia (Scop.) Fritsch., M. dalmatica Benth., and Satureja cuneifolia Ten. and its secretory elements. J. Essent. Oil Res. 2010, 22, 91–96. [Google Scholar] [CrossRef]
- Vladimir-Knežević, S.; Blažeković, B.; Bival Štefan, M.; Alegro, A.; Köszegi, T.; Petrik, J. Antioxidant activities and polyphenolic contents of three selected Micromeria species from Croatia. Molecules 2011, 16, 1454–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radulović, N.S.; Blagojević, P.D. Volatile secondary metabolites of Micromeria dalmatica Benth. (Lamiaceae): Biosynthetical and chemotaxonomical aspects. Chem. Biodivers. 2012, 9, 1303–1319. [Google Scholar] [CrossRef] [PubMed]
- Marin, M.; Jasnić, N.; Ascensão, L. Histochemical, micromorphology and ultrastructural investigation in glandular trichomes of Micromeria thymifolia. Bot. Serb. 2013, 37, 49–53. [Google Scholar]
- Tošić, S.; Stojičić, D.; Stankov-Jovanović, V.; Mitić, V.; Mihajilov-Krstev, T.; Zlatković, B. Chemical composition, antioxidant and antimicrobial activities of micropropagated and native Micromeria pulegium (Lamiaceae) extracts. Oxid. Commun. 2015, 38, 55–66. [Google Scholar]
- Stojičić, D.; Tošić, S.; Slavkovska, V.; Zlatković, B.; Budimir, S.; Janošević, D.; Uzelac, B. Glandular trichomes and essential oil characteristics of in vitro propagated Micromeria pulegium (Rochel) Benth. (Lamiaceae). Planta 2016, 244, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Mladenova, T.R.; Stoyanov, P.S.; Michova-Nankova, I.K.; Mladenov, R.D.; Boyadzhiev, D.T.; Bivolarska, A.V.; Todorov, K.T. Comparative leaf epidermis analyses of Micromeria frivaldszkyana (Degen) Velen. and Clinopodium vulgare L. (Lamiaceae) from Bulgarka Nature Park, Bulgaria. Ecol. Balk. 2019, 11, 133–140. [Google Scholar]
- Bardy, K.E.; Schönswetter, P.; Schneeweiss, G.M.; Fischer, M.A.; Albach, D.C. Extensive gene flow blurs species boundaries among Veronica barrelieri, V. orchidea and V. spicata (Plantaginaceae) in southeastern Europe. Taxon 2011, 60, 108–121. [Google Scholar] [CrossRef]
- Caković, D.; Stešević, D.; Schönswetter, P.; Frajman, B. How many taxa? Spatiotemporal evolution and taxonomy of Amphoricarpos (Asteraceae, Carduoideae) on the Balkan Peninsula. Org. Divers. Evol. 2015, 15, 429–445. [Google Scholar] [CrossRef]
- Rešetnik, I.; Temunović, M.; Liber, Z.; Šatovic, Z.; Bogdanović, S. Phylogeography of Campanula fenestrellata s.l. (Campanulaceae) in the northern Adriatic. Plant Syst. Evol. 2020, 306, 42. [Google Scholar] [CrossRef]
- Slavkovska, V.; Couladis, M.; Bojovic, S.; Tzakou, O.; Pavlovic, M.; Lakusic, B.; Jancic, R. Essential oil and its systematic significance in species of Micromeria Bentham from Serbia & Montenegro. Plant Syst. Evol. 2005, 255, 1–15. [Google Scholar] [CrossRef]
- Rodrigues, L.; Póvoa, P.; van den Berg, C.; Figueiredo, A.C.; Moldão, M.; Monteiro, A. Genetic diversity in Mentha cervina based on morphological traits, essential oils profile and ISSRs markers. Biochem. Syst. Ecol. 2013, 51, 50–59. [Google Scholar] [CrossRef]
- Feulner, M.; Pointner, S.; Heuss, L.; Aas, G.; Paule, P.; Dötterl, S. Floral scent and its correlation with AFLP data in Sorbus. Org. Divers. Evol. 2014, 14, 339–348. [Google Scholar] [CrossRef]
- Heywood, V.H. The role of chemistry in plant systematics. Pure Appl. Chem. 1973, 34, 355–376. [Google Scholar] [CrossRef]
- Crawford, D.J. Flavonoid chemistry and angiosperm evolution. Bot. Rev. 1978, 44, 431–456. [Google Scholar] [CrossRef]
- Gershenzon, J.; Mabry, T.J. Secondary metabolites and the higher classification of angiosperms. Nord. J. Bot. 1983, 3, 5–34. [Google Scholar] [CrossRef]
- Judd, W.S.; Campbell, C.S.; Kellogg, E.A.; Stevens, P.F.; Donoghue, M.J. Taxonomic evidence: Structural and biochemical characters. In Plant Systematics: A Phylogenetic Approach, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2002; pp. 55–81. [Google Scholar]
- Zhang, Y.; Deng, T.; Sun, L.; Landis, J.B.; Moore, M.J.; Wang, H.; Wang, Y.; Hao, X.; Chen, J.; Li, S.; et al. Phylogenetic patterns suggest frequent multiple origins of secondary metabolites across the seed-plant ‘tree of life’. Nati. Sci. Rev. 2021, 8, nwaa105. [Google Scholar] [CrossRef] [PubMed]
- Kremer, D.; Dunkić, V.; Ruščić, M.; Matevski, V.; Ballian, D.; Bogunić, F.; Eleftheriadou, E.; Stešević, D.; Kosalec, I.; Bezić, N.; et al. Micromorphological traits and essential oil contents of Micromeria kerneri Murb. and M. juliana (L.) Benth. (Lamiaceae). Phytochemistry 2014, 98, 128–136. [Google Scholar] [CrossRef]
- Kremer, D.; Dunkić, V.; Stešević, D.; Kosalec, I.; Ballian, D.; Bogunić, F.; Bezić, N.; Stabentheiner, E. Micromorphological traits and essential oil of Micromeria longipedunculata Bräuchler (Lamiaceae). Cent. Eur. J. Biol. 2014, 9, 559–568. [Google Scholar] [CrossRef] [Green Version]
- Dunkić, V.; Kremer, D.; Jurišić Grubešić, R.; Vuković Rodríguez, J.; Ballian, D.; Bogunić, F.; Stešević, D.; Kosalec, I.; Bezić, N.; Stabentheiner, E. Micromorphological and phytochemical traits of four Clinopodium L. species (Lamiaceae). S. Afr. J. Bot. 2017, 111, 232–241. [Google Scholar] [CrossRef]
- Vandas, C. Novae Plantae Balcanicae. Mag. Bot. Lapok 1905, 4, 109–113. [Google Scholar]
- Gómez, A.; Lunt, D.H. Refugia Within Refugia: Patterns of Phylogeographic Concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia; Weiss, S., Ferrand, N., Eds.; Springer: Berlin, Germany, 2007; pp. 155–188. [Google Scholar]
- Krystufek, B.; Buzan, E.V.; Hutchinson, W.F.; Hänfling, B. Phylogeography of the rare Balkan endemic Martino’s vole, Dinaromys bogdanovi, reveals strong differentiation within the western Balkan Peninsula. Mol. Ecol. 2007, 16, 1221–1232. [Google Scholar] [CrossRef]
- Ursenbacher, S.; Schweiger, S.; Tomović, L.; Crnobrnja-Isailović, J.; Fumagalli, L.; Mayer, W. Molecular phylogeography of the nose-horned viper (Vipera ammodytes, Linnaeus (1758)): Evidence for high genetic diversity and multiple refugia in the Balkan Peninsula. Mol. Phylogenet. Evol. 2008, 46, 1116–1128. [Google Scholar] [CrossRef]
- Surina, B.; Schönswetter, P.; Schneeweiss, G.M. Quaternary range dynamics of ecologically divergent species (Edraianthus serpyllifolius and E. tenuifolius, Campanulaceae) within the Balkan refugium. J. Biogeogr. 2011, 38, 1381–1393. [Google Scholar] [CrossRef]
- Melito, S.; Sias, A.; Petretto, G.L.; Chessa, M.; Pintore, G.; Porceddu, A. Genetic and metabolite diversity of Sardinian populations of Helichrysum italicum. PLoS ONE 2013, 8, e79043. [Google Scholar] [CrossRef] [Green Version]
- Kostadinova, E.; Alipieva, K.; Stefova, M.; Stafilov, T.; Antonova, D.; Evstatieva, L.J.; Matevski, V.; Kulevanova, S.; Stefkov, G.; Bankova, V. Chemical composition of the essential oils of three Micromeria species growing in Macedonia and Bulgaria. Maced. J. Chem. Chem. Eng. 2007, 26, 3–7. [Google Scholar] [CrossRef]
- Stojanović, G.; Palić, I.; Ursić-Janković, J. Composition and antimicrobial activity of the essential oil of Micromeria cristata and Micromeria juliana. Flavour Fragr. J. 2006, 21, 77–79. [Google Scholar] [CrossRef]
- Çarikçi, S. The essential oil components of five Micromeria species grown in Anatolia. BAÜ Fen Bil. Enst. Derg. Cilt 2013, 15, 73–79. [Google Scholar]
- Kremer, D.; Stabentheiner, E.; Dunkić, V.; Dragojević Müller, I.; Vujić, L.; Kosalec, I.; Ballian, D.; Bogunić, F.; Bezić, N. Micromorphological and chemotaxonomical traits of Micromeria croatica (Pers.) Schott. Chem. Biodivers. 2012, 49, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Vuko, E.; Rusak, G.; Dunkić, V.; Kremer, D.; Kosalec, I.; Rađa, B.; Bezić, N. Inhibition of satellite RNA associated cucumber mosaic virus infection by essential oil of Micromeria croatica (Pers.) Schott. Molecules 2019, 24, 1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzakou, O.; Couladis, M. The essential oil of Micromeria graeca (L.) Bentham et Reichenb. growing in Greece. Flavour Fragr. J. 2001, 16, 107–109. [Google Scholar] [CrossRef]
- Mincheva, I.; Jordanova, M.; Benbassat, N.; Aneva, I.; Kozuharova, E. Ethnobotany and exploitation of medicinal plants in the Rhodope Mountains—is there a hazard for Clinopodium dalmaticum? Pharmacia 2019, 66, 49–52. [Google Scholar] [CrossRef]
- Zheljazkov, V.D.; Micalizzi, G.; Semerdjieva, I.; Mondello, L. Chemical composition of the essential oil of the endemic species Micromeria frivaldszkyana (Degen) Velen. Molecules 2019, 24, 440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xavier, J.K.A.M.; Maia, L.; Figueiredo, P.L.B.; Folador, A.; Ramos, A.R.; Andrade, E.H.; Maia, J.G.S.; Setzer, W.N.; da Silva, J.K.R. Essential oil composition and DNA barcode and identification of Aniba species (Lauraceae) growing in the Amazon region. Molecules 2021, 26, 1914. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Sauve, R.J.; Long, D.; Cherry, M. Genetic and phytochemical diversity assessment among eleven Hypericum accessions via AFLP and HPLC analyses. J. Herbs Spices Med. Plants 2007, 12, 97–105. [Google Scholar] [CrossRef]
- Pluhár, Z.; Kocsis, M.; Kuczmog, A.; Csete, S.; Simkó, H.; Sárosi, S.; Molnár, P.; Horváth, G. Essential oil composition and preliminary molecular study of four Hungarian Thymus species. Acta. Biol. Hung. 2012, 63, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Stökl, J.; Schlüter, P.M.; Stuessy, T.F.; Paulus, H.F.; Assum, G.; Ayasse, M. Scent variation and hybridization cause the displacement of a sexually deceptive orchid species. Am. J. Bot. 2008, 95, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Trindade, H.; Costa, M.M.; Sofia, B.L.A.; Pedro, L.G.; Figueiredo, A.C.; Barroso, J.G. Genetic diversity and chemical polymorphism of Thymus caespititius from Pico, São Jorge and Terceira islands (Azores). Biochem. Syst. Ecol. 2008, 36, 790–797. [Google Scholar] [CrossRef]
- Trindade, H.; Costa, M.M.; Lima, S.B.; Pedro, L.G.; Figueiredo, A.C.; Barroso, J.G. A combined approach using RAPD, ISSR and volatile analysis for the characterization of Thymus caespititius from Flores, Corvo and Graciosa islands (Azores, Portugal). Biochem. Syst. Ecol. 2009, 37, 670–677. [Google Scholar] [CrossRef]
- Lima, A.S.; Trindade, H.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Volatile and molecular analysis of Juniperus brevifolia (Seub.) Antoine, an Azorean endemic species. Biochem. Syst. Ecol. 2010, 38, 621–629. [Google Scholar] [CrossRef]
- Emami-Tabatabaei, S.-S.; Larijani, K.; Mehregan, I. Application and limitation of molecular data and essential oil content in identification of Leutea elbursensis Mozaff in northern Iran. Acta Bot. Croat. 2018, 77, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carović-Stanko, K.; Liber, Z.; Politeo, O.; Strikić, F.; Kolak, I.; Milos, M.; Šatovic, Z. Molecular and chemical characterization of the most widespread Ocimum species. Plant Syst. Evol. 2011, 294, 253–262. [Google Scholar] [CrossRef]
- Schönswetter, P.; Tribsch, A. Vicariance and dispersal in the alpine perennial Bupleurum stellatum L. (Apiaceae). Taxon 2005, 54, 725–732. [Google Scholar] [CrossRef] [Green Version]
- Ehrich, D. AFLPDAT: A collection of R functions for convenient handling of AFLP data. Mol. Ecol. Notes 2006, 6, 603–604. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication, 1st ed.; University of Illinois Press: Urbana, IL, USA, 1949; 117p. [Google Scholar]
- Lewontin, R.C. The Apportionment of Human Diversity. In Evolutionary Biology; Dobzhansky, T., Hecht, M.K., Steere, W.C., Eds.; Springer: New York, NY, USA, 1972; pp. 381–398. [Google Scholar] [CrossRef]
- Zhivotovsky, L.A. Estimating population structure in diploids with multilocus dominant DNA markers. Mol. Ecol. 1999, 8, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Vekemans, X.; Beauwens, T.; Lemaire, M.; Roldán-Ruiz, I. Data from amplified fragment length polymorphism (AFLP) markers show indication of size homoplasy and of a relationship between degree of homoplasy and fragment size. Mol. Ecol. 2002, 11, 139–151. [Google Scholar] [CrossRef]
- Dice, L.R. Measures of the amount of ecologic association between species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitocondrial DNA restriction sites. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Corander, J.; Waldmann, P.; Sillanpää, M.J. Bayesian analysis of genetic differentiation between populations. Genetics 2003, 163, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.L.; Liu, J.X. StructureSelector: A web-based software to select and visualize the optimal number of clusters using multiple methods. Mol. Ecol. Resour. 2018, 18, 176–177. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. CLUMPAK: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [Green Version]
- Corander, J.; Siren, J.; Arjas, E. Bayesian spatial modeling of genetic population structure. Comput. Stat. 2008, 23, 111–129. [Google Scholar] [CrossRef]
- Corander, J.; Marttinen, P. Bayesian identification of admixture events using multilocus molecular markers. Mol. Ecol. 2006, 15, 2833–2843. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4.1 ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2017. [Google Scholar]
- NIST02 Chemistry WebBook; National Institute Standards and Technology, U.S. Department of Commerce: Gaithersburg, MD, USA, 2022. Available online: https://webbook.nist.gov/ (accessed on 12 March 2022). [CrossRef]
- SAS Institute Inc. Base SAS® 9.3 Procedures Guide: Statistical Procedures; SAS Institute Inc.: Cary, NC, USA, 2011. [Google Scholar]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar]
- Rohlf, F.J. Exeter Software (Firm) NTSYS-Pc: Numerical Taxonomy and Multivariate Analysis System; Applied Biostatistics, Inc.: Setauket, NY, USA, 2009. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa According to: | Sample Site | Voucher No. | Altitude | |||||
|---|---|---|---|---|---|---|---|---|
| [1] (Code) | Balkan Literature | (Population) | (m) | |||||
| M. cristata ssp. cristata (Mc1) | M. cristata | Humsko brdo Mt (Serbia) | HFK-HR-51126 | 387 | ||||
| M. cristata ssp. cristata (Mc2) | M. cristata | Vitosha Mt (Bulgaria) | HFK-HR-51132 | 980 | ||||
| M. cristata ssp. cristata (Mc3) | M. cristata | Demir Kapija (N. Macedonia) | HFK-HR-51141 | 111 | ||||
| M. cristata ssp. cristata (Mc4) | M. cristata | Nomos Serron (Greece) | HFK-HR-51145 | 190 | ||||
| M. cristata ssp. cristata (Mc5) | M. cristata | Pαikon Mt (Greece) | HFK-HR-51146 | 1650 | ||||
| M. cristata ssp. kosaninii (McK) | M. kosaninii | Pletvar (N. Macedonia) | HFK-HR-51142 | 1020 | ||||
| M. croatica (Mcr1) | M. croatica | Rossijev kuk (Croatia) | HFK-HR-51016 | 1641 | ||||
| M. croatica (Mcr2) | M. croatica | Bačić kuk (Croatia) | HFK-HR-51012 | 1159 | ||||
| M. croatica (Mcr3) | M. croatica | Stupačinovo (Croatia) | HFK-HR-51017 | 1058 | ||||
| M. croatica (Mcr4) | M. croatica | Bojinac (Croatia) | HFK-HR-51013 | 1046 | ||||
| M. croatica (Mcr5) | M. croatica | Prezid (Croatia) | HFK-HR-51010 | 991 | ||||
| M. croatica (Mcr6) | M. croatica | Diva Grabovica (BIH) | HFK-HR-51007 | 251 | ||||
| M. croatica (Mcr7) | M. croatica | Dubočani (BIH) | HFK-HR-51020 | 715 | ||||
| M. croatica (Mcr8) | M. croatica | Rakitnica (BIH) | HFK-HR-51019 | 943 | ||||
| M. croatica (Mcr9) | M. croatica | Glavatičevo (BIH) | HFK-HR-51005 | 366 | ||||
| M. croatica (Mcr10) | M. croatica | Babji zub (Montenegro) | HFK-HR-51003 | 1438 | ||||
| M. croatica (McrP1) | M. pseudocroatica | Pijavičino (Croatia) | HFK-HR-51032 | 443 | ||||
| M. croatica (McrP2) | M. pseudocroatica | Prapratno (Croatia) | HFK-HR-51033 | 159 | ||||
| M. graeca ssp. graeca (Mg1) | M. graeca | Malo zlo polje (Croatia) | HFK-HR-51036 | 137 | ||||
| M. graeca ssp. graeca (Mg2) | M. graeca | Komiža (Croatia) | HFK-HR-51037 | 43 | ||||
| M. graeca ssp. fruticulosa (MgF) | M. fruticulosa | Sušac Island (Croatia) | HFK-HR-51038 | 10 | ||||
| M. juliana (Mj1) | M. juliana | Omiška Dinara Mt (Croatia) | HFK-HR-51048 | 91 | ||||
| M. juliana (Mj2) | M. juliana | Sniježnica Mt (Croatia) | HFK-HR-51041 | 512 | ||||
| M. juliana (Mj3) | M. juliana | Lastva (BIH) | HFK-HR-51049 | 382 | ||||
| M. juliana (Mj4) | M. juliana | Lovćen Mt (Montenegro) | HFK-HR-51056 | 948 | ||||
| M. juliana (Mj5) | M. juliana | Krivošije Mt (Montenegro) | HFK-HR-51059 | 194 | ||||
| M. juliana (Mj6) | M. juliana | Cijevna Canyon (Montenegro) | HFK-HR-51045 | 157 | ||||
| M. juliana (Mj7) | M. juliana | Babuna River Canyon (N. Macedonia) | HFK-HR-51052 | 179 | ||||
| M. juliana (Mj8) | M. juliana | Rajec Reka (N. Macedonia) | HFK-HR-51168 | 199 | ||||
| M. juliana (Mj9) | M. juliana | Cholomon Mt (Greece) | HFK-HR-51188 | 1100 | ||||
| M. juliana (Mj10) | M. juliana | Nomos Serron (Greece) | HFK-HR-51147 | 182 | ||||
| M. kerneri (Mk1) | M. kerneri | Zavratnica (Croatia) | HFK-HR-51018 | 161 | ||||
| M. kerneri (Mk2) | M. kerneri | Starigrad Paklenica (Croatia) | HFK-HR-51063 | 83 | ||||
| M. kerneri (Mk3) | M. kerneri | Gradina (Croatia) | HFK-HR-51061 | 207 | ||||
| M. kerneri (Mk4) | M. kerneri | Korčula Island (Croatia) | HFK-HR-51064 | 219 | ||||
| M. kerneri (Mk5) | M. kerneri | Mostar (BIH) | HFK-HR-51044 | 65 | ||||
| M. longipedunculata (Ml1) | M. parviflora | Jazina (BIH) | HFK-HR-51046 | 342 | ||||
| M. longipedunculata (Ml2) | M. parviflora | Krivošije Mt (Montenegro) | HFK-HR-51047 | 619 | ||||
| M. longipedunculata (Ml3) | M. parviflora | Nikšić (Montenegro) | HFK-HR-51066 | 605 | ||||
| M. longipedunculata (Ml4) | M. parviflora | Cijevna Canyon (Montenegro) | HFK-HR-51067 | 161 | ||||
| M. microphylla (Mm) | M. microphylla | Svetac Island (Croatia) | HFK-HR-51039 | 28 | ||||
| C. dalmaticum (Cd1) | M. dalmatica | Mt Lovćen (Montenegro) | HFK-HR-51073 | 1424 | ||||
| C. dalmaticum (Cd2) | M. dalmatica | Mt Orjen (Montenegro) | HFK-HR-51051 | 1074 | ||||
| C. dalmaticum (CdB1) | M. bulgarica | Uhlovitsa cave (Bulgaria) | HFK-HR-51076 | 1040 | ||||
| C. dalmaticum (CdB2) | M. bulgarica | Mesta River Valley (Bulgaria) | HFK-HR-51134 | 580 | ||||
| C. dalmaticum (CdB3) | M. bulgarica | Vlahina Mt (Bulgaria) | HFK-HR-51135 | 1140 | ||||
| C. frivaldszkyanum (Cf1) | M. frivaldszkyana | Ostrusha peak (Bulgaria) | HFK-HR-51093 | 1405 | ||||
| C. frivaldszkyanum (Cf2) | M. frivaldszkyana | Vikanata Skala Nature Monument (Bulgaria) | HFK-HR-51137 | 1040 | ||||
| C. pulegium (Cp) | M. pulegium | Međeđa (BIH) | HFK-HR-51050 | 571 | ||||
| C. serpyllifolium (Cs) | M. albanica | Prizren (Kosovo) | HFK-HR-51074 | 367 | ||||
| C. thymifolium (Ct1) | M. thymifolia | Učka Mt (Croatia) | HFK-HR-51077 | 1189 | ||||
| C. thymifolium (Ct2) | M. thymifolia | Dokozina plan (Croatia) | HFK-HR-51082 | 1441 | ||||
| C. thymifolium (Ct3) | M. thymifolia | Šušanj (Croatia) | HFK-HR-51081 | 604 | ||||
| C. thymifolium (Ct4) | M. thymifolia | Panos (Croatia) | HFK-HR-51084 | 1148 | ||||
| C. thymifolium (Ct5) | M. thymifolia | Blidinje (BIH) | HFK-HR-51042 | 1195 | ||||
| C. thymifolium (Ct6) | M. thymifolia | Diva Grabovica (BIH) | HFK-HR-51086 | 252 | ||||
| C. thymifolium (Ct7) | M. thymifolia | Manastir Morača (Montenegro) | HFK-HR-51053 | 301 | ||||
| Taxa (Code) | Latitude (N) | Longitude (E) | n | p(%) | Npr | I | DW | HE (FIS = 0) |
| M. cristata (Mc1) | 43°22′45.0″ | 21°53′50.1″ | 9 | 25.68 | 1 | 0.185 | 2445.84 | 0.093 |
| M. cristata (Mc2) | 42°29′33.4″ | 23°11′43.1″ | 7 | 26.92 | 1 | 0.204 | 3253.74 | 0.109 |
| M. cristata (Mc3) | 41°24′18.1″ | 22°15′47.0″ | 12 | 41.68 | 2 | 0.287 | 3606.54 | 0.118 |
| M. cristata (Mc4) | 41°15′10.3″ | 23°24′49.8″ | 8 | 34.65 | 2 | 0.256 | 2785.33 | 0.117 |
| M. cristata (Mc5) | 40°57′21.2″ | 22°20′02.0″ | 3 | 19.54 | 0 | 0.179 | 2700.39 | 0.122 |
| M. cristata ssp. kosaninii (McK) | 41°22′09.0″ | 21°39′06.1″ | 10 | 41.56 | 0 | 0.295 | 4153.29 | 0.130 |
| M. croatica (Mcr1) | 44°45′51.1″ | 4°59′17.1″ | 5 | 14.58 | 0 | 0.117 | 1242.40 | 0.073 |
| M. croatica (Mcr2) | 44°34′45.2″ | 15°05′49.5″ | 5 | 14.70 | 0 | 0.118 | 1139.31 | 0.075 |
| M. croatica (Mcr3) | 44°32′37.5″ | 15°10′04.7″ | 5 | 16.47 | 0 | 0.133 | 1311.92 | 0.083 |
| M. croatica (Mcr4) | 44°20′57.7″ | 15°24′50.3″ | 5 | 18.95 | 0 | 0.155 | 1508.70 | 0.090 |
| M. croatica (Mcr5) | 44°15′17.8″ | 15°48′58.2″ | 5 | 14.34 | 0 | 0.115 | 1237.31 | 0.073 |
| M. croatica (Mcr6) | 43°35′59.0″ | 17°41′04.3″ | 5 | 18.54 | 0 | 0.155 | 1357.73 | 0.090 |
| M. croatica (Mcr7) | 43°35′10.1″ | 18°04′44.0″ | 4 | 15.76 | 0 | 0.135 | 1540.99 | 0.085 |
| M. croatica (Mcr8) | 43°34′10.1″ | 18°05′59.2″ | 5 | 18.06 | 0 | 0.147 | 1232.93 | 0.083 |
| M. croatica (Mcr9) | 43°30′21.1″ | 18°06′20.3″ | 5 | 17.77 | 0 | 0.143 | 1379.19 | 0.084 |
| M. croatica (Mcr10) | 42°52′28.2″ | 19°23′05.1″ | 5 | 19.48 | 0 | 0.162 | 2049.82 | 0.095 |
| M. croatica (McrP1) | 42°57′01.5″ | 17°21′52.2″ | 10 | 25.86 | 1 | 0.178 | 2318.02 | 0.085 |
| M. croatica (McrP2) | 42°49′28.1″ | 17°40′23.7″ | 10 | 29.93 | 1 | 0.207 | 3004.68 | 0.097 |
| M. graeca (Mg1) | 43°03′43.3″ | 16°12′55.9″ | 4 | 18.00 | 0 | 0.154 | 4188.99 | 0.108 |
| M. graeca (Mg2) | 43°02′17.4″ | 16°05′49.5″ | 5 | 19.48 | 0 | 0.158 | 4575.80 | 0.105 |
| M. graeca ssp. fruticulosa (MgF) | 43°02′13.4″ | 16°05′50.5″ | 11 | 28.34 | 4 | 0.195 | 6622.21 | 0.099 |
| M. juliana (Mj1) | 42°26′40.4″ | 13°36′90.7″ | 10 | 30.64 | 0 | 0.218 | 1840.88 | 0.112 |
| M. juliana (Mj2) | 42°32′53.9″ | 18°22′02.3″ | 9 | 33.23 | 0 | 0.234 | 1617.68 | 0.112 |
| M. juliana (Mj3) | 42°41′45.8″ | 18°29′26.9″ | 10 | 24.50 | 0 | 0.171 | 1287.44 | 0.091 |
| M. juliana (Mj4) | 42°24′26.0″ | 18°47′15.9″ | 5 | 23.44 | 0 | 0.190 | 1584.60 | 0.117 |
| M. juliana (Mj5) | 42°29′47.8″ | 18°38′57.1″ | 5 | 20.43 | 0 | 0.162 | 1386.40 | 0.103 |
| M. juliana (Mj6) | 42°25′44.6″ | 19°28′53.5″ | 8 | 38.61 | 1 | 0.287 | 2257.51 | 0.131 |
| M. juliana (Mj7) | 41°41′02.6″ | 21°48′11.7″ | 10 | 38.96 | 0 | 0.273 | 1685.18 | 0.123 |
| M. juliana (Mj8) | 41°26′12.2″ | 21°52′06.4″ | 8 | 31.29 | 1 | 0.240 | 2009.83 | 0.117 |
| M. juliana (Mj9) | 40°27′30.0″ | 23°31′04.6″ | 10 | 35.01 | 1 | 0.246 | 2766.48 | 0.122 |
| M. juliana (Mj10) | 41°16′10.8″ | 23°25′06.7″ | 9 | 31.17 | 0 | 0.227 | 1691.23 | 0.116 |
| M. kerneri (Mk1) | 44°42′02.2″ | 14°54′45.6″ | 10 | 25.44 | 0 | 0.177 | 2007.33 | 0.099 |
| M. kerneri (Mk2) | 44°17′35.1″ | 15°26′35.1″ | 8 | 26.92 | 0 | 0.200 | 1598.30 | 0.108 |
| M. kerneri (Mk3) | 43°45′55.9″ | 15°59′12.1″ | 9 | 26.39 | 0 | 0.198 | 1352.29 | 0.103 |
| M. kerneri (Mk4) | 42°57′04.5″ | 17°05′56.1″ | 8 | 23.61 | 0 | 0.171 | 1409.70 | 0.095 |
| M. kerneri (Mk5) | 43°20′36.6″ | 17°48′37.9″ | 10 | 20.72 | 0 | 0.139 | 1489.38 | 0.079 |
| M. longipedunculata (Ml1) | 42°42′15.9″ | 18°30′33.3″ | 10 | 24.85 | 0 | 0.166 | 1668.95 | 0.081 |
| M. longipedunculata (Ml2) | 42°32′46.2″ | 18°42′35.4″ | 5 | 16.94 | 1 | 0.137 | 2381.71 | 0.085 |
| M. longipedunculata (Ml3) | 42°46′02.7″ | 18°57′24.0″ | 10 | 24.50 | 0 | 0.166 | 1503.14 | 0.082 |
| M. longipedunculata (Ml4) | 42°25′44.6″ | 19°28′53.5″ | 10 | 29.46 | 0 | 0.196 | 1736.85 | 0.088 |
| M. microphylla (Mm) | 43°01′07.8″ | 15°45′08.2″ | 9 | 34.30 | 0 | 0.239 | 2097.51 | 0.120 |
| C. dalmaticum (Cd1) | 42°23′45.6″ | 18°50′09.5″ | 4 | 19.42 | 0 | 0.168 | 2225.44 | 0.106 |
| C. dalmaticum (Cd2) | 42°33′45.1″ | 18°37′36.6″ | 4 | 18.77 | 0 | 0.160 | 3159.10 | 0.107 |
| C. dalmaticum (CdB1) | 41°30′49.9″ | 24°39′35.2″ | 9 | 35.48 | 1 | 0.248 | 2764.17 | 0.113 |
| C. dalmaticum (CdB2) | 41°40′46.6″ | 23°43′29.7″ | 9 | 36.30 | 1 | 0.255 | 2861.36 | 0.114 |
| C. dalmaticum (CdB3) | 41°50′30.6″ | 22°59′27.8″ | 8 | 33.53 | 0 | 0.243 | 2693.05 | 0.116 |
| C. frivaldszkyanum (Cf1) | 42°43′54.7″ | 25°15′53.9″ | 8 | 26.33 | 0 | 0.190 | 1953.05 | 0.100 |
| C. frivaldszkyanum (Cf2) | 42°45′57.6″ | 25°30′08.1″ | 9 | 29.75 | 0 | 0.210 | 2592.09 | 0.104 |
| C. pulegium (Cp) | 43°43′57.6″ | 19°11′10.9″ | 3 | 13.75 | 0 | 0.126 | 2032.69 | 0.101 |
| C. serpyllifolium (Cs) | 42°11′52.1″ | 20°46′11.2″ | 3 | 19.78 | 0 | 0.182 | 2514.92 | 0.123 |
| C. thymifolium (Ct1) | 45°17′08.1″ | 14°12′02.4″ | 9 | 26.68 | 0 | 0.193 | 1632.85 | 0.095 |
| C. thymifolium (Ct2) | 44°39′04.3″ | 15°02′39.1″ | 10 | 24.73 | 0 | 0.168 | 1472.09 | 0.085 |
| C. thymifolium (Ct3) | 44°31′33.8″ | 15°06′45.1″ | 9 | 27.92 | 0 | 0.198 | 1787.01 | 0.096 |
| C. thymifolium (Ct4) | 44°26′06.1″ | 15°16′35.2″ | 10 | 27.80 | 0 | 0.195 | 1415.32 | 0.094 |
| C. thymifolium (Ct5) | 43°31′07.8″ | 17°23′09.8″ | 10 | 22.02 | 0 | 0.156 | 1719.07 | 0.081 |
| C. thymifolium (Ct6) | 43°35′59.0″ | 17°41′04.3″ | 9 | 24.73 | 0 | 0.176 | 1522.22 | 0.090 |
| C. thymifolium (Ct7) | 42°45′50.9″ | 19°23′34.6″ | 9 | 23.79 | 1 | 0.169 | 1985.02 | 0.088 |
| Taxon Code | Number of Populations | Population Variation (%) | Φ | p (Φ) | |
|---|---|---|---|---|---|
| Among | Within | ||||
| Mc | 5 | 24.82 | 75.18 | 0.248 | <0.0001 |
| McK | 1 | – | – | – | – |
| Mcr | 10 | 14.37 | 85.63 | 0.144 | <0.0001 |
| McrP | 2 | 12.97 | 87.03 | 0.130 | <0.0001 |
| Mg | 2 | 5.04 | 94.96 | 0.050 | 0.1184 |
| MgF | 1 | – | – | – | – |
| Mj | 10 | 15.99 | 84.01 | 0.160 | <0.0001 |
| Mk | 5 | 34.55 | 65.55 | 0.344 | <0.0001 |
| Ml | 4 | 20.44 | 79.56 | 0.204 | <0.0001 |
| Mm | 1 | – | – | – | – |
| Cd | 2 | 11.39 | 88.61 | 0.114 | 0.0293 |
| CdB | 3 | 13.26 | 86.74 | 0.133 | <0.0001 |
| Cf | 2 | 7.75 | 92.25 | 0.077 | <0.0002 |
| Cp | 1 | – | – | – | – |
| Cs | 1 | – | – | – | – |
| Ct | 7 | 18.03 | 81.97 | 0.180 | <0.0001 |
| Analysis Between | Source of Variation | df | Variance Components | Percent of Total Variance | Φ | p (Φ) |
|---|---|---|---|---|---|---|
| Between taxa | 1 | 1.247 | 0.87 | 0.009 | 0.336 | |
| Mc and McK | Among populations within taxa | 4 | 33.494 | 23.49 | 0.237 | 0.0001 |
| Within populations | 43 | 107.859 | 75.64 | 0.244 | 0.0001 | |
| Between taxa | 1 | 25.412 | 16.76 | 0.168 | 0.0001 | |
| Mc1,2,4 and | Among populations within taxa | 4 | 18.385 | 12.12 | 0.146 | 0.0001 |
| Mc3,5, McK | Within populations | 43 | 107.859 | 71.12 | 0.289 | 0.0001 |
| Between taxa | 1 | 42.040 | 33.49 | 0.335 | 0.0001 | |
| Mcr and McrP | Among populations within taxa | 10 | 11.098 | 8.84 | 0.133 | 0.0001 |
| Within populations | 57 | 72.401 | 57.67 | 0.423 | 0.0001 | |
| Between taxa | 1 | 103.963 | 55.11 | 0.551 | 0.0001 | |
| Mg and MgF | Among populations within taxa | 1 | 4.010 | 2.13 | 0.047 | 0.0001 |
| Within populations | 17 | 80.671 | 42.76 | 0.572 | 0.0001 | |
| Between taxa | 1 | 2.174 | 1.79 | 0.018 | 0.0001 | |
| Mk1–5 and Mm | Among populations within taxa | 4 | 39.215 | 32.32 | 0.329 | 0.0001 |
| Within populations | 48 | 79.927 | 65.88 | 0.341 | 0.0001 | |
| Between taxa | 1 | 10.966 | 9.76 | 0.098 | 0.197 | |
| Mk1,2,4,5 and | Among populations within taxa | 3 | 22.807 | 20.29 | 0.225 | 0.0001 |
| Mm | Within populations | 40 | 78.629 | 69.95 | 0.300 | 0.0001 |
| Between taxa | 1 | 26.066 | 18.22 | 0.182 | 0.002 | |
| Mj and Mk1–5 | Among population within taxa | 13 | 25.796 | 18.03 | 0.220 | 0.0001 |
| Within populations | 114 | 91.237 | 63.76 | 0.362 | 0.0001 | |
| Between taxa | 1 | 42.786 | 27.66 | 0.277 | 0.0001 | |
| Mj and Mk1,2,4,5 | Among populations within taxa | 12 | 20.317 | 13.13 | 0.181 | 0.0001 |
| Within populations | 106 | 91.601 | 59.21 | 0.408 | 0.0001 | |
| Between taxa | 1 | 33.181 | 21.91 | 0.219 | 0.0001 | |
| Cd and CdB | Among populations within taxa | 3 | 15.084 | 9.96 | 0.128 | 0.0001 |
| Within populations | 29 | 103.201 | 68.13 | 0.319 | 0.0001 | |
| Between taxa | 1 | 27.520 | 19.62 | 0.196 | 0.0001 | |
| CdB and Cf | Among populations within taxa | 3 | 13.551 | 9.66 | 0.120 | 0.0001 |
| Within populations | 38 | 99.228 | 70.73 | 0.293 | 0.0001 | |
| Between taxa | 1 | 45.458 | 32.20 | 0.322 | 0.336 | |
| Cd and Cf | Among populations within taxa | 2 | 8.599 | 6.09 | 0.090 | 0.0001 |
| Within populations | 21 | 87.135 | 61.71 | 0.383 | 0.0001 |
| Compound | PC1 | PC2 | ||
|---|---|---|---|---|
| α-Pinene | 0.237 | ns | −0.010 | ns |
| β-Pinene | 0.275 | * | 0.059 | ns |
| α-Campholenal | 0.083 | ns | 0.157 | ns |
| Menthone | −0.446 | *** | −0.182 | ns |
| Camphor | 0.264 | * | 0.692 | *** |
| Pinocarvone | −0.186 | ns | 0.191 | ns |
| Borneol | 0.364 | ** | 0.814 | *** |
| Verbenone | 0.654 | *** | −0.558 | *** |
| Pulegone | −0.744 | *** | −0.245 | ns |
| Piperitonene | −0.360 | ** | 0.142 | ns |
| Piperitone oxide | −0.484 | *** | −0.151 | ns |
| E-Caryophyllene | 0.625 | *** | −0.194 | ns |
| Germacrene D | 0.463 | *** | 0.479 | *** |
| Spathulenol | −0.290 | * | −0.037 | ns |
| Caryophyllene oxide | 0.836 | *** | −0.335 | * |
| α-Muurolol | −0.041 | ns | −0.130 | ns |
| α-Cadinol | 0.105 | ns | 0.305 | * |
| α-Bisabolol | −0.084 | ns | 0.122 | ns |
| Eudesm-7(11)-en-4-ol | −0.023 | ns | −0.081 | ns |
| Thymol | 0.278 | * | −0.330 | * |
| Docosane | 0.250 | ns | −0.635 | *** |
| Eigenvalue | 3.468 | 2.677 | ||
| % of variance | 16.51 | 12.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kremer, D.; Dunkić, V.; Radosavljević, I.; Bogunić, F.; Ivanova, D.; Ballian, D.; Stešević, D.; Matevski, V.; Ranđelović, V.; Eleftheriadou, E.; et al. Phytochemicals and Their Correlation with Molecular Data in Micromeria and Clinopodium (Lamiaceae) Taxa. Plants 2022, 11, 3407. https://doi.org/10.3390/plants11233407
Kremer D, Dunkić V, Radosavljević I, Bogunić F, Ivanova D, Ballian D, Stešević D, Matevski V, Ranđelović V, Eleftheriadou E, et al. Phytochemicals and Their Correlation with Molecular Data in Micromeria and Clinopodium (Lamiaceae) Taxa. Plants. 2022; 11(23):3407. https://doi.org/10.3390/plants11233407
Chicago/Turabian StyleKremer, Dario, Valerija Dunkić, Ivan Radosavljević, Faruk Bogunić, Daniella Ivanova, Dalibor Ballian, Danijela Stešević, Vlado Matevski, Vladimir Ranđelović, Eleni Eleftheriadou, and et al. 2022. "Phytochemicals and Their Correlation with Molecular Data in Micromeria and Clinopodium (Lamiaceae) Taxa" Plants 11, no. 23: 3407. https://doi.org/10.3390/plants11233407