

Cell Membrane Features as Potential Breeding Targets to Improve Cold Germination Ability of Seeds

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
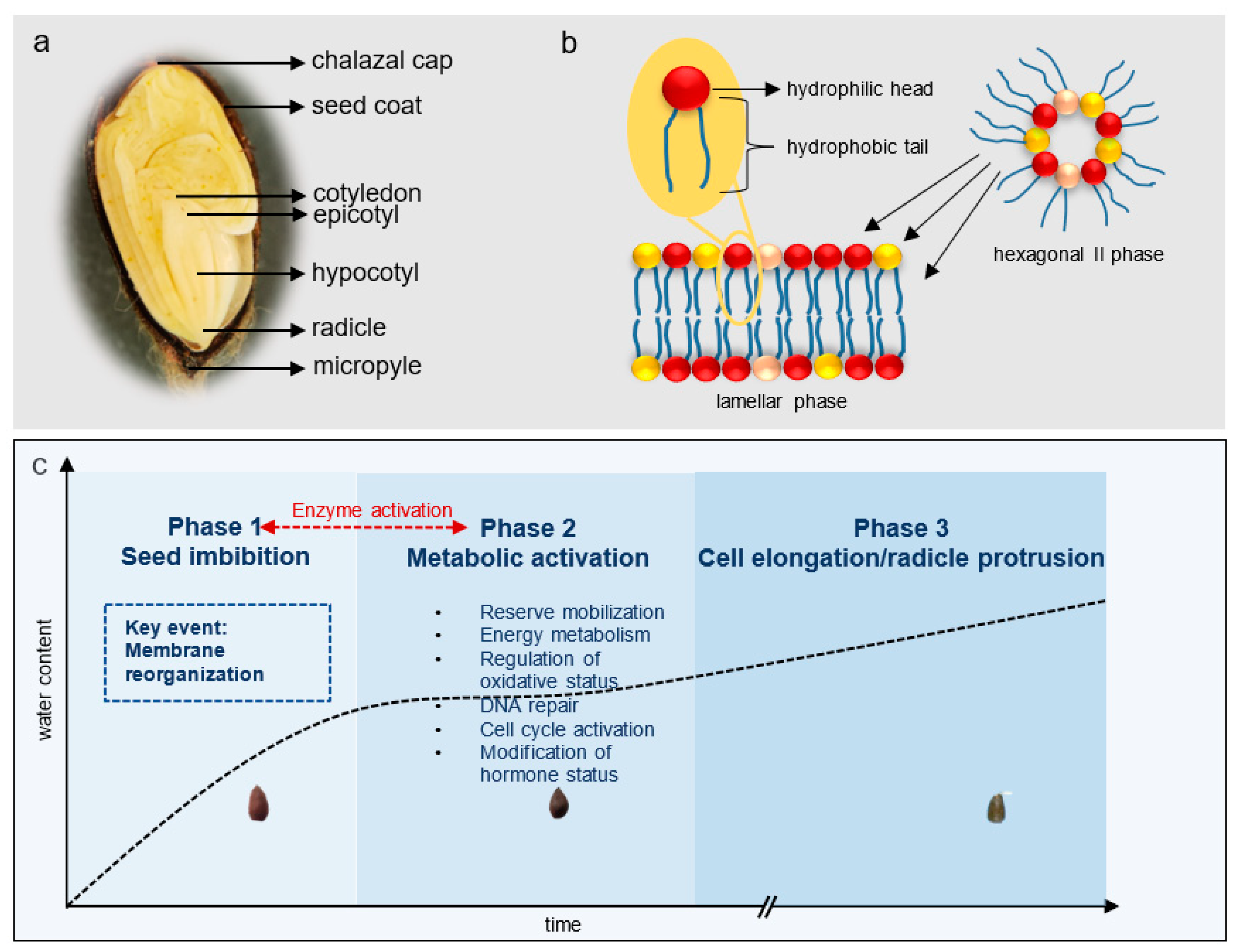
2. Role of Cell Membranes in Early Germination
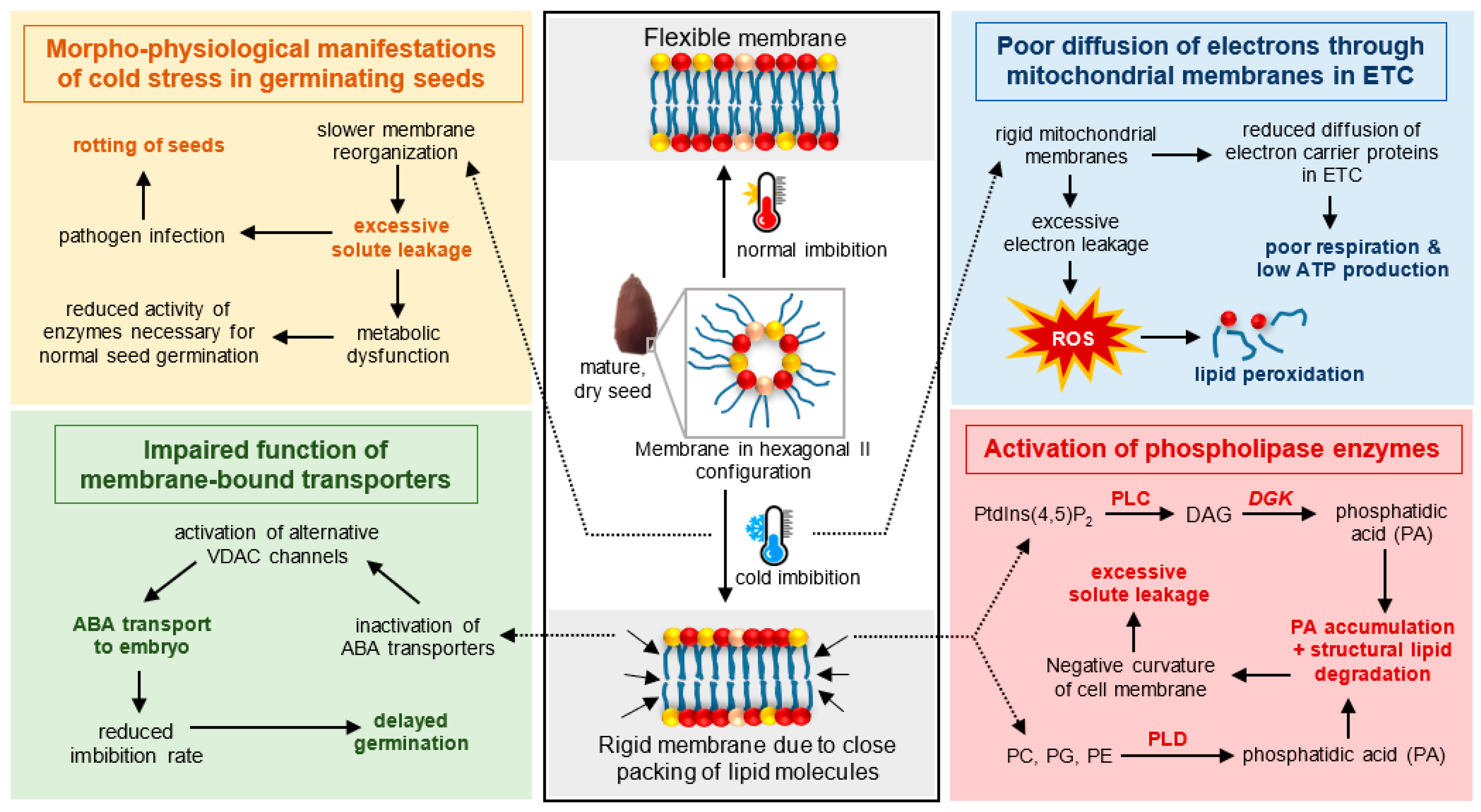
3. Cellular Responses to Cold-Induced Stress in Cell Membranes
4. Modifying Cell Membrane Properties towards Enhancing Cold Germination Ability: A Case Study in Cotton
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hassan, M.A.; Xiang, C.; Farooq, M.; Muhammad, N.; Yan, Z.; Hui, X.; Yuanyuan, K.; Bruno, A.K.; Lele, Z.; Jincai, L. Cold stress in wheat: Plant acclimation responses and management strategies. Front. Plant Sci. 2021, 12, 676884. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://ipad.fas.usda.gov/highlights/2008/02/MassiveSnowStorm.htm (accessed on 1 October 2022).
- Available online: https://www.abc.net.au/news/2018-09-16/wa-farmers-facing-frost-damage-as-chilly-spell-sweeps-south/10253186 (accessed on 1 October 2022).
- Available online: https://today.tamu.edu/2021/03/02/agricultural-losses-from-winter-storm-exceed-600-million/ (accessed on 1 October 2022).
- Shim, J.; Gannaban, R.B.; De Los Reyes, B.G.; Angeles-Shim, R.B. Identification of novel sources of genetic variation for the improvement of cold germination ability in upland cotton (Gossypium hirsutum). Euphytica 2019, 215, 190. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Muhammad, I.; Lan, H.; Xia, C. Recent Advances in the Analysis of Cold Tolerance in Maize. Front. Plant Sci. 2022, 13, 866034. [Google Scholar] [CrossRef] [PubMed]
- Casto, A.L.; Murphy, K.M.; Gehan, M.A. Coping with cold: Sorghum cold stress from germination to maturity. Crop. Sci. 2021, 61, 3894–3907. [Google Scholar] [CrossRef]
- Pan, Y.; Liang, H.; Gao, L.; Dai, G.; Chen, W.; Yang, X.; Qing, D.; Gao, J.; Wu, H.; Huang, J.; et al. Transcriptomic profiling of germinating seeds under cold stress and characterization of the cold-tolerant gene LTG5 in rice. BMC Plant Biol. 2020, 20, 371. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, A.; Li, W. How membranes organize during seed germination: Three patterns of dynamic lipid remodelling define chilling resistance and affect plastid biogenesis. Plant Cell Environ. 2015, 38, 1391–1403. [Google Scholar] [CrossRef] [Green Version]
- Stanghellini, M.E.; Hancock, J.G. The sporangium of Pythium ultimum as a survival structure in the soil. Phytopathology 1973, 61, 157–164. [Google Scholar] [CrossRef]
- Keeling, B. Soybean seed rot and the relation of seed exudate to host susceptibility. Phytopathology 1974, 64, 1445–1447. [Google Scholar] [CrossRef]
- Serrano, M.; Robertson, A. Cold stress at planting increase susceptibility to damping-off caused by Pythium sylvaticum. Phytopathology 2016, 106, 40. [Google Scholar]
- Martin, F.N.; Loper, J.E. Soilborne plant diseases caused by Pythium spp.: Ecology, epidemiology, and prospects for biological control. Crit. Rev. Plant Sci. 1999, 18, 111–181. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Bassel, G.W. Seed vigour and crop establishment: Extending performance beyond adaptation. J. Exp. Bot. 2016, 67, 567–591. [Google Scholar] [CrossRef] [PubMed]
- Jame, Y.W.; Cutforth, H.W. Simulating the effects of temperature and seeding depth on germination and emergence of spring wheat. Agric. For. Meteorol. 2004, 124, 207–218. [Google Scholar] [CrossRef]
- Verheul, M.J.; Picatto, C.; Stamp, P. Growth and development of maize seedlings under chilling conditions in the field. Eur. J. Agron. 1996, 5, 31–43. [Google Scholar] [CrossRef]
- Hope, H.J.; White, R.P.; Dwyer, L.M.; Maamari, R.; Seguin, S.; Hamilton, R.I. Low temperature emergence potential of short season maize hybrids grown under controlled environment and plot conditions. Can. J. Plant Sci. 1992, 72, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Kuraparthy, V.; Fang, H.; Zhu, L.; Sood, S.; Jones, D.C. High-density linkage map construction and QTL analyses for fiber quality, yield and morphological traits using CottonSNP63K array in upland cotton (Gossypium hirsutum L.). BMC Genom. 2019, 20, 889. [Google Scholar] [CrossRef] [Green Version]
- Steponkus, P.L. Role of plasma membrane in freezing injury and cold acclimation. Annu. Rev. Plant Physiol. 1984, 35, 543–584. [Google Scholar] [CrossRef]
- Sanghera, G.S.; Wani, S.H.; Hussain, W.; Singh, N.B. Engineering cold stress tolerance in crop plants. Curr. Genoms. 2011, 12, 30–43. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Wang, X. Crosstalk between phospholipase D and sphingosine kinase in plant stress signaling. Front. Plant Sci. 2012, 3, 51. [Google Scholar] [CrossRef] [Green Version]
- Raju, S.K.K.; Barnes, A.C.; Schnable, J.C.; Roston, R.L. Low-temperature tolerance in land plants: Are transcript and membrane responses conserved? Plant Sci. 2018, 276, 73–86. [Google Scholar] [CrossRef]
- Ali, A.S.; Elozeir, A.A. Metabolic processes during seed germination. In Advances in Seed Biol; Jimenez-Lopez, J.C., Ed.; IntechOpen: London, UK, 2017; pp. 141–166. [Google Scholar]
- Simpson, D.M.; Adams, C.L.; Stone, G.M. Anatomical Structure of the Cottonseed Coat as Related to Problems of Germination; Tech. Bull., No. 734; US Department of Agriculture: Washington, DC, USA, 1940; pp. 1–23.
- Sanchez, J.; Mangat, P.K.; Angeles-Shim, R.B. Weathering the cold: Modifying membrane and storage fatty acid composition of seeds to improve cold germination ability in upland cotton (Gossypium hirsutum L.). Agron. J. 2019, 9, 684. [Google Scholar] [CrossRef] [Green Version]
- Simon, E.W. Phospholipids and plant membrane permeability. New Phytol. 1974, 73, 377–420. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manz, B.; Muller, K.; Kucera, B.; Volke, F.; Leubner-Metzger, G. Water uptake and distribution in germinating tobacco seeds investigated in vivo by nuclear magnetic resonance imaging. Plant Physiol. 2005, 138, 1538–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uemura, M.; Joseph, R.A.; Steponkus, P.L. Cold acclimation of Arabidopsis thaliana (effect on plasma membrane lipid composition and freeze-induced lesions). Plant Physiol. 1995, 109, 15–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miquel, M.; James, D., Jr.; Dooner, H.; Browse, J. Arabidopsis requires polyunsaturated lipids for low-temperature survival. Proc. Natl. Acad. Sci. USA 1993, 90, 6208–6212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlova, I.V.; Serebriiskaya, T.S.; Popov, V.; Merkulova, N.; Nosov, A.M.; Trunova, T.I.; Tsydendambaev, V.D.; Los, D.A. Transformation of tobacco with a gene for the thermophilic acyl-lipid desaturase enhances the chilling tolerance of plants. Plant Cell Physiol. 2003, 44, 447–450. [Google Scholar] [CrossRef] [Green Version]
- Barrero-Sicilia, C.; Silvestre, S.; Haslam, R.P.; Michaelson, L.V. Lipid remodelling: Unravelling the response to cold stress in Arabidopsis and its extremophile relative Eutrema salsugineum. Plant Sci. 2017, 263, 194–200. [Google Scholar] [CrossRef]
- Cahoon, E.B.; Lindqvist, Y.; Schneider, G.; Shanklin, J. Redesign of soluble fatty acid desaturases from plants for altered substrate specificity and double bond position. Proc. Natl. Acad. Sci. USA 1997, 94, 4872–4877. [Google Scholar] [CrossRef] [Green Version]
- Nachtschatt, M.; Okada, S.; Speight, R. Integral membrane fatty acid desaturases: A review of biochemical, structural, and biotechnological advances. Eur. J. Lipid. Sci. Technol. 2020, 122, 2000181. [Google Scholar] [CrossRef]
- Browse, J.; Somerville, C. Glycerolipid synthesis: Biochemistry and regulation. Annl. Rev. Plant Biol. 1991, 42, 467–506. [Google Scholar] [CrossRef]
- Dong, C.J.; Cao, N.; Zhang, Z.G.; Shang, Q.M. Characterization of the fatty acid desaturase genes in cucumber: Structure, phylogeny, and expression patterns. PLoS ONE 2016, 11, e0149917. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, W.; He, Q.; Daud, M.K.; Chen, J.; Zhu, S. Characterization of 19 genes encoding membrane-bound fatty acid desaturases and their expression profiles in Gossypium raimondii under low temperature. PLoS ONE 2015, 10, e0123281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dar, A.A.; Choudhury, A.R.; Kancharla, P.K.; Arumugam, N. The FAD2 gene in plants: Occurrence, regulation, and role. Front. Plant Sci. 2017, 8, 1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noblet, A.; Leymarie, J.; Bailly, C. Chilling temperature remodels phospholipidome of Zea mays seeds during imbibition. Sci. Rep. 2018, 7, 8886. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yesbergenova-Cuny, Z.; Biniek, C.; Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. Revisiting the role of ethylene and N-end rule pathway on chilling-induced dormancy release in Arabidopsis seeds. Int. J. Mol. Sci. 2018, 19, 3577. [Google Scholar] [CrossRef] [Green Version]
- Al Amin, N.; Ahmad, N.; Wu, N.; Pu, X.; Ma, T.; Du, Y.; Bo, X.; Wang, N.; Sharif, R.; Wang, P. CRISPR-Cas9 mediated targeted disruption of FAD2–2 microsomal omega-6 desaturase in soybean (Glycine max. L). BMC Biotechnol. 2019, 19, 1–10. [Google Scholar] [CrossRef]
- Zhukovsky, M.A.; Filograna, A.; Luini, A.; Corda, D.; Valente, C. Phosphatidic acid in membrane rearrangements. Febs Lett. 2019, 593, 2428–2451. [Google Scholar] [CrossRef] [Green Version]
- Testerink, C.; Munnik, T. Phosphatidic acid: A multifunctional stress signaling lipid in plants. Trends Plant Sci. 2005, 10, 368–375. [Google Scholar] [CrossRef]
- Oblozinsky, M.; Ulbrich-Hofmann, R.; Bezakova, L. Head group specificity of phospholipase D isoenzymes from poppy seedlings (Papaver somniferum L.). Biotechnol. Lett. 2005, 27, 181–185. [Google Scholar] [CrossRef]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14 (Suppl. 1), S165–S183. [Google Scholar] [CrossRef] [PubMed]
- Katagiri, T.; Lshiyama, K.; Kato, T.; Tabata, S.; Kobayashi, M.; Shinozaki, K. An important role of phosphatidic acid in ABA signaling during germination in Arabidopsis thaliana. Plant J. 2005, 43, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Munnik, T. Phosphatidic acid: An emerging plant lipid second messenger. Trends Plant Sci. 2001, 6, 227–233. [Google Scholar] [CrossRef]
- Huo, C.; Zhang, B.; Wang, H.; Wang, F.; Liu, M.; Gao, Y.; Zhang, W.; Deng, Z.; Sun, D.; Tang, W. Comparative study of early cold-regulated proteins by two-dimensional difference gel electrophoresis reveals a key role for phospholipase Dα1 in mediating cold acclimation signaling pathway in rice. Mol. Cell. Proteom. 2016, 15, 1397–1411. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.H.; Henderson, D.A.; Zhu, J.K. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell 2005, 17, 3155–3175. [Google Scholar] [CrossRef] [Green Version]
- Dhaliwal, L.K.; Gannaban, R.B.; Shrestha, A.; Shim, J.; Mangat, P.K.; Singleton, J.J.; Angeles-Shim, R.B. Integrated morpho-biochemical and transcriptome analyses reveal multidimensional response of upland cotton (Gossypium hirsutum L.) to low temperature stress during seedling establishment. Plant Environ. Interact. 2021, 2, 290–302. [Google Scholar] [CrossRef]
- Madhusudanan, K.N.; Johnson, K.M. Solute leakage from winged bean (Psophocarpus tetragonolobus) seeds. Seed Sci. Technol. 1990, 18, 157–162. [Google Scholar]
- Nelson, E.B. Rapid germination of sporangia of Pythium species in response to volatiles from germinating seeds. Phytopathology 1987, 77, 1108–1112. [Google Scholar] [CrossRef]
- Nelson, E.B.; Craft, C.M. Comparative germinationofculture-producedand plant-produced sporangia of Pythium ultimum in response to soluble seed exudates and exudate components. Phytopathology 1989, 79, 1009–1013. [Google Scholar] [CrossRef]
- Wolfe, J.; Steponkus, P.L. The stress-strain relation of the plasma membrane of isolated plant protoplasts. Biochim. Biophys. Acta Biomembr. 1981, 643, 663–668. [Google Scholar] [CrossRef]
- Willing, R.P.; Leopold, A.C. Cellular expansion at low temperatures as a cause of membrane lesions. Plant Physiol. 1983, 71, 118–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Wei, B.; Gao, Z.; Zhou, Y.; Shi, F.; Zhou, X.; Zhou, Q.; Ji, S. Changes in membrane lipid composition and function accompanying chilling injury in Bell Peppers. Plant Cell Physiol. 2017, 59, 167–178. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, R.; Zhang, F.; Tao, F.; Li, W. Lipid profiling and tolerance to low-temperature stress in Thellungiella salsuginea in comparison with Arabidopsis thaliana. Biol. Plant 2013, 57, 149–153. [Google Scholar] [CrossRef]
- Gohrbandt, M.; Lipski, A.; Grimshaw, J.W.; Buttress, J.A.; Baig, Z.; Herkenhoff, B.; Walter, S.; Kurre, R.; Deckers-Hebestreit, G.; Strahl, H. Low membrane fluidity triggers lipid phase separation and protein segregation in living bacteria. EMBO J. 2022, 41, e109800. [Google Scholar] [CrossRef] [PubMed]
- Budin, I.; de Rond, T.; Chen, Y.; Chan, L.J.G.; Petzold, C.J.; Keasling, J.D. Viscous control of cellular respiration by membrane lipid composition. Science 2018, 362, 1186–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Han, Q.; Ding, C.; Huang, Y.; Liao, J.; Chen, T.; Feng, S.; Zhou, L.; Zhang, Z.; Chen, Y.; et al. Effect of low temperature on chlorophyll biosynthesis and chloroplast biogenesis of rice seedlings during greening. Int. J. Mol. Sci. 2020, 21, 1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, J.S.P.; Rajendrak Prasad, S.; Banerjee, R.; Thammineni, C. Seed birth to death: Dual functions of reactive oxygen species in seed physiology. Ann. Bot. 2015, 116, 663–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blokhina, O.; Fagerstedt, K.V. Oxidative metabolism, ROS and NO under oxygen deprivation. Plant Physiol. Biochem. 2010, 48, 359–373. [Google Scholar]
- Barba-Espín, G.; Hernández, J.A.; Diaz-Vivancos, P. Role of H2O2 in pea seed germination. Plant Signal. Behave. 2012, 7, 193–195. [Google Scholar] [CrossRef] [Green Version]
- Wojtyla, L.; Lechowska, K.; Kubala, S.; Garnczarska, M. Different modes of hydrogen peroxide action during seed germination. Front. Plant Sci. 2016, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Roxas, V.P.; Lodhi, S.A.; Garrett, D.K.; Mahan, J.R.; Allen, R.D. Stress tolerance in transgenic tobacco seedlings that overexpress glutathione S-transferase/glutathione peroxidase. Plant Cell Physiol. 2000, 41, 1229–1234. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar]
- Iqbal, N.; Hussain, S.; Raza, M.A.; Yang, C.Q.; Safdar, M.E.; Brestic, M.; Aziz, A.; Hayyat, M.S.; Asghar, M.A.; Wang, X.C.; et al. Drought tolerance of soybean (Glycine max L. Merr.) by improved photosynthetic characteristics and an efficient antioxidant enzyme activities under a split-root system. Front. Physiol. 2019, 10, 786. [Google Scholar] [CrossRef] [Green Version]
- Daszkowska-Golec, A. Arabidopsis seed germination under abiotic stress as a concert of action of phytohormones. OMICS A J. Integr. Biol. 2011, 15, 763–774. [Google Scholar] [CrossRef]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef]
- Vishal, B.; Kumar, P.P. Regulation of Seed Germination and Abiotic Stresses by Gibberellins and Abscisic Acid. Front. Plant Sci. 2018, 9, 838. [Google Scholar]
- Rajjou, L.; Huguet, R.; Robin, C.; Belghazi, M.; Job, C. Proteomic investigation of the effect of salicylic acid on Arabidopsis seed germination and establishment of early defense mechanisms. Plant Physiol. 2006, 141, 910–923. [Google Scholar] [CrossRef] [Green Version]
- Steber, C.M.; McCourt, P. A role for brassinosteroids in germination in Arabidopsis. Plant Physiol. 2001, 125, 763–769. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.; Ma, Y.; Chen, Y.; Chen, X.; Wei, A. Plant Hormone Response to Low-Temperature Stress in Cold-Tolerant and Cold-Sensitive Varieties of Zanthoxylum bungeanum Maxim. Front. Plant Sci. 2022, 13, 847202. [Google Scholar] [CrossRef]
- Cuevas, J.C.; López-Cobollo, R.; Alcázar, R.; Zarza, X.; Koncz, C.; Altabella, T.; Salinas, J.; Tiburcio, A.F.; Ferrando, A. Putrescine is involved in Arabidopsis freezing tolerance and cold acclimation by regulating abscisic acid levels in response to low temperature. Plant Physiol. 2008, 148, 1094–1105. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A.; Akihiro, T.; Ito, K.; Kumagai, S.; Sugita, R.; Tanoi, K.; Rahman, A. ATP binding cassette proteins ABCG37 and ABCG33 function as potassium-independent cesium uptake carriers in Arabidopsis roots. Mol. Plant 2021, 14, 664–678. [Google Scholar] [CrossRef] [PubMed]
- Arif, Y.; Sami, F.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salicylic acid in relation to other phytohormones in plant: A study towards physiology and signal transduction under challenging environment. Environ. Exp. Bot. 2020, 175, 104040. [Google Scholar]
- Meena, Y.K.; Kaur, N. Salicylic acid enhanced low temperature stress tolerance in vegetables: A review. Chem. Sci. Rev. Lett. 2017, 6, 2428–2435. [Google Scholar]
- Lee, Y.; Choi, Y.B.; Suh, S.; Lee, J.; Assmann, S.M.; Joe, C.O.; Kelleher, J.E.; Crain, R.C. Abscisic acid-induced phosphoinositide turnover in guard cell protoplasts of Vicia faba. Plant Physiol. 1996, 110, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Ohme-Takagi, M.; Shinshi, H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell 1995, 7, 173–182. [Google Scholar] [PubMed] [Green Version]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate regulates the inducer of cbf expression-C-repeat binding factor/DRE binding factor1 cascade and freezing tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Truong, T.T.; Barrero, J.M.; Jacobsen, J.V.; Hocart, C.H.; Gubler, F. A role for jasmonates in the release of dormancy by cold stratification in wheat. J. Exp. Bot. 2016, 67, 3497–3508. [Google Scholar] [CrossRef]
- Hussain, S.; Kim, S.H.; Bahk, S.; Ali, A.; Nguyen, X.C.; Yun, D.J.; Chung, W.S. The Auxin Signaling Repressor IAA8 Promotes Seed Germination Through Down-Regulation of ABI3 Transcription in Arabidopsis. Front. Plant Sci. 2020, 11, 111. [Google Scholar]
- Gangadhar, B.H.; Sajeesh, K.; Venkatesh, J.; Baskar, V.; Abhinandan, K.; Yu, J.W.; Prasad, R.; Mishra, R.K. Enhanced tolerance of transgenic potato plants over-expressing non-specific lipid transfer protein-1 (StnsLTP1) against multiple abiotic stresses. Front. Plant Sci. 2016, 7, 1228. [Google Scholar] [CrossRef] [Green Version]
- Available online: http://www.icac.org (accessed on 15 August 2022).
- Lehman, S.G. Studies on treatment of cotton seed. N. C. Agric. Exp. Stn. Tech. Bull. 1925, 26, 71. [Google Scholar]
- Ludwig, C.A. The germination of cottonseed at low temperature. J. Agric. Res. 1932, 44, 367–380. [Google Scholar]
- Burke, J.J.; Mohan, J.R.; Hatfield, J.L. Crop specific thermal kinetic windows in relation to wheat and cotton biomass production. Agron. J. 1988, 80, 553–556. [Google Scholar] [CrossRef]
- Gipson, J.R.; Ray, L.L.; Flowers, C.L. Influence of night temperature on seed development of five varieties of cotton. In Proceedings of the Beltwide Cotton Production Research Conferences, New Orleans, LA, USA, 7–8 January 1969; Brown, P.B., Ed.; National Cotton Council of America: Memphis, TN, USA, 1969; pp. 117–118. [Google Scholar]
- Krzyzanowski, F.C.; Delouche, J.C. Germination of cotton seed in relation to temperature. Rev. Bras. Sementes. 2011, 33, 543–548. [Google Scholar] [CrossRef] [Green Version]
- Pettigrew, W.T. Improved yield potential with early planting cotton production system. Agron. J. 2002, 94, 997–1003. [Google Scholar] [CrossRef]
- Bange, M.P.; Milroy, S.P. Growth and dry matter partitioning of diverse cotton genotypes. Field. Crops. Res. 2004, 87, 73–87. [Google Scholar] [CrossRef]
- Mauget, S.A.; Adhikari, P.; Leiker, G.; Baumhardt, R.L.; Thorp, K.R.; Ale, S. Modeling the effects of management and elevation on West Texas dryland cotton production. Agric. For. Meterol. 2017, 247, 385–398. [Google Scholar] [CrossRef]
- Radcliffe, J.D.; Czajka-Narins, D.M.; Imrhan, V. Fatty acid composition of serum, adipose tissue and liver in rats fed diets containing corn oil or cottonseed oil. Plant Foods Hum. Nutr. 2004, 69, 73–77. [Google Scholar] [CrossRef]
- Davis, K.E.; Prasad, C.; Imrhan, V. Consumption of a diet rich in cottonseed oil (CSO) lowers total and LDL cholesterol in normo-cholesterolemic subjects. Nutrients 2012, 4, 602–610. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Zhang, Q.; Lv, J.; Qiao, K.; Fan, S.; Ma, Q.; Zhang, C. Genome-Wide Analysis of the Phospholipase D Family in Five Cotton Species, and Potential Role of GhPLD2 in Fiber Development and Anther Dehiscence. Front. Plant Sci. 2021, 12, 728025. [Google Scholar] [CrossRef]
- Thompson, C.N.; Hendon, B.R.; Mishra, D.; Rieff, J.M.; Lowery, C.C. Cotton (Gossypium hirsutum L.) mutants with reduced levels of palmitic acid (C16:0) in seed lipids. Euphytica 2019, 215, 112. [Google Scholar] [CrossRef]
- Chapman, K.D.; Austin-Brown, S.; Sparace, S.A.; Kinney, A.J.; Ripp, K.G.; Pirtle, I.L.; Pirtle, R.M. Transgenic cotton plants with increased seed oleic acid content. J. Am. Oil Chem. Soc. 2001, 78, 941–947. [Google Scholar] [CrossRef]
- Zhang, M.; Fan, J.; Taylor, D.C.; Ohlrogge, J.B. DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell 2009, 21, 3885–3901. [Google Scholar] [CrossRef] [Green Version]
- Snider, J.L.; Pilon, C.; Virk, G. Seed characteristics and seedling vigor. In Cotton Seed Seedlings; Chastain, D.R., Kaur, G., Reddy, K.R., Oosterhuis, D.M., Eds.; The Cotton Foundation: Cordova, TN, USA, 2020; pp. 9–22. [Google Scholar]
- Contreras, F.X.; Ernst, A.M.; Wieland, F.; Brügger, B. Specificity of intramembrane protein-lipid interactions. Cold Spring Harb. Perspect. Biol. 2011, 3, a004705. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Ding, N.Z. Plant Unsaturated Fatty Acids: Multiple Roles in Stress Response. Front. Plant Sci. 2020, 11, 562785. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yue, X.; An, L. Integrated regulation triggered by a cryophyte ω-3 desaturase gene confers multiple-stress tolerance in tobacco. J. Exp. Bot. 2018, 69, 2131–2148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iba, K. Acclimative response to temperature stress in higher plants: Approaches of gene engineering for temperature tolerance. Annu. Rev. Plant Biol. 2002, 53, 225–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugly, S.; Somerville, C. A role for membrane lipid polyunsaturation in chloroplast biogenesis at low temperature. Plant Physiol. 1992, 99, 197–202. [Google Scholar] [CrossRef]
- Doria, E.; Pagano, A.; Ferreri, C.; Vita, L.A.; Anca, M.; de Sousa, A.S.; Almao, B. How does the seed pre-germinative metabolism fight against imbibition damage? Emerging roles of fatty acid cohort and antioxidant defence. Front. Plant Sci. 2019, 10, 1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, A.A.; Oliveira, M.D.A.; Matthews, S. The role of imbibition damage in determining the vigour of white and coloured seed lots of dwarf French bean (Phaseolus vulgaris). J. Exp. Bot. 1986, 37, 716–722. [Google Scholar] [CrossRef]
- Lin, Y.; Xin, X.; Yin, G.; He, J.; Zhou, Y.; Chen, J.; Lu, X. Membrane phospholipids remodeling upon imbibition in Brassica napus L. seeds. Biochem. Biophys. Res. Commun. 2019, 515, 289–295. [Google Scholar] [CrossRef]
- Shewry, P.R.; Pinfield, N.J.; Stobart, A.K. Phospholipids and the phospholipid fatty acids of germinating hazel seeds (Corylus avellana L.). J. Exp. Bot. 1973, 24, 1100–1105. [Google Scholar] [CrossRef]
- Nonogaki, H. MicroRNA gene regulation cascades during early stages of plant development. Plant Cell Physiol. 2010, 51, 1840–1846. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Mittler, R. Reactive oxygen species and temperature stresses: A delicate balance between signaling and destruction. Physiol. Plantarum. 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Saita, E.; Albanesi, D.; de Mendoza, D. Sensing membrane thickness: Lessons learned from cold stress. Biochim. Biophys. Acta 2016, 1861, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hong, Y.; Wang, X. Phospholipase D- and phosphatidic acid-mediated signaling in plants. Biochim. Biophys. Acta 2009, 1791, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Guschina, I.A.; Harwood, J.L. Mechanisms of temperature adaptation in poikilotherms. Febs. Lett. 2006, 580, 5477–5483. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhaliwal, L.K.; Angeles-Shim, R.B. Cell Membrane Features as Potential Breeding Targets to Improve Cold Germination Ability of Seeds. Plants 2022, 11, 3400. https://doi.org/10.3390/plants11233400
Dhaliwal LK, Angeles-Shim RB. Cell Membrane Features as Potential Breeding Targets to Improve Cold Germination Ability of Seeds. Plants. 2022; 11(23):3400. https://doi.org/10.3390/plants11233400
Chicago/Turabian StyleDhaliwal, Lakhvir Kaur, and Rosalyn B. Angeles-Shim. 2022. "Cell Membrane Features as Potential Breeding Targets to Improve Cold Germination Ability of Seeds" Plants 11, no. 23: 3400. https://doi.org/10.3390/plants11233400