
The Outline of Seed Silhouettes: A Morphological Approach to Silene (Caryophyllaceae)
 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. General Morphological Analysis
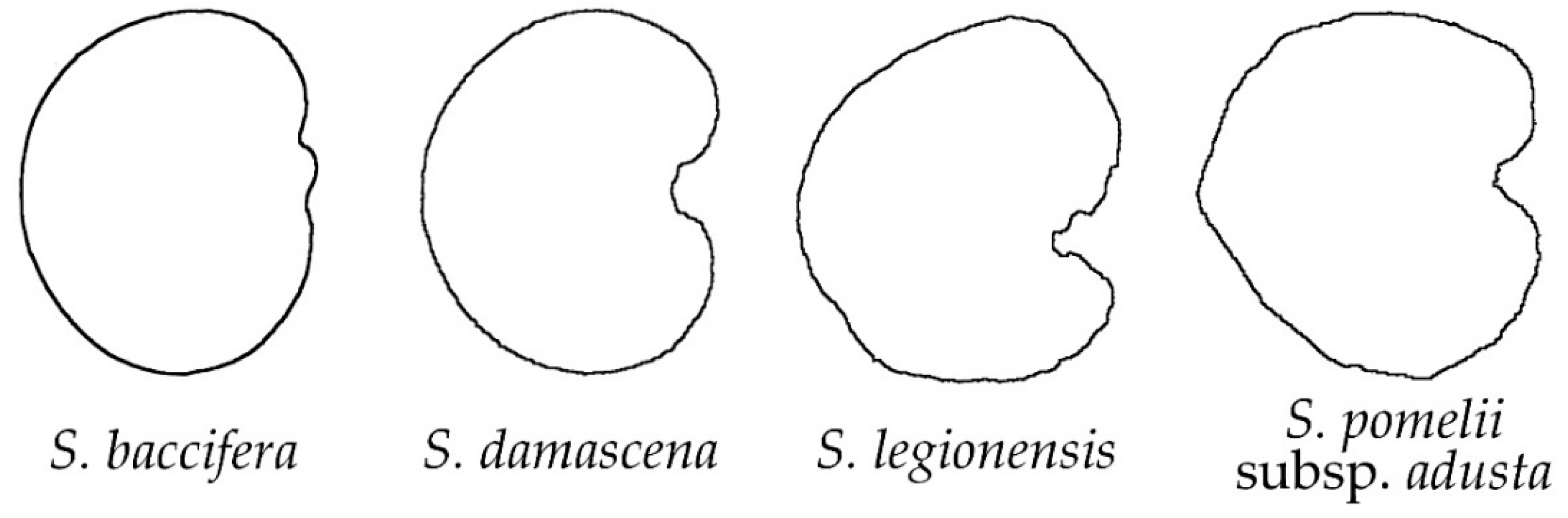
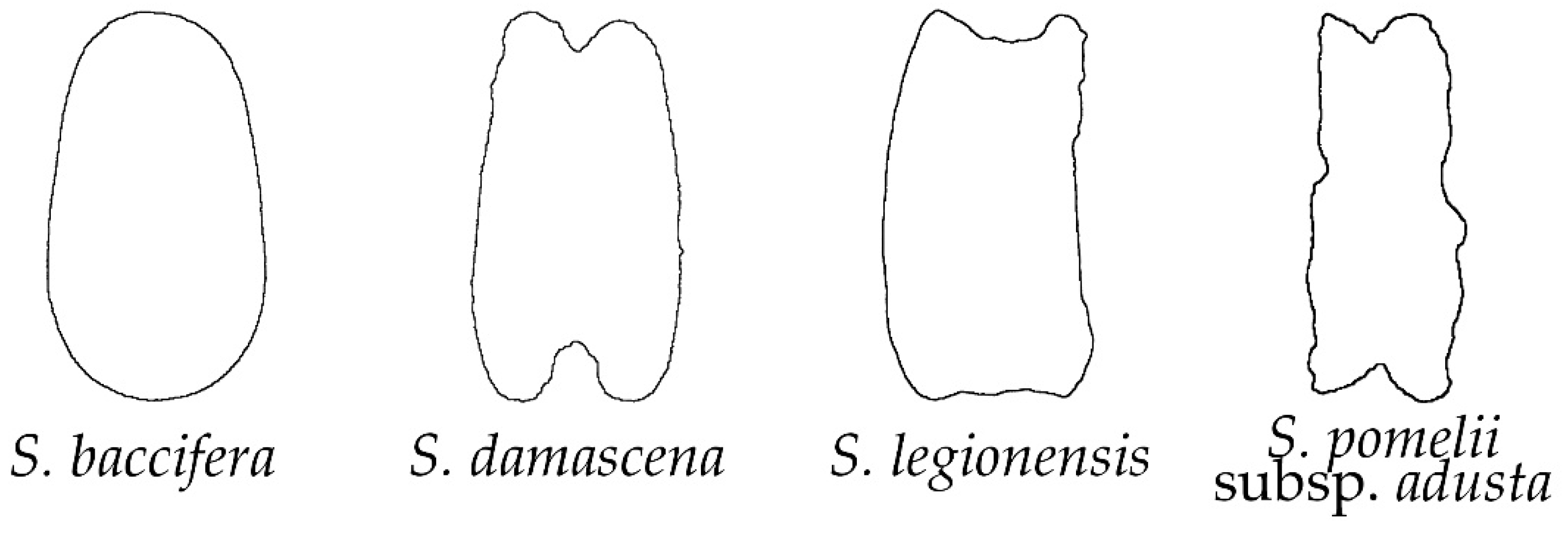
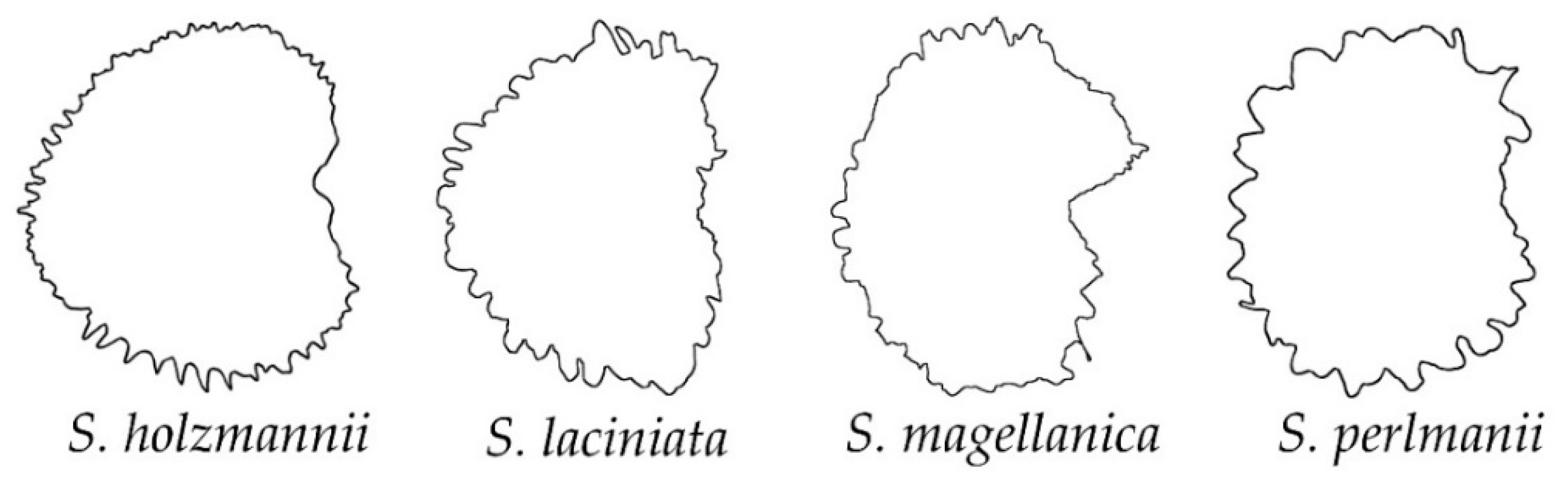
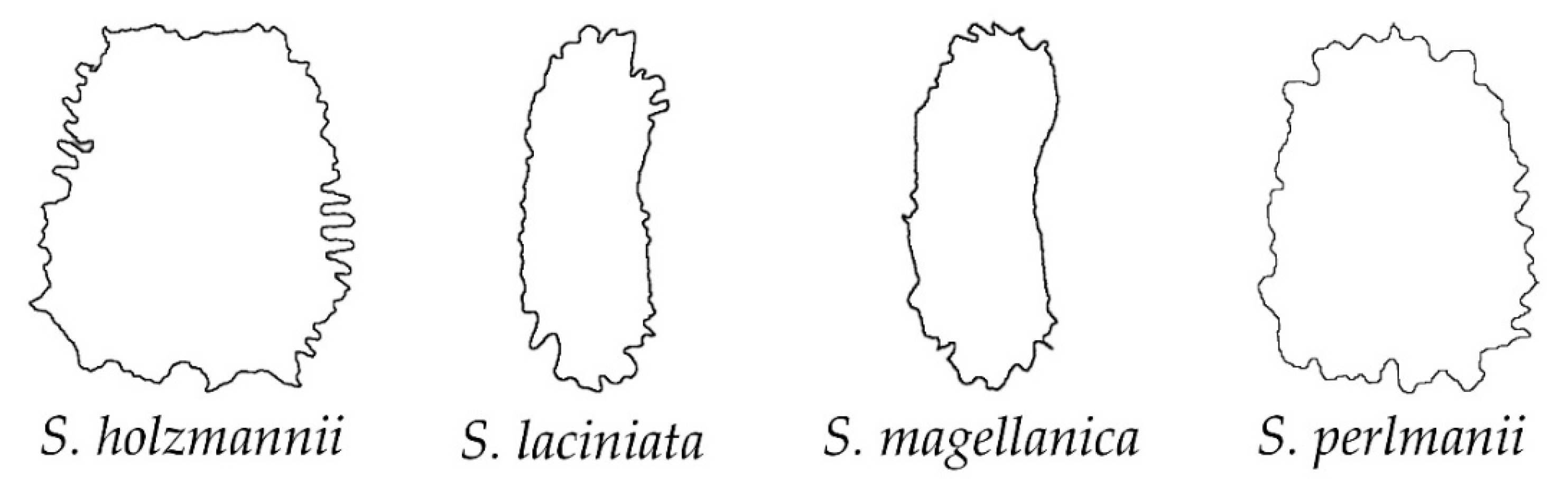
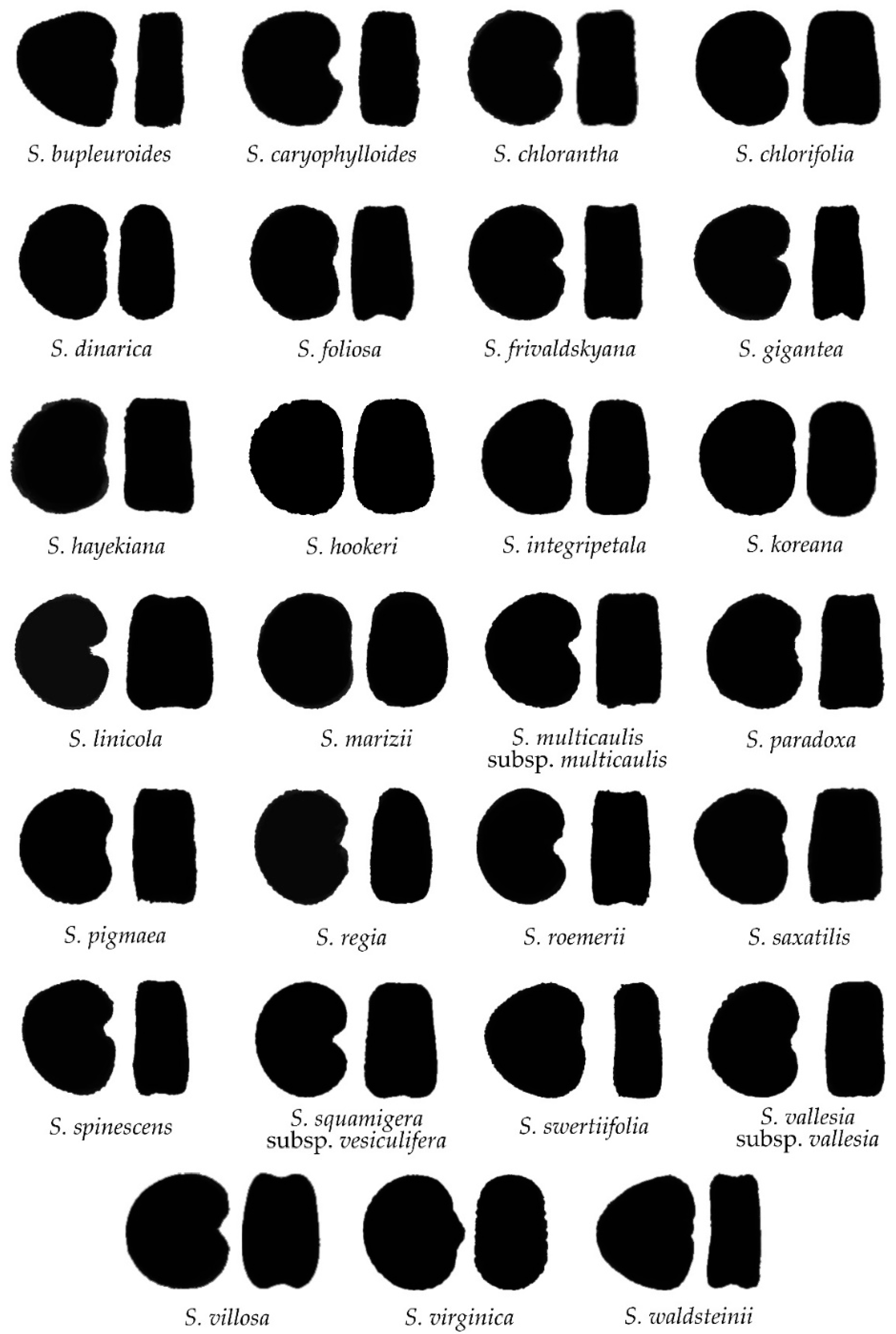
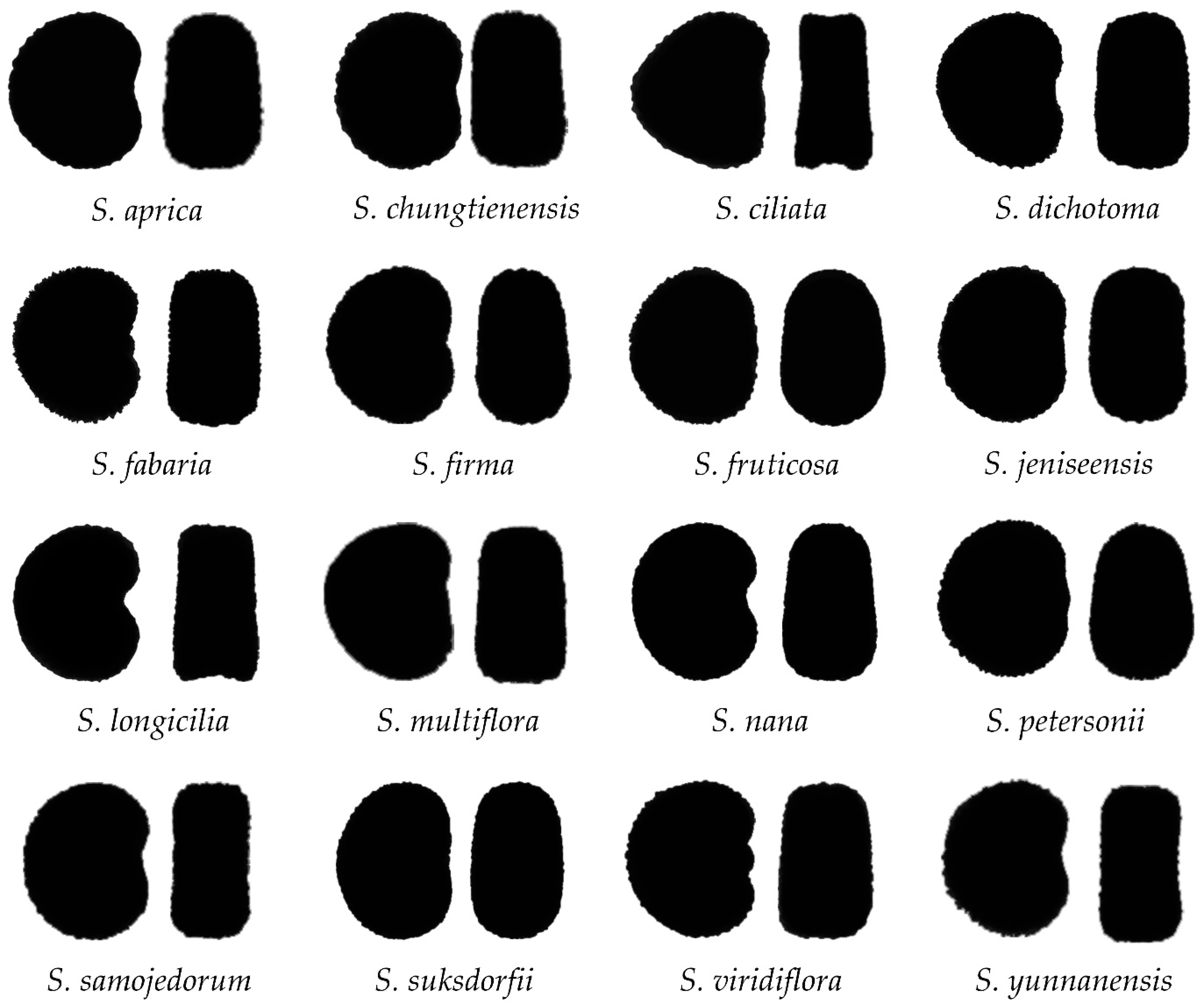
2.2. Morphological Types Based on the Outlines of the Seed Silhouettes
2.3. The Average Silhouettes in Each Morphological Group
2.4. Morphological Comparison between Seed Groups
2.5. The Relationship between Groups and Taxonomic Sections
3. Discussion
4. Materials and Methods
4.1. Seeds
4.2. Seed Images
4.3. Seed Individual Outline and Average Silhouettes
4.4. General Morphological Description
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


Appendix B




Appendix C

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphological Type | Smooth | Rugose | Echinate |
|---|---|---|---|
| S. subg. Silene | S. apetala, S. borderei, S. colorata, S. colpophylla, S. damascena, S. crassipes, S. diversifolia, S. legionensis, S. micropetala, S. nicaeensis, S. pomeli-adusta, S. ramosissima, S. secundiflora, S. vivianii (13) | S. acaulis, S. argaea, S. bupleuroides, S. caryophylloides, S. chlorantha, S. chlorifolia, S. dinarica, S. disticha, S. foliosa, S. frivaldskyana, S. gigantea, S. hayekiana, S. koreana, S. inaperta, S. linicola, S. mellifera, S. multicaulis, S. muscipula, S. nocturna, S. otites, S. paradoxa, S. portensis, S. pygmaea, S. roemeri, S. ruprechtii, S. saxatilis, S. saxifraga, S. scabriflora, S. sclerocarpa, S. spinescens, S. squamigera, S.swertiifolia, S. stricta, S. tridentata, S. valesia, S. villosa, S. waldsteinii, S. wolgensis (37) | S. ciliata, [S. coutinhoi], [S. fruticosa], S. jeniseensis, [S. longicilia], [S. multiflora], [S. nutans], [S. pseudoatocion], [S. viridiflora] (9) |
| S. subg. Behenantha | S. baccifera, S. littorea (2) | S. acutifolia, S. conica, S. elisabethae, S. integripetala, S. marizii, [S. regia], [S. virginica], [S. zawadzkii] (8) | S. aprica, S. behen, S. caroliniana, S. chungtienensis, S. dichotoma, S. diclinis, S. dioica, S. fabaria, S. firma, S. foetida, S. gallica, S.hookeri, S. latifolia, S. nana, S. noctiflora, S. pendula, S. petersonii, S. samojedorum, S. suksdorfii, S. viscosa, S. vulgaris, S. yunnanensis (20) |
| S. subg. Lychnis | S. coronaria | S. chalcedonica, S. flos-cuculi, S. flos-jovis |
| Species | Source | Place of Origin | Subgenus (Section) |
|---|---|---|---|
| Eudianthe coeli-rosa (L.) Fenzl ex Endl. | Jardin Botanique de la ville de Lyon | Haute-Corse (2B), Désert des Agriates, plage de Loto | - |
| Silene aprica Turcz. ex Fisch. & C.A. Mey. | Chollipo Arboretum | Botanical Garden | Behenantha (Physolychnis) |
| S. baccifera (L.) Durande | Kärntner Botanikzentrum | Kärnten: Grafenstein, Sabuatach, glade Pinus sylvestris forest, on conglomerate, 615m (19.8.2012) | Behenantha (Cuccubalus) |
| S. bupleuroides L. | Hortus Botanicus Vacratot, Hungary | - | Silene (Sclerocalycinae) |
| S. caryophylloides (Poir.) Otth | Botanischer Garten der Universität Tübingen | Holubec; Turkei, Ulu Dag | Silene (Auriculatae) |
| S. chlorantha (Willd.) Ehrh. | Botanischer Garten der Universität Potsdam | Germany. Brandenburg, Odergebiet, an der Bahn, SW Bahnhof Podelzig, | Silene (Siphonomorpha) |
| S. chlorifolia Sm. | BG der Martin-Luther-Univ. Halle-Wittenberg | - | Silene (Sclerocalycinae) |
| S. chungtienensis (Speg.) Bocquet | Botanic Garden of the University of Copenhagen | - | Behenantha (Physolychnis) |
| S. ciliata Pourr. | Botany Hung. Acad. of Sciences | Botanical Garden | Silene (Silene) |
| S. damascena Boiss. and Gaill. | The Botanical Garden Tel Aviv University | Mount Hermon | Silene (Silene) |
| S. dichotoma Ehrh. | Botanic Garden of the University of Copenhagen | - | Behenantha (Dichotomae) |
| S. dinarica Spreng. | Botanicka Zahrada Teplice | - | Silene (Siphonomorpha) |
| S. fabaria (L.) Coyte | Botanischer Garten der Universität Bonn | Chakidiki, south of Ouranopolis, towards the border of Athos | Behenantha (Behenantha) |
| S. firma Siebold and Zucc. | The Hiroshima Botanical Garden | North-western Hiroshima pref., a pass near the mountain, 800m, oct 2014 | Behenantha (Physolychnis) |
| S. foliosa Maxim. | Vladivostok Botanical Garden | Gamow Peninsula | Silene (Siphonomorpha) |
| S. frivaldskyana Hampe | Siberian Botanical Garden of Tomsk State University | - | Silene (Siphonomorpha) |
| S. fruticosa L. | Jardin Botanique de Dijon | - | Silene (Siphonomorpha) |
| S. gigantea (L.) L. | Julia & Alexander N. Diomides Botanic Garden | Cult./ATHD | Silene (Siphonomorpha) |
| S. hayekiana Hand.-Mazz. & Janch. | University Botanic Gardens Ljubljana | Mazzeti and Janchen—Kucelj | Silene (Siphonomorpha) |
| S. holzmani Heldr. ex Boiss. | Julia & Alexander N. Diomides Botanic Garden | Spont./Glaronisi islet-Pigadia-Karpathos Island | Behenantha (Behenantha) |
| S. hookeri Nutt. | Botanicka Zahrada Teplice | - | Behenantha (Physolychnis) |
| S. integripetala Bory and Chaub. | BG der Martin-Luther-Univ. Halle-Wittenberg | - | Behenantha (Sedoides) |
| S. jeniseensis Willd. | Vladivostok Botanical Garden | Gamow Peninsula | Silene (Siphonomorpha) |
| S. koreana Kom. | Botanic Garden of Perm State University | - | Silene (Siphonomorpha) |
| S. laciniata Cav. | The Medicinal Herb Garden, Universit of Washington | - | Behenantha (Physolychnis) |
| S. legionensis Lag. | BG Universidade de Coimbra | Alimonde—Bragança | Silene (Silene) |
| S. linicola C.C. Gmel. | Jardin Botanique de la ville de Lyon | Aube (10), entre Auxerre et Troyes | Silene (Lasiocalycinae) |
| S. longicilia (Brot.) Otth | BG Universidade de Coimbra | Serra da Boa Viagem—Figueira da Foz | Silene (Siphonomorpha) |
| S. magellanica (Desr.) Bocquet | Station Alpine du Lautaret. Univ. Joseph Fourier | Punta Arenas (Chili, 10 m) | Behenantha (Physolychnis) |
| S. marizii Samp. | BG Universidade de Coimbra | Nespereira—Celorico da Beira | Behenantha (Melandrium) |
| S. multicaulis Guss. subsp. multicaulis | Bundesgärten Alpengarten im Belvedere (Viena, Austria) | - | Silene (Siphonomorpha) |
| S. multiflora (Erhr.) Pers. | Botany Hung. Acad. of Sciences | Botanical Garden | Silene (Siphonomorpha) |
| S. nana Kar. and Kir. | National Botanical Garden of Iran | Arak, 20 km to Borujerd, Robatmil to Chepeghli village | Behenantha (Saponarioides) |
| S. paradoxa L. | Station Alpine du Lautaret. Univ. Joseph Fourier | Défilé d’Inzecca (Corse, 250 m) | Silene (Siphonomorpha) |
| S. perlmanii W.L.Wagner, D.R.Herbst and Sohmer | Botanischer Garten der Universität Zürich | - | Silene (Sclerophyllae) |
| S. petersonii Maguire | BG der Martin-Luther-univ. Halle-Wittenberg | Mount Brocken Garden | Behenantha (Physolychnis) |
| S. pomelii Batt. subsp. adusta (Ball) Maire | Station Alpine du Lautaret. Univ. Joseph Fourier | Essaouira (Maroc), 0 m | Silene (Silene) |
| S. pygmaea Adams | St Andrews Botanical Garden | - | Silene (Auriculatae) |
| S. regia Sims | Botanischer Garten Universität Hamburg | Nachzucht BG Hamburg (Winona, MN/US; Prairie Moon Nursery) | Behenantha (Physolychnis) |
| S. roemeri Friv. | St Andrews Botanical Garden | - | Silene (Siphonomorpha) |
| S. samojedorum (Sambuk) Oxelman | Hortus Botanicus Patavinus | - | Behenantha (Physolychnis) |
| S. saxatilis Sims | Botanischer Garten München-Nymphenburg | Georgia, Reg. Guria, cerca de Bakhmaro. 1950 m | Silene (Siphonomorpha) |
| S. spinescens Sm. | Julia & Alexander N. Diomides Botanic Garden | Spont./Sounion National Park-Attiki | Silene (Siphonomorpha) |
| S. squamigera Boiss. subsp. vesiculifera (J.Gay ex Boiss.) Coode and Cullen | Botanic Garden of the University of Copenhagen | - | Silene (Lasiocalycinae) |
| S. suksdorfii B.L. Rob. | Botanischer Garten München-Nymphenburg | - | Behenantha (Physolychnis) |
| S. swertiifolia Boiss. | National Botanical Garden of Iran | Elam, Before the tunnel Javar | Silene (Sclerocalycinae) |
| S. vallesia L. subsp. vallesia | Station Alpine du Lautaret. Univ. Joseph Fourier | Villar d’Arène, 1700 m | Silene (Auriculatae) |
| S. villosa Forssk. | The Botanical Garden Tel Aviv University | - | Silene (Silene) |
| S. virginica L. | Botanischer Garten Universität Hamburg | US: North Carolina; Burke County (USA-Reise, 2013) | Behenantha (Physolychnis) |
| S. viridiflora L. | Gradina Botanica “Alexandru Borza” Cluj-Napoca Romania | Botanical Garden | Silene (Siphonomorpha) |
| S. waldsteinii Griseb. | Göteborg Botanical Garden | Greece, Florinis, Mt. Voras | Silene (Siphonomorpha) |
| S. yunnanensis Franch. | Botanischer Garten der Universität Zürich | - | Behenantha (Cucubaloides) |
References
- Melzheimer, V. Caryophyllaceae: Silene L. In Flora Iranica, 163; Rechinger, K.H., Ed.; Akad. Druck-und Verlagsanstalt: Graz, Austria, 1988; pp. 341–508. [Google Scholar]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2022. Available online: https://www.plantsoftheworldonline.org/ (accessed on 15 June 2022).
- Greuter, W. Silene (Caryophyllaceae) in Greece: A Subgeneric and Sectional Classification. Taxon 1995, 44, 543–581. [Google Scholar] [CrossRef]
- Hosny, A.I.; Zareh, M.M. Taxonomic studies of Silenoideae (Caryophyllaceae) in Egypt. 2. Taxonomic significance of seed characters in genus Silene L. with special reference to the seed coat sculpture. Taeckholmia 1993, 14, 37–44. [Google Scholar]
- El Oqlah, A.A.; Karim, F.M. Morphological and anatomical studies of seed coat in Silene species (Caryophyllaceae) from Jordan. Arab Gulf J. Sci. Res. 1990, 8, 121–139. [Google Scholar]
- Ocaña, M.E.; Juan, R.; Fernández, I.; Pastor, J. Estudio morfológico de semillas de Silene (Caryophyllaceae) del suroeste de España. Lagascalia 2011, 31, 21–45. [Google Scholar]
- Martín-Gómez, J.J.; Porceddu, M.; Bacchetta, G.; Cervantes, E. Seed Morphology in Species from the Silene mollissima Aggregate (Caryophyllaceae) by Comparison with Geometric Models. Plants 2022, 11, 901. [Google Scholar] [CrossRef]
- Brullo, C.; Brullo, S.; Giusso del Galdo, G.; Minissale, P.; Sciandrello, S. Silene peloritana (Caryophyllaceae) a new species from Sicily. Phytotaxa 2014, 172, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Bacchetta, G.; Carta, A.; Paradis, G.; Piazza, C.; Peruzzi, L. Further insights into the taxonomy of the Silene nocturna species complex (Caryophyllaceae): A systematic survey of the taxa from Sardinia and Corsica. Phytotaxa 2014, 175, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Sáez, L.; Mesbah, M.; López-Alvarado, J.; Bacchetta, G.; El Mokni, R.; Peruzzi, L.; Oxelman, B. Re-establishment of Silene neglecta Ten. (Caryophyllaceae) with taxonomic notes on some related taxa. PhytoKeys 2022, 195, 143–160. [Google Scholar] [CrossRef]
- Arman, M.; Gholipour, A. Seed morphology diversity in some Iranian endemic Silene (Caryophyllaceae) species and their taxonomic significance. Acta Biol. Szeged. 2013, 57, 31–37. [Google Scholar]
- Gholipour, A.; Kuhdar, F. Seed morphology diversity in the certain Silene species (Caryophyllaceae) from Iran and its taxonomical significance. Taxon. Biosyst. 2014, 6, 107–118. [Google Scholar]
- Keshavarzi, M.; Mahdavinejad, M.; Sheidai, M.; Gholipour, A. Seed and pollen morphology of some Silene species (Caryophyllaceae) in Iran. Phytol. Balc. 2015, 21, 7–12. [Google Scholar]
- Pusalkar, P.K.; Singh, D.K.; Lakshminarasimhan, P. Silene gangotriana (Caryophyllaceae): A new species from Western Himalaya, India. Kew Bull. 2004, 59, 621–624. [Google Scholar] [CrossRef]
- Lin, N.; Zhang, D.-G.; Huang, X.-H.; Zhang, J.-W.; Yang, J.-Y.; Tojibaev, K.; Wang, H.-C.; Deng, T. Silene sunhangii (Caryophyllaceae), a new species from China. PhytoKeys 2019, 135, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moiloa, N.A.; Mesbah, M.; Nylinder, S.; Manning, S.; Forest, F.; de Boer, H.J.; Bacon, C.D.; Oxelman, B. Biogeographic origins of southern African Silene (Caryophyllaceae). Mol. Phylogen. Evol. 2021, 162, 107199. [Google Scholar] [CrossRef]
- Manning, J.C.; Goldblatt, P. A taxonomic revision of the southern African native and naturalized species of Silene L. (Caryophyllaceae). Bothalia 2012, 42, 147–186. [Google Scholar] [CrossRef]
- Francis MacBride, J.; Dahlgreen, B.E. Flora of Peru. Field Mus. Nat. Hist. 1937, 13, 634. Available online: https://www.biodiversitylibrary.org/item/19766#page/9/mode/1up (accessed on 1 December 2022).
- Franquemont, C.; Plowman, T.; Franquemont, E.; King, T.; Niezgoda, C.; Davis, W.; Sperling, C.R. The Ethnobotany of Chinchero an Andean Community in southern Peru. Bot. New Ser. 1990, 24, 49. [Google Scholar]
- Boissier, E. Flora Orientalis; Georg H.: Basel, Switzerland; Geneva, Switzerland, 1867; Volume 1, pp. 567–656. Available online: https://www.biodiversitylibrary.org/item/60323#page/7/mode/1up (accessed on 1 December 2022).
- Rohrbach, P. Monographic der Gattung Silene; Verlag von Engelmann: Leipzig, Germany, 1869; pp. 1–249. Available online: https://www.biodiversitylibrary.org/bibliography/15462 (accessed on 1 December 2022).
- Chowdhuri, P.K. Studies in the genus Silene. Notes Royal Bot. Gard. Edinb. 1957, 22, 221–287. [Google Scholar]
- Ghazanfar, S.A. Seed characters as diagnostic in the perennial sections of the genus Silene L. (Family Caryophyllaceae). Pak. J. Bot. 1983, 15, 7–12. [Google Scholar]
- Fawzi, N.; Fawzy, A.; Mohamed, A. Seed morphological studies on some species of Silene L. (Caryophyllaceae). Int. J. Bot. 2010, 6, 287–292. [Google Scholar] [CrossRef]
- Hoseini, E.; Assadi, M.; Edalatiyan, M.N.; Ghahremaninejad, F. Seed micromorphology and its implication in subgeneric classification of Silene (Caryophyllaceae, Sileneae). Flora 2017, 228, 31–38. [Google Scholar] [CrossRef]
- Atazadeh, N.; Keshavarzi, M.; Sheidai, M.; Gholipour, A. Seed morphology of Silene commelinifolia Boiss. Complex (Caryophyllaceae Juss.). Mod. Phytomorphol. 2017, 11, 5–13. [Google Scholar]
- Ullah, F.; Papini, A.; Shah, S.N.; Zaman, W.; Sohail, A.; Iqbal, M. Seed micromorphology and its taxonomic evidence in subfamily Alsinoideae (Caryophyllaceae). Microsc. Res. Techniq. 2019, 82, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Murru, V.; Grillo, O.; Santo, A.; Ucchesu, M.; Piazza, C.; Gaio, A.; Carta, A.; Bacchetta, G. Seed morpho-colorimetric analysis on some Tyrrhenian species of the Silene mollissima aggregate (Caryophyllaceae). Flora 2019, 258, 151445. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Rewicz, A.; Rodriguez-Lorenzo, J.L.; Janousek, B.; Cervantes, E. Seed morphology in Silene based on geometric models. Plants 2020, 9, 1787. [Google Scholar] [CrossRef]
- Juan, A.; Martín-Gómez, J.J.; Rodriguez-Lorenzo, J.L.; Janousek, B.; Cervantes, E. New techniques for seed shape description in Silene species. Taxonomy 2021, 2, 1–19. [Google Scholar] [CrossRef]
- Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Juan, A.; Tocino, Á.; Janousek, B.; Cervantes, E. Seed Morphological Properties Related to Taxonomy in Silene L. Species. Taxonomy 2022, 2, 298–323. [Google Scholar] [CrossRef]
- Rodríguez-Lorenzo, J.L.; Martín-Gómez, J.J.; Tocino, Á.; Juan, A.; Janoušek, B.; Cervantes, E. New Geometric Models for Shape Quantification of the Dorsal View in Seeds of Silene Species. Plants 2022, 11, 958. [Google Scholar] [CrossRef]
- Gray, A. Modern Differential Geometry of Curves and Surfaces with Mathematica; CRC Press: Boca Raton, FL, USA, 1998; pp. 163–165. [Google Scholar]
- Jafari, F.; Zarre, S.; Gholipour, A.; Eggens, F.; Rabler, R.K.; Oxelman, B. A new taxonomic backbone for the infrageneric classification of the species-rich genus Silene (Caryophyllaceae). Taxon 2020, 69, 337–368. [Google Scholar] [CrossRef]
- Cervantes, E.; Rodríguez-Lorenzo, J.L.; Gutiérrez del Pozo, D.; Martín-Gómez, J.J.; Janousek, B.; Tocino, Á.; Juan, A. Seed Silhouettes as Geometric Objects: New Applications of Elliptic Fourier Transform to Seed Morphology. Horticulturae 2022, 8, 974. [Google Scholar] [CrossRef]
- Prentice, H.C. Numerical analysis of infraspecific variation in European Silene alba and S. dioica (Caryophyllaceae). Bot. J. Linn. Soc. 1979, 78, 181–212. [Google Scholar] [CrossRef]
- Talavera, S. Silene L. Flora Iberica. In Real Jardín Botánico; Castroviejo, S., Laínz, M., López González, G., Monserrat, P., Muñoz Garmendia, F., Paiva, J., Villar, L., Eds.; CSIC: Madrid, Spain, 1990; Volume 2, pp. 313–406. [Google Scholar]
- Cervantes, E.; Martín-Gómez, J.J.; Espinosa-Roldán, F.E.; Muñoz-Organero, G.; Tocino, Á.; Cabello-Sáenz de Santamaría, F. Seed Morphology in Key Spanish Grapevine Cultivars. Agronomy 2021, 11, 734. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ.; U.S. National Institutes of Health: Bethesda, MD, USA, 2018. Available online: http://imagej.nih.gov/ij/ (accessed on 15 June 2022).
- Zdilla, M.J.; Hatfield, S.A.; McLean, K.A.; Cyrus, L.M.; Laslo, J.M.; Lambert, H.W. Circularity, solidity, axes of a best fit ellipse, aspect ratio, and roundness of the foramen ovale: A morphometric analysis with neurosurgical considerations. J. Craniofacial Surg. 2016, 27, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, E.; Martín-Gómez, J.J.; Saadaoui, E. Updated Methods for Seed Shape Analysis. Scientifica 2016, 2016, 5691825. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Campbell, G.; Skillings, J.H. Nonparametric Stepwise Multiple Comparison Procedures. J. Am. Stat. Assoc. 1985, 80, 998–1003. [Google Scholar] [CrossRef]
- Sokal, R.R.; Braumann, C.A. Significance Tests for Coefficients of Variation and Variability Profiles. Syst. Zool. 1980, 29, 50. [Google Scholar] [CrossRef]








| Dorsal View (n = 1012) | Lateral View (n = 1025) | |
|---|---|---|
| Area | Mean value: 0.83 mm2 (SD = 0.46; CV = 55.6) Min. value: 0.15 mm2 (S. chungtienensis) Max. value: 2.47 mm2 (S. linicola) | Mean value: 1.10 mm2 (SD = 0.63; CV = 57.2) Min. value: 0.21 mm2 (S. chungtienensis) Max. value: 4.28 mm2 (S. swertiifolia) |
| Perimeter | Mean value: 3.98 mm (SD = 1.18; CV = 29.6) Min. value: 1.72 mm (S. chungtienensis) Max. value: 8.62 mm (S. fabaria) | Mean value: 4.49mm (SD = 1.46; CV = 32.4) Min. value: 1.95 mm (S. chungtienensis) Max. value: 11.36 mm (S. laciniata) |
| Circularity | Mean value: 0.62 (SD = 0.11; CV = 17.1) Min. value: 0.25 (S. holzmannii) Max. value: 0.84 (A. marizii) | Mean value: 0.66 (SD = 0.13; CV = 19.1) Min. value: 0.21 (S. holzmannii) Max. value: 0.87 (S. marizii) |
| Roundness | Mean value: 0.57 (SD = 0.10; CV = 18.1) Min. value: 0.29 (S. bupleuroides) Max. value: 0.89 (S. petersonii) | Mean value: 0.81 (SD = 0.06; CV = 6.9) Min. value: 0.62 (S. linicola) Max. value: 0.98 (S. paradoxa) |
| Aspect ratio | Mean value: 1.80 (SD = 0.35; CV = 19.2) Min. value: 1.12 (S. petersonii) Max. value: 3.47 (S. bupleuroides) | Mean value: 1.25 (SD = 0.09; CV = 7.1) Min. value: 1.02 (S. paradoxa) Max. value: 1.63 (S. linicola) |
| Solidity | Mean value: 0.934 (SD = 0.03; CV = 3.6) Min. value: 0.76 (S. bupleuroides) Max. value: 0.991 (S. baccifera) | Mean value: 0.946 (SD = 0.02; CV = 2.3) Min. value: 0.805 (S. perlmanii) Max. value: 0.983 (S. baccifera) |
| Seed Type or Species | N | A | P | C | S |
|---|---|---|---|---|---|
| Smooth | 73 | 1.31 b (45.2) | 4.49 b (21.7) | 0.77 d (8.2) | 0.961 d (1.7) |
| Rugose | 540 | 1.19 b (55.4) | 4.47 b (28.3) | 0.71 c (11.2) | 0.955 c (1.3) |
| Echinate | 323 | 0.89 a (59.4) | 4.22 a (37.0) | 0.60 b (17.1) | 0.939 b (1.6) |
| Papillose | 69 | 1.22 b (49.5) | 6.24 c (27.1) | 0.39 a (21.9) | 0.892 a (2.2) |
| E. coeli-rosa | 20 | 0.61 a (17.0) | 3.48 a (10.2) | 0.63 b (7.6) | 0.936 b (0.8) |
| Seed Type or Species | N | A | P | AR | C | R | S |
|---|---|---|---|---|---|---|---|
| Smooth | 69 | 0.88 b (52.5) | 4.01 b (19.6) | 1.87 c (19.4) | 0.64 c (20.2) | 0.55 a (16.7) | 0.922 b (5.6) |
| Rugose | 540 | 0.90 b (52.9) | 4.02 b (26.4) | 1.86 c (18.3) | 0.66 c (11.9) | 0.56 a (17.8) | 0.943 c (2.9) |
| Echinate | 318 | 0.72 a (62.2) | 3.73 a (34.9) | 1.71 b (14.4) | 0.59 b (14.9) | 0.60 b (15.4) | 0.931 b (3.1) |
| Papillose | 65 | 0.83 b (33.2) | 5.10 c (25.0) | 1.82 a (33.4) | 0.42 a (22.4) | 0.61 c (27.6) | 0.884 a (2.9) |
| E. coeli-rosa | 20 | 0.45 a (7.5) | 2.95 a (5.5) | 1.60 a (5.8) | 0.65 c (5.9) | 0.63 c (5.6) | 0.939 b,c (1.1) |
| Seed Type or Species | N | C | S |
|---|---|---|---|
| Smooth | 4 | 0.76 b (5.6) | 0.962 a,b (1.5) |
| Rugose | 27 | 0.79 b (3.8) | 0.971 b (0.8) |
| Echinate | 16 | 0.77 b (7.0) | 0.970 b (0.7) |
| Papillose | 4 | 0.64 a (12.0) | 0.946 a (1.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Gómez, J.J.; Rodríguez-Lorenzo, J.L.; Tocino, Á.; Janoušek, B.; Juan, A.; Cervantes, E. The Outline of Seed Silhouettes: A Morphological Approach to Silene (Caryophyllaceae). Plants 2022, 11, 3383. https://doi.org/10.3390/plants11233383
Martín-Gómez JJ, Rodríguez-Lorenzo JL, Tocino Á, Janoušek B, Juan A, Cervantes E. The Outline of Seed Silhouettes: A Morphological Approach to Silene (Caryophyllaceae). Plants. 2022; 11(23):3383. https://doi.org/10.3390/plants11233383
Chicago/Turabian StyleMartín-Gómez, José Javier, José Luis Rodríguez-Lorenzo, Ángel Tocino, Bohuslav Janoušek, Ana Juan, and Emilio Cervantes. 2022. "The Outline of Seed Silhouettes: A Morphological Approach to Silene (Caryophyllaceae)" Plants 11, no. 23: 3383. https://doi.org/10.3390/plants11233383