
Stomata in Close Contact: The Case of Pancratium maritimum L. (Amaryllidaceae)

 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
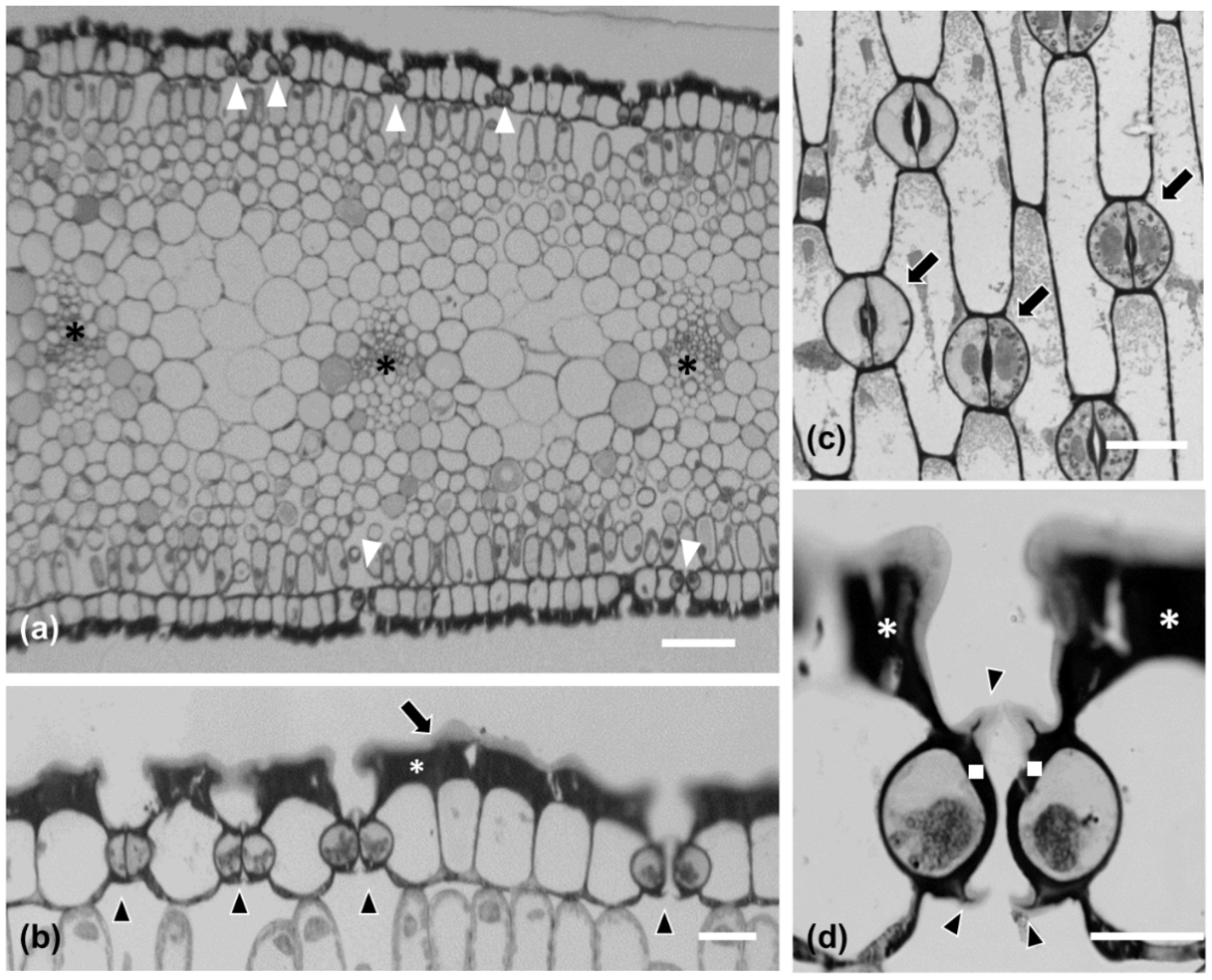
2.1. Morphological and Anatomical Features of Pancratium maritimum Leaf
2.2. Fine Structure of Pancratium maritimum Guard Cells
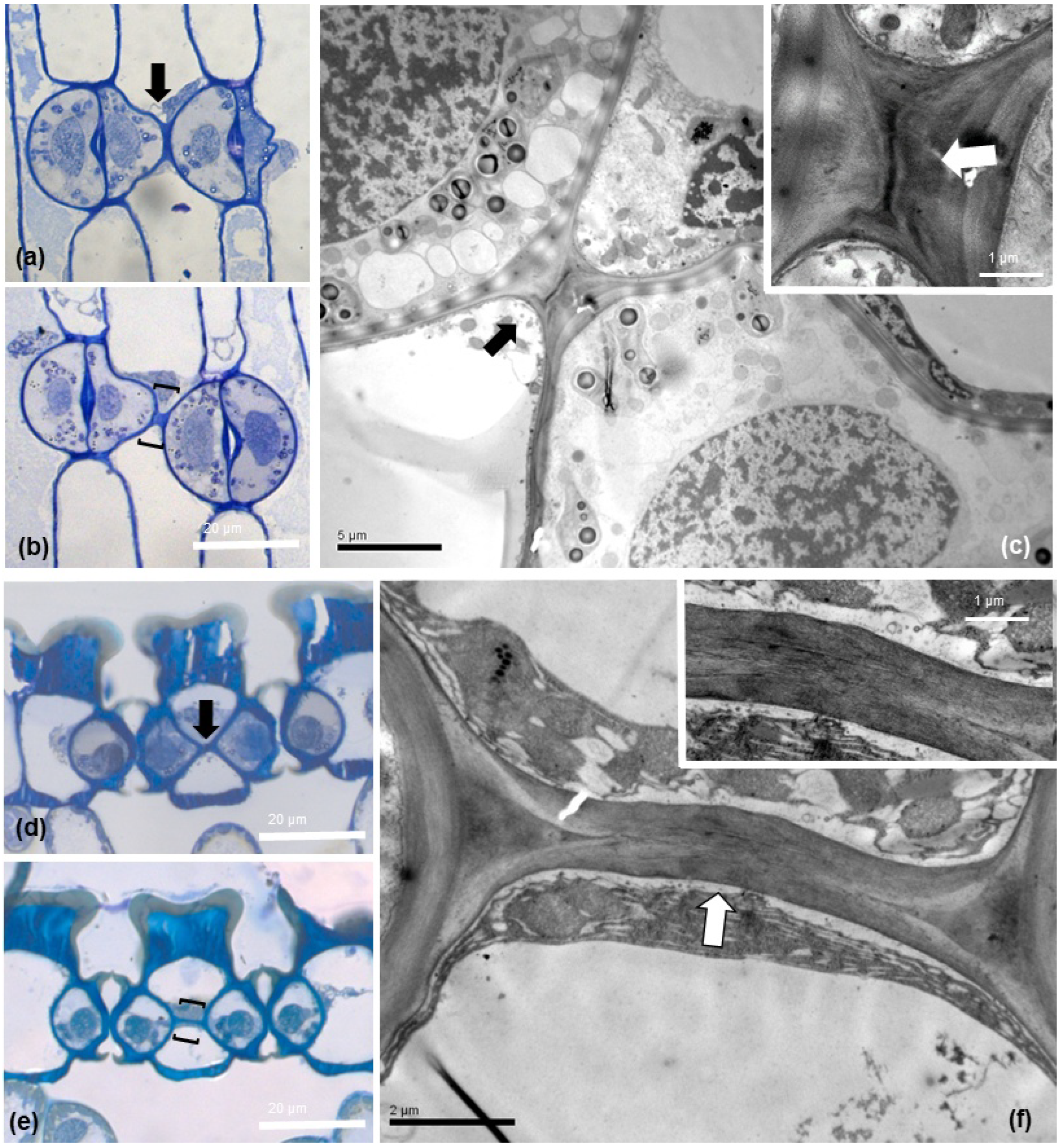
2.3. Neighboring Stomata Are Often Connected by a Strand
2.4. Morphology and Fine Structure of Connecting Strands
2.5. Composition of the Strand Connecting Adjacent Stomata
2.6. Cell Wall Matrix Material Detection at the Connection Strand
2.7. High Crystalline Cellulose Content in the Connecting Strands
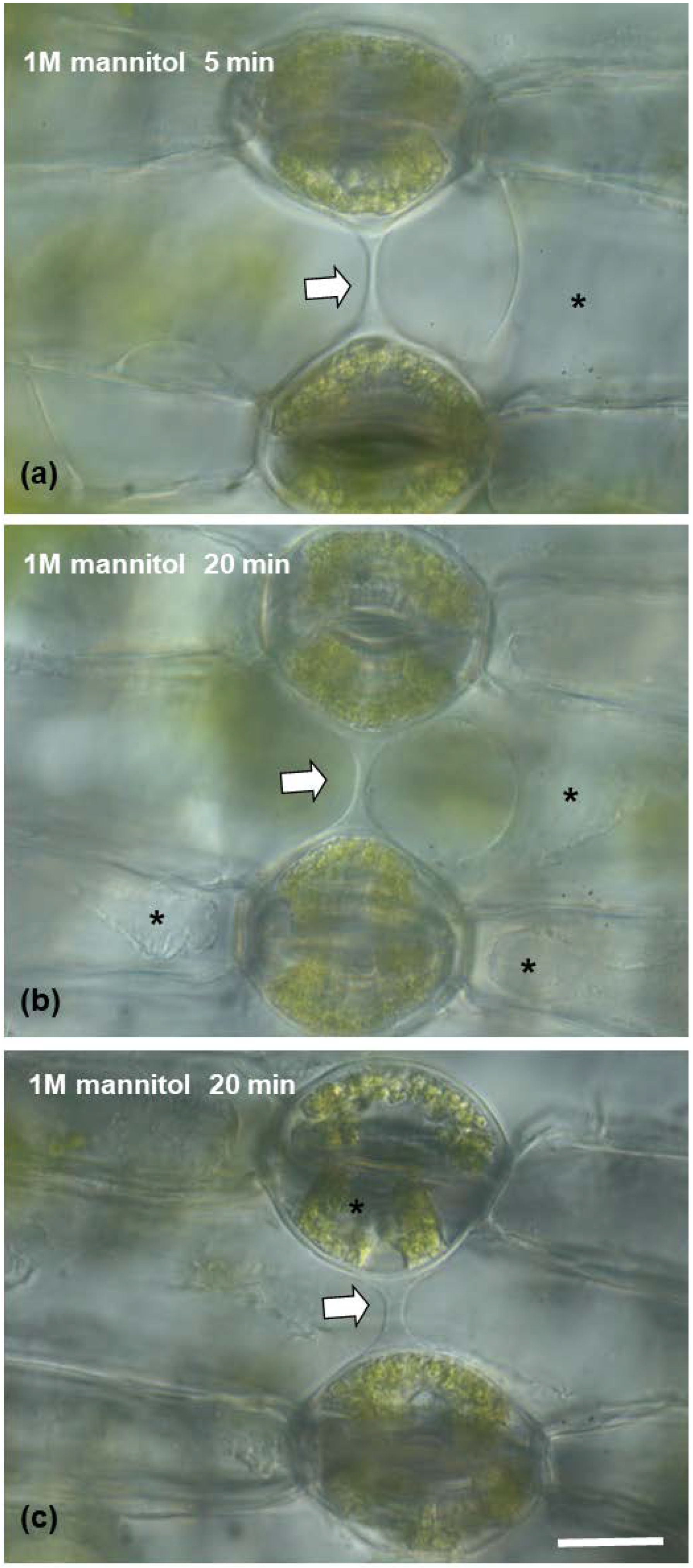
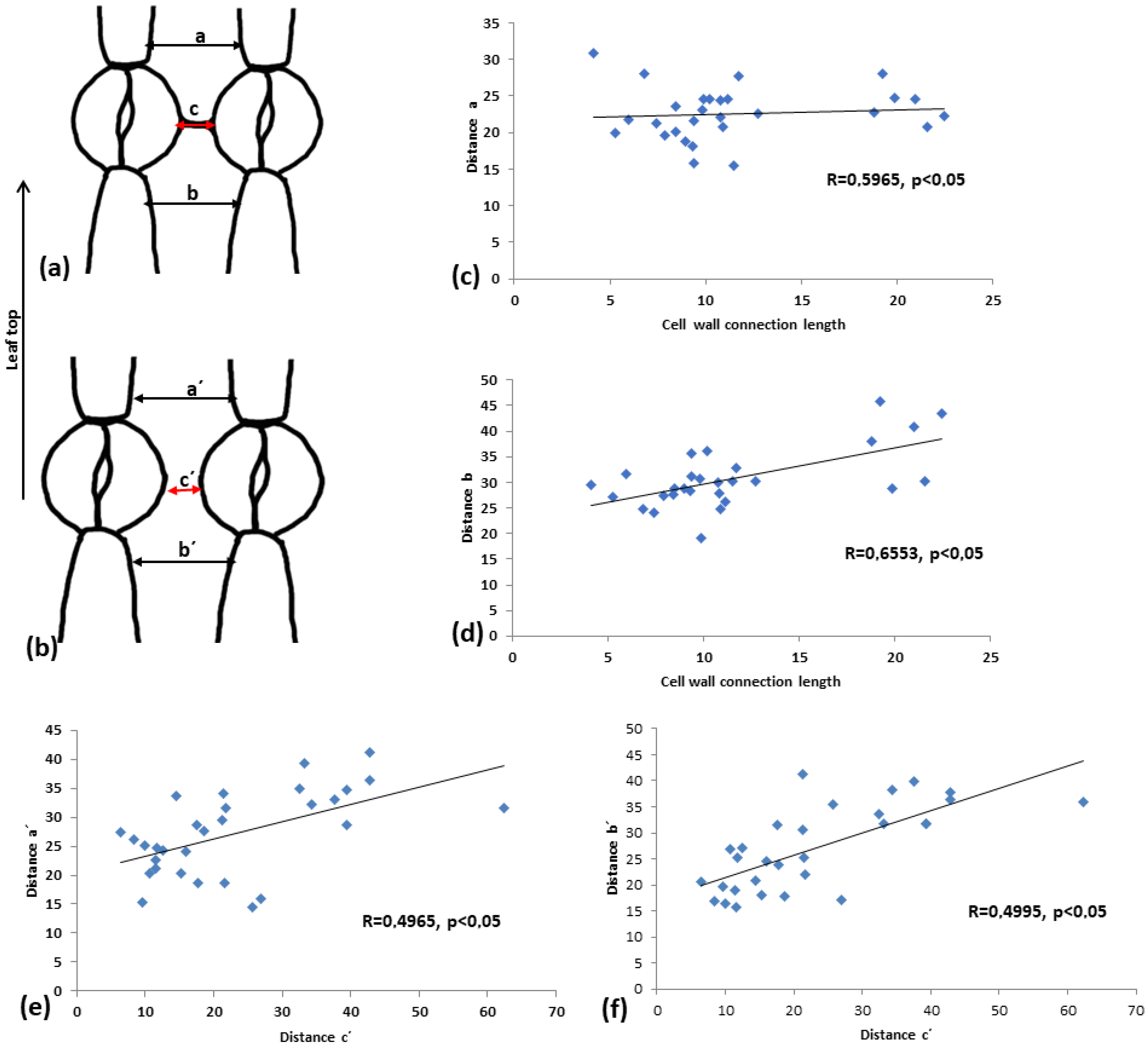
2.8. The Cell Wall Connection Strand Seriously Affects Guard Cell Shape and Pore Aperture
2.9. The Connection Strand Limits the Expansion of the Intervening Pavement Cell
2.10. The Cell Wall Strand Is a Universal Feature Present in Various Subfamilies of Amaryllidaceae
3. Discussion
4. Materials and Methods
4.1. Plant Material
Chemicals
4.2. Methodology
4.2.1. Light Microscopy
Measurement of Stomatal Density in Various Leaf Areas
Measurement of the Morphological Characteristics of the Neighboring Stomata
Stomatal Pore Opening
4.2.2. Experimental Procedures
Aniline Blue Stain
FM 4–64 stain
Tannic Acid–Ferric Chloride Stain
Propidium Iodide Staining
Pectinase Treatment
Mannitol Treatment
Cell Wall Material Immunolocalization
Sample Preparation for Transmission Electron Microscopy (TEM)
Polarized Light Microscopy
Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, J.W.; Harris, B.J.; Hetherington, A.J.; Hurtado-Castano, N.; Brench, R.A.; Casson, S.; Williams, T.A.; Gray, J.E.; Hetherington, A.M. The origin and evolution of stomata. Curr. Biol. 2022, 32, R539–R553. [Google Scholar] [CrossRef] [PubMed]
- Driesen, E.; den Ende, W.; De Proft, M.; Saeys, W. Influence of Environmental Factors Light, CO2, Temperature, and Relative Humidity on Stomatal Opening and Development: A Review. Agronomy 2020, 10, 1975. [Google Scholar] [CrossRef]
- Herrmann, A.; Torii, K.U. Shouting out loud: Signaling modules in the regulation of stomatal development. Plant Physiol. 2021, 185, 765–780. [Google Scholar] [CrossRef]
- McKown, K.H.; Bergmann, D.C. Stomatal development in the grasses: Lessons from models and crops (and crop models). New Phytol. 2020, 227, 1636–1648. [Google Scholar] [CrossRef] [Green Version]
- Nunes, T.D.G.; Zhang, D.; Raissig, M.T. Form, development and function of grass stomata. Plant J. 2020, 101, 780–799. [Google Scholar] [CrossRef] [PubMed]
- Chater, C.C.C.; Caine, R.S.; Fleming, A.J.; Gray, J.E. Origins and evolution of stomatal development. Plant Physiol. 2017, 174, 624–638. [Google Scholar] [CrossRef] [Green Version]
- Han, S.K.; Torii, K.U. Lineage-specific stem cells, signals and asymmetries during stomatal development. Development 2016, 143, 1259–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spiegelhalder, R.P.; Raissig, M.T. Morphology made for movement: Formation of diverse stomatal guard cells. Curr. Opin. Plant Biol. 2021, 63, 102090. [Google Scholar] [CrossRef]
- Torii, K.U. Stomatal development in the context of epidermal tissues. Ann. Bot. 2021, 128, 137–148. [Google Scholar] [CrossRef]
- Apostolakos, P.; Galatis, B. Induction, polarity and spatial control of cytokinesis in some abnormal subsidiary cell mother cells of Zea mays. Protoplasma 1987, 140, 26–42. [Google Scholar] [CrossRef]
- Serna, L.; Fenoll, C. Tracing the ontogeny of stomatal clusters in Arabidopsis with molecular markers. Plant J. 1997, 12, 747–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papanatsiou, M.; Amtmann, A.; Blatt, M.R. Stomatal clustering in Begonia associates with the kinetics of leaf gaseous exchange and influences water use efficiency. J. Exp. Bot. 2017, 68, 2309–2315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, P.; Or, D. Effects of stomata clustering on leaf gas exchange. New Phytol. 2015, 207, 1015–1025. [Google Scholar] [CrossRef]
- Meerow, A.W.; Snijman, D.A. Amaryllidaceae BT. In Flowering Plants Monocotyledons: Lilianae (Except Orchidaceae); Kubitzki, K., Ed.; Springer: Berlin/Heidelberg, Germany, 1998; pp. 83–110. ISBN 978-3-662-03533-7. [Google Scholar]
- De Castro, O.; Brullo, S.; Colombo, P.; Jury, S.; De Luca, P.; Di Maio, A. Phylogenetic and biogeographical inferences for Pancratium (Amaryllidaceae), with an emphasis on the Mediterranean species based on plastid sequence data. Bot. J. Linn. Soc. 2012, 170, 12–28. [Google Scholar] [CrossRef] [Green Version]
- Zahreddine, H.; Clubbe, C.; Baalbaki, R.; Ghalayini, A.; Talhouk, S.N. Status of native species in threatened Mediterranean habitats: The case of Pancratium maritimum L. (sea daffodil) in Lebanon. Biol. Conserv. 2004, 120, 11–18. [Google Scholar] [CrossRef]
- Grassi, F.; Cazzaniga, E.; Minuto, L.; Peccenini, S.; Barberis, G.; Basso, B. Evaluation of biodiversity and conservation strategies in Pancratium maritimum L. for the NorthernTyrrhenian Sea. Biodivers. Conserv. 2005, 14, 2159–2169. [Google Scholar] [CrossRef]
- Carfagna, S.; Salbitani, G.; Innangi, M.; Menale, B.; De Castro, O.; Di Martino, C.; Crawford, T.W. Simultaneous Biochemical and Physiological Responses of the Roots and Leaves of Pancratium maritimum (Amaryllidaceae) to Mild Salt Stress. Plants 2021, 10, 345. [Google Scholar] [CrossRef]
- Pouris, J.; Meletiou-Christou, M.-S.; Chimona, C.; Rhizopoulou, S. Seasonal Functional Partitioning of Carbohydrates and Proline among Plant Parts of the Sand Daffodil. Agronomy 2020, 10, 539. [Google Scholar] [CrossRef] [Green Version]
- Perrone, R.; Salmeri, C.; Brullo, S.; Colombo, P.; De Castro, O. What do leaf anatomy and micro-morphology tell us about the psammophilous Pancratium maritimum L. (Amaryllidaceae) in response to sand dune conditions? Flora–Morphol. Distrib. Funct. Ecol. Plants 2015, 213, 20–31. [Google Scholar] [CrossRef]
- Rudall, P.J.; Chen, E.D.; Cullen, E. Evolution and development of monocot stomata. Am. J. Bot. 2017, 104, 1122–1141. [Google Scholar] [CrossRef]
- Shah, G.L.; Gopal, B.V. Structure and Development of Stomata on the Vegetative and Floral Organs of Some Amaryllidaceae. Ann. Bot. 1970, 34, 737–749. [Google Scholar] [CrossRef]
- Arroyo, S.C.; Cutler, D.F. Evolutionary and Taxonomic Aspects of the Internal Morphology in Amaryllidaceae from South America and Southern Africa. Kew Bull. 1984, 39, 467–498. [Google Scholar] [CrossRef]
- Awasthi, D.K.; Kumar, V.; Rawat, R. Stomatal studies in Amaryllidaceae with special reference to stomatal abnormalities. Proc. Plant Sci. 1984, 93, 629–633. [Google Scholar] [CrossRef]
- Hashemloian, B.D.; Azimi, A.A. Abnormal and cytoplasmic connection of guard cells of stomata of leafs of six species of the monocots. J. Plant Sci. 2014, 2, 334–338. [Google Scholar] [CrossRef]
- Du, J.; Anderson, C.T.; Xiao, C. Dynamics of pectic homogalacturonan in cellular morphogenesis and adhesion, wall integrity sensing and plant development. Nat. Plants 2022, 8, 332–340. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wang, M.; Song, X.; Zhou, G.; Kong, Y. The evolving views of the simplest pectic polysaccharides: Homogalacturonan. Plant Cell Rep. 2022, 41, 2111–2123. [Google Scholar] [CrossRef]
- Novaković, L.; Guo, T.; Bacic, A.; Sampathkumar, A.; Johnson, K.L. Hitting the Wall—Sensing and Signaling Pathways Involved in Plant Cell Wall Remodeling in Response to Abiotic Stress. Plants 2018, 7, 89. [Google Scholar] [CrossRef] [Green Version]
- De Lorenzo, G.; Ferrari, S.; Giovannoni, M.; Mattei, B.; Cervone, F. Cell wall traits that influence plant development, immunity, and bioconversion. Plant J. 2019, 97, 134–147. [Google Scholar] [CrossRef]
- Yang, M.; Sack, F.D. The too many mouths and four lips mutations affect stomatal production in Arabidopsis. Plant Cell 1995, 7, 2227–2239. [Google Scholar]
- Casson, S.; Gray, J.E. Influence of environmental factors on stomatal development. New Phytol. 2008, 178, 9–23. [Google Scholar] [CrossRef]
- Akita, K.; Higaki, T. An Induction System for Clustered Stomata by Sugar Solution Immersion Treatment in Arabidopsis thaliana Seedlings. J. Vis. Exp. 2019, 144, e58951. [Google Scholar] [CrossRef]
- Liu, C.; Li, Y.; Xu, L.; Li, M.; Wang, J.; Yan, P.; He, N. Stomatal arrangement pattern: A new direction to explore plant adaptation and evolution. Front. Plant Sci. 2021, 12, 655255. [Google Scholar] [CrossRef] [PubMed]
- Tsakali, A.; Asitzoglou, I.-C.; Basdeki, V.; Podia, V.; Adamakis, I.-D.S.; Giannoutsou, E.; Haralampidis, K. The Role of PME2 and PME3 in Arabidopsis Stomatal Development and Morphology. Biol. Life Sci. Forum 2022, 11, 36. [Google Scholar]
- Paraskevopoulou, D.; Anezakis, N.; Giannoutsou, E.; Sotiriou, P.; Adamakis, I.-D.S. The Stomata of the Katanin Mutants, fra2, lue1 and bot1. Biol. Life Sci. Forum 2021, 4, 30. [Google Scholar]
- Hara, K.; Kajita, R.; Torii, K.U.; Bergmann, D.C.; Kakimoto, T. The secretory peptide gene EPF1 enforces the stomatal one-cell-spacing rule. Genes Dev. 2007, 21, 1720–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.; Kuan, C.; Guo, Z.; Yang, H.; Chung, K.; Ho, C.K. Stomatal clustering in Begonia improves water use efficiency by modulating stomatal movement and leaf structure. Plant-Environ. Interact. 2022, 3, 141–154. [Google Scholar] [CrossRef]
- Pouris, J.; Rhizopoulou, S. On Pancratium maritimum (sea daffodil, sea lily, sand lily). Hortic. Int. J. 2018, 2, 116–118. [Google Scholar]
- Giannopoulos, K.; Tan, K.; Vold, G. Contributions to the bulb flora of Ilias (NW Peloponnese, Greece): Amaryllidaceae, Araceaeand Aristolochiaceae. Phytol. Balcan. 2021, 27, 97–106. [Google Scholar]
- Wang, Y.; Chen, X.; Xiang, C. Stomatal density and bio-water saving. J. Integr. Plant Biol. 2007, 49, 1435–1444. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Relaxation in a high-stress environment: The molecular bases of extensible cell walls and cell enlargement. Plant Cell 1997, 9, 1031. [Google Scholar] [CrossRef] [Green Version]
- Allen, G.J.; Chu, S.P.; Harrington, C.L.; Schumacher, K.; Hoffmann, T.; Tang, Y.Y.; Grill, E.; Schroeder, J.I. A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature 2001, 411, 1053–1057. [Google Scholar] [CrossRef] [PubMed]
- Sachs, T. Stomata as an example of meristemoid development. In Pattern Formation in Plant Tissues; Cambridge University Press: Cambridge, UK, 1991; pp. 101–117. [Google Scholar]
- Cheng, R.-Y.; Xie, D.-F.; Zhang, X.-Y.; Fu, X.; He, X.-J.; Zhou, S.-D. Comparative Plastome Analysis of Three Amaryllidaceae Subfamilies: Insights into Variation of Genome Characteristics, Phylogeny, and Adaptive Evolution. BioMed Res. Int. 2022, 2022, 3909596. [Google Scholar] [CrossRef] [PubMed]
- Zamil, M.S.; Geitmann, A. The middle lamella—More than a glue. Phys. Biol. 2017, 14, 15004. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Chen, Y.; Kandemir, B.; Yi, H.; Wang, J.Z.; Puri, V.M.; Anderson, C.T. Balancing strength and flexibility: How the synthesis, organization, and modification of guard cell walls govern stomatal development and dynamics. Front. Plant Sci. 2018, 9, 1202. [Google Scholar] [CrossRef] [Green Version]
- Apostolakos, P.; Giannoutsou, E.; Galatis, B. Callose: A multifunctional (1,3)-β–D-glucan involved in morphogenesis and function of angiosperm stomata. J. Biol. Res.-Thessaloniki 2021, 28, 17. [Google Scholar] [CrossRef]
- Giannoutsou, E.; Apostolakos, P.; Galatis, B. Spatio-temporal diversification of the cell wall matrix materials in the developing stomatal complexes of Zea mays. Planta 2016, 244, 1125–1143. [Google Scholar] [CrossRef]
- Apostolakos, P.; Galatis, B. Microtubule and actin filament organization during stomatal morphogenesis in the fern Asplenium nidus. II. Guard cells. New Phytol. 1999, 141, 209–223. [Google Scholar] [CrossRef]
- Rigal, A.; Doyle, S.M.; Robert, S. Live cell imaging of FM4-64, a tool for tracing the endocytic pathways in Arabidopsis root cells. In Plant Cell Expansion; Springer: Berlin/Heidelberg, Germany, 2015; pp. 93–103. [Google Scholar]
- Hornatowska, J. Visualisation of Pectins and Proteins by Microscopy; STFI-Packforsk Report; STFI-Packforsk: Stockholm, Sweden, 2005; 22p. [Google Scholar]
- Bidhendi, A.J.; Chebli, Y.; Geitmann, A. Fluorescence visualization of cellulose and pectin in the primary plant cell wall. J. Microsc. 2020, 278, 164–181. [Google Scholar] [CrossRef]
- Komis, G.; Apostolakos, P.; Galatis, B. Altered patterns of tubulin polymerization in dividing leaf cells of Chlorophyton comosum after a hyperosmotic treatment. New Phytol. 2001, 149, 193–207. [Google Scholar] [CrossRef]
- Apostolakos, P.; Livanos, P.; Nikolakopoulou, T.L.; Galatis, B. The role of callose in guard-cell wall differentiation and stomatal pore formation in the fern Asplenium nidus. Ann. Bot. 2009, 104, 1373–1387. [Google Scholar] [CrossRef] [Green Version]
- Giannoutsou, E.; Sotiriou, P.; Nikolakopoulou, T.L.; Galatis, B.; Apostolakos, P. Callose and homogalacturonan epitope distribution in stomatal complexes of Zea mays and Vigna sinensis. Protoplasma. 2019, 257, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Verhertbruggen, Y.; Marcus, S.E.; Haeger, A.; Ordaz-Ortiz, J.J.; Knox, J.P. An extended set of monoclonal antibodies to pectic homogalacturonan. Carbohydr. Res. 2009, 344, 1858–1862. [Google Scholar] [CrossRef] [PubMed]
- Liners, F.; Letesson, J.-J.; Didembourg, C.; Van Cutsem, P. Monoclonal antibodies against pectin: Recognition of a conformation induced by calcium. Plant Physiol. 1989, 91, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Shtein, I.; Shelef, Y.; Marom, Z.; Zelinger, E.; Schwartz, A.; Popper, Z.A.; Bar-On, B.; Harpaz-Saad, S. Stomatal cell wall composition: Distinctive structural patterns associated with different phylogenetic groups. Ann. Bot. 2017, 119, 1021–1033. [Google Scholar] [CrossRef] [Green Version]
- Shtein, I.; Popper, Z.A.; Harpaz-Saad, S. Permanently open stomata of aquatic angiosperms display modified cellulose crystallinity patterns. Plant Signal. Behav. 2017, 12, 1021–1033. [Google Scholar] [CrossRef] [PubMed]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saridis, P.; Georgiadou, X.; Shtein, I.; Pouris, J.; Panteris, E.; Rhizopoulou, S.; Constantinidis, T.; Giannoutsou, E.; Adamakis, I.-D.S. Stomata in Close Contact: The Case of Pancratium maritimum L. (Amaryllidaceae). Plants 2022, 11, 3377. https://doi.org/10.3390/plants11233377
Saridis P, Georgiadou X, Shtein I, Pouris J, Panteris E, Rhizopoulou S, Constantinidis T, Giannoutsou E, Adamakis I-DS. Stomata in Close Contact: The Case of Pancratium maritimum L. (Amaryllidaceae). Plants. 2022; 11(23):3377. https://doi.org/10.3390/plants11233377
Chicago/Turabian StyleSaridis, Pavlos, Xenia Georgiadou, Ilana Shtein, John Pouris, Emmanuel Panteris, Sophia Rhizopoulou, Theophanis Constantinidis, Eleni Giannoutsou, and Ioannis-Dimosthenis S. Adamakis. 2022. "Stomata in Close Contact: The Case of Pancratium maritimum L. (Amaryllidaceae)" Plants 11, no. 23: 3377. https://doi.org/10.3390/plants11233377