

The Effects of Microbiota on the Herbivory Resistance of the Giant Duckweed Are Plant Genotype-Dependent

{kind=link}
{kind=link}
Abstract
:1. Introduction
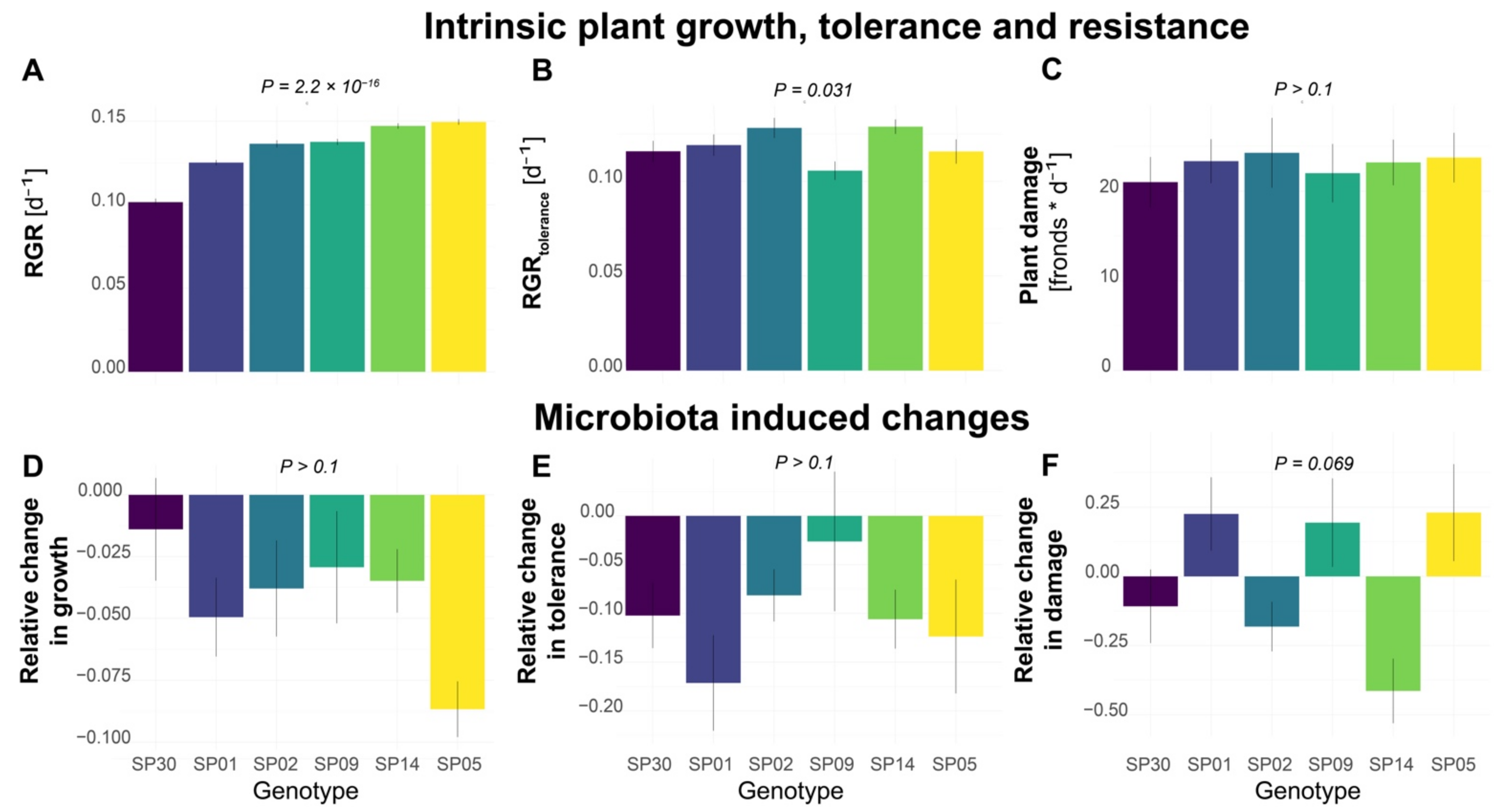
2. Results
2.1. Different Genotypes Varied in Intrinsic Growth Rate, but Not Resistance to Herbivory by Snails
2.2. Microbiota Inoculation Altered Tolerance and Resistance to Herbivory in a Genotype-Specific Manner
3. Discussion
4. Methods
4.1. Duckweed Growth with and without Microbiota-Inoculation
4.2. Microbe Inoculum
4.3. Herbivore Assay
4.4. Statistics and Data Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Statements
References
- Müller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The plant microbiota: Systems-level insights and perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Shikano, I.; Rosa, C.; Tan, C.-W.; Felton, G.W. Tritrophic interactions: Microbe-mediated plant effects on insect herbivores. Annu. Rev. Phytopathol. 2017, 55, 313–331. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; van der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friman, J.; Karssemeijer, P.N.; Haller, J.; de Kreek, K.; van Loon, J.J.A.; Dicke, M. Shoot and root insect herbivory change the plant rhizosphere microbiome and affects cabbage–insect interactions through plant–soil feedback. New Phytol. 2021, 232, 2475–2490. [Google Scholar] [CrossRef]
- Toyama, T.; Yu, N.; Kumada, H.; Sei, K.; Ike, M.; Fujita, M. Accelerated aromatic compounds degradation in aquatic environment by use of interaction between Spirodela polyrrhiza and bacteria in its rhizosphere. J. Biosci. Bioeng. 2006, 101, 346–353. [Google Scholar] [CrossRef] [Green Version]
- Hoang, H.; Yu, N.; Toyama, T.; Inoue, D.; Sei, K.; Ike, M. Accelerated degradation of a variety of aromatic compounds by Spirodela polyrrhiza-bacterial associations and contribution of root exudates released from S. polyrrhiza. J. Environ. Sci. 2010, 22, 494–499. [Google Scholar] [CrossRef] [PubMed]
- Toyama, T.; Mori, K.; Tanaka, Y.; Ike, M.; Morikawa, M. Growth promotion of giant duckweed Spirodela polyrhiza (Lemnaceae) by Ensifer sp. SP4 through enhancement of nitrogen metabolism and photosynthesis. Mol. Plant Microbe. Interact. 2021, 35, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Toporowska, M. Degradation of three microcystin variants in the presence of the macrophyte Spirodela polyrhiza and the associated microbial communities. Int. J. Environ. Res. Public Health 2022, 19, 6086. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, A.M.; Laurich, J.; Lash, E.; Frederickson, M.E. Mutualistic outcomes across plant populations, microbes, and environments in the duckweed Lemna minor. Microb. Ecol. 2020, 80, 384–397. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, H.; Kuroda, M.; Morikawa, M.; Ike, M. Evaluation of environmental bacterial communities as a factor affecting the growth of duckweed Lemna minor. Biotechnol. Biofuels 2017, 10, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roijackers, R.; Szabó, S.; Scheffer, M. Experimental analysis of the competition between algae and duckweed. Arch. Hydrobiol. 2004, 160, 401–412. [Google Scholar] [CrossRef]
- Appenroth, K.-J.; Nickel, G. Turion formation in Spirodela polyrhiza: The environmental signals that induce the developmental process in nature. Physiol. Plant. 2010, 138, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.; Xu, J.; Acosta, K.; Poulev, A.; Lebeis, S.L.; Lam, E. Bacterial production of indole related compounds reveals their role in association between duckweeds and endophytes. Front. Chem. 2018, 6, 265. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Sharma, S.S. Nutrient deficiency-dependent anthocyanin development in Spirodela polyrhiza L. Schleid. Biol. Plant. 1999, 42, 621–624. [Google Scholar] [CrossRef]
- Bultman, T.L.; Borowicz, K.L.; Schneble, R.M.; Coudron, T.A.; Bush, L.P. Effect of a fungal endophyte on the growth and survival of two Euplectrus parasitoids. Oikos 1997, 78, 170–176. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Bortfeld-Miller, M.; Ackermann, M.; Vorholt, J.A. A synthetic community approach reveals plant genotypes affecting the phyllosphere microbiota. PLoS Genet. 2014, 10, e1004283. [Google Scholar] [CrossRef] [Green Version]
- Acosta, K.; Xu, J.; Gilbert, S.; Denison, E.; Brinkman, T.; Lebeis, S.; Lam, E. Duckweed hosts a taxonomically similar bacterial assemblage as the terrestrial leaf microbiome. PLoS ONE 2020, 15, e0228560. [Google Scholar] [CrossRef] [Green Version]
- Malacrinò, A.; Böttner, L.; Nouere, S.; Huber, M.; Schäfer, M.; Xu, S. Induced responses contribute to rapid plant adaptation to herbivory. bioRxiv 2022. [Google Scholar] [CrossRef]
- Appenroth, K. Media for in vitro-cultivation of duckweed. Duckweed Forum 2015, 3, 180–186. [Google Scholar]
- Ziegler, P.; Adelmann, K.; Zimmer, S.; Schmidt, C.; Appenroth, K.-J. Relative in vitro growth rates of duckweeds (Lemnaceae)–the most rapidly growing higher plants. Plant Biol. 2015, 17, 33–41. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 1 October 2022).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schäfer, M.; Xu, S. The Effects of Microbiota on the Herbivory Resistance of the Giant Duckweed Are Plant Genotype-Dependent. Plants 2022, 11, 3317. https://doi.org/10.3390/plants11233317
Schäfer M, Xu S. The Effects of Microbiota on the Herbivory Resistance of the Giant Duckweed Are Plant Genotype-Dependent. Plants. 2022; 11(23):3317. https://doi.org/10.3390/plants11233317
Chicago/Turabian StyleSchäfer, Martin, and Shuqing Xu. 2022. "The Effects of Microbiota on the Herbivory Resistance of the Giant Duckweed Are Plant Genotype-Dependent" Plants 11, no. 23: 3317. https://doi.org/10.3390/plants11233317