

Quantitative Real-Time PCR Based on SYBR Green Technology for the Identification of Philaenus italosignus Drosopoulos & Remane (Hemiptera Aphrophoridae)
 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. DNA Extraction
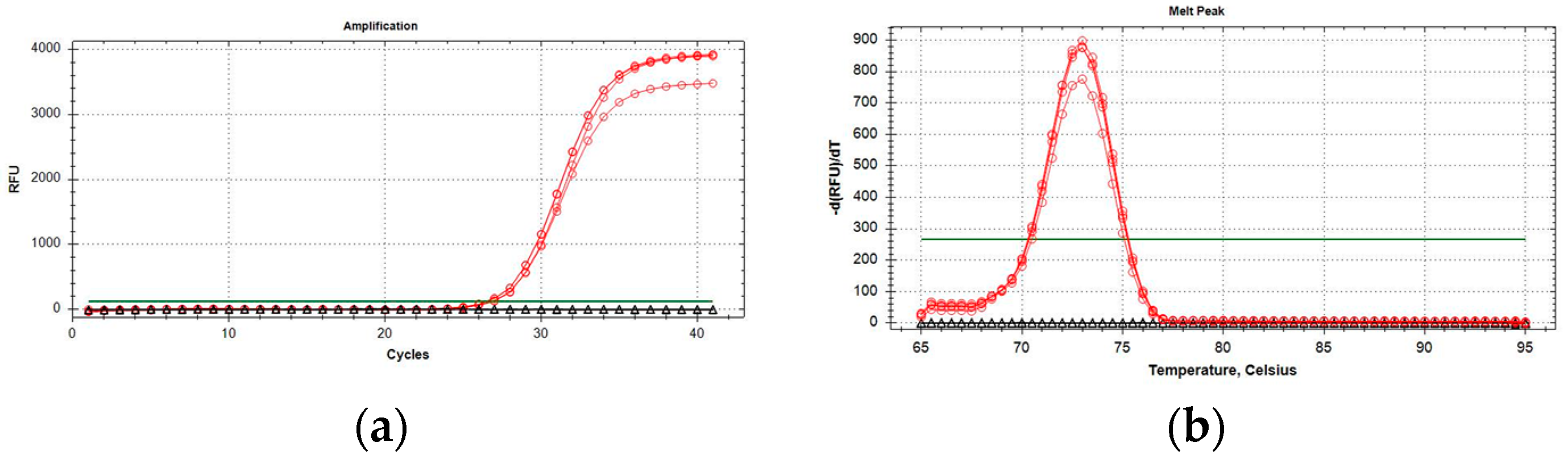
2.2. Development and Optimization of P. italosignus-Specific qPCR
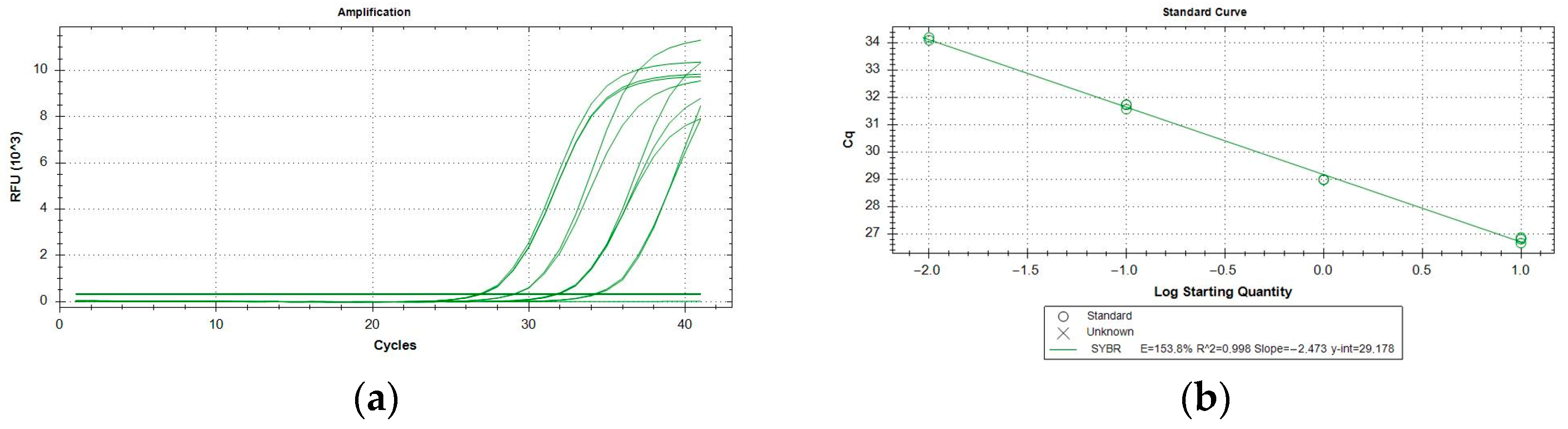
2.3. Performance Characteristics
2.4. Field Protocol Validation
3. Discussion
4. Material and Methods
4.1. Insect Samples for the Setup of the Molecular Diagnostic Protocol
4.2. Heterozygosity of the Elongation Factor-1α Gene in Philaenus italosignus and P. spumarius
4.3. DNA Extraction
4.4. Design of P. italosignus Primers
4.5. QPCR Optimization
4.6. Performance Characteristics
4.7. Insect Sampling for Protocol Validation
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cavalieri, V.; Altamura, G.; Fumarola, G.; di Carolo, M.; Saponari, M.; Cornara, D.; Bosco, D.; Dongiovanni, C. Transmission of Xylella fastidiosa Subspecies Pauca Sequence Type 53 by Different Insect Species. Insects 2019, 10, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drosopoulos, S.; Remane, R. Biogeographic studies on the spittlebug Philaenus signatus Melichar, 1896 species group (Hemiptera: Aphrophoridae) with the description of two new allopatric species. Ann. Soc. Entomol. Fr. N.S. 2000, 36, 269–277. [Google Scholar]
- Panzavolta, T.; Bracalini, M.; Croci, F.; Ghelardini, L.; Luti, S.; Campigli, S.; Goti, E.; Marchi, R.; Tiberi, R.; Marchi, G. Philaenus italosignus a potential vector of Xylella fastidiosa: Occurrence of the spittlebug on olive trees in Tuscany (Italy). Bull. Insectol. 2019, 72, 317–320. [Google Scholar]
- Marchi, G.; Rizzo, D.; Ranaldi, F.; Ghelardini, L.; Ricciolini, M.; Scarpelli, I.; Drosera, L.; Goti, E.; Capretti, P.; Surico, G. First detection of Xylella fastidiosa subsp. multiplex DNA in Tuscany (Italy). Phytopathol. Mediterr. 2018, 57, 363–364. [Google Scholar] [CrossRef]
- Saponari, M.; D’Attoma, G.; Kubaa, R.A.; Loconsole, G.; Altamura, G.; Zicca, S.; Rizzo, D.; Boscia, D. A new variant of Xylella fastidiosa subspecies multiplex detected in different host plants in the recently emerged outbreak in the region of Tuscany, Italy. Eur. J. Plant Pathol. 2019, 154, 1195–1200. [Google Scholar] [CrossRef] [Green Version]
- EPPO (European and Mediterranean Plant Protection Organization). PM 7/141 (1) Philaenus spumarius, Philaenus italosignus and Neophilaenus campestris. EPPO Bull. 2020, 50, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Lahbib, N.; Picciotti, U.; Boukhris-Bouhachem, S.; Garganese, F.; Porcelli, F. Morphs of Philaenus species, candidate Xylella fastidiosa vectors. Bull. Insectol. 2022, 75, 197–209. [Google Scholar]
- Seabra, S.; Pina-Martins, F.; Marabuto, E.; Yurtsever, S.; Halkka, O.; Quartau, J.; Paulo, O. Molecular phylogeny and DNA barcoding in the meadow-spittlebug Philaenus spumarius (Hemiptera, Cercopidae) and its related species. Mol. Phylogenetics Evol. 2010, 56, 462–467. [Google Scholar] [CrossRef]
- Ioos, R.; Fourrier, C.; Iancu, G.; Gordon, T.R. Sensitive Detection of Fusarium circinatum in Pine Seed by Combining an Enrichment Procedure with a Real-Time Polymerase Chain Reaction Using Dual-Labeled Probe Chemistry. Phytopathology 2009, 99, 582–590. [Google Scholar] [CrossRef] [Green Version]
- Di Serio, F.; Bodino, N.; Cavalieri, V.; Demichelis, S.; Di Carolo, M.; Dongiovanni, C.; Fumarola, G.; Gilioli, G.; Guerrieri, E.; Picciotti, U.; et al. Collection of data and information on biology and control of vectors of Xylella fastidiosa. EFSA Support. Publ. 2019, 16, EN-1628. [Google Scholar] [CrossRef]
- Liccardo, A.; Fierro, A.; Garganese, F.; Picciotti, U.; Porcelli, F. A biological control model to manage the vector and the infection of Xylella fastidiosa on olive trees. PLoS ONE 2020, 15, e0232363. [Google Scholar] [CrossRef] [PubMed]
- Picciotti, U.; Lahbib, N.; Sefa, V.; Porcelli, F.; Garganese, F. Aphrophoridae Role in Xylella fastidiosa subsp. pauca ST53 Invasion in Southern Italy. Pathogens 2021, 10, 1035. [Google Scholar] [CrossRef]
- Lantero, E.; Matallanas, B.; Pascual, S.; Callejas, C. PCR Species-Specific Primers for Molecular Gut Content Analysis to Determine the Contribution of Generalist Predators to the Biological Control of the Vector of Xylella fastidiosa. Sustainability 2018, 10, 2207. [Google Scholar] [CrossRef]
- Rizzo, D.; Da Lio, D.; Bartolini, L.; Salemi, C.; Pennacchio, F.; Rapisarda, C.; Rossi, E. The Rapid Identification of Anoplophora chinensis (Coleoptera: Cerambycidae) from Adult, Larval, and Frass Samples Using TaqMan Probe Assay. J. Econ. Èntomol. 2021, 114, 2229–2235. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.; Ochoa-Corona, F.M.; Opit, G.P.; Li, Z.-H.; Kučerová, Z.; Stejskal, V.; Yang, Q.-Q. PCR and isothermal-based molecular identification of the stored-product psocid pest Lepinotus reticulatus (Psocoptera: Trogiidae). J. Stored Prod. Res. 2012, 49, 184–188. [Google Scholar] [CrossRef]
- Rizzo, D.; Da Lio, D.; Bartolini, L.; Cappellini, G.; Bruscoli, T.; Salemi, C.; Aronadio, A.; Del Nista, D.; Pennacchio, F.; Boersma, N.; et al. Development of Three Molecular Diagnostic Tools for the Identification of the False Codling Moth (Lepidoptera: Tortricidae). J. Econ. Èntomol. 2021, 114, 1796–1807. [Google Scholar] [CrossRef]
- Rodrigues, A.S.B.; Silva, S.E.; Marabuto, E.; Silva, D.N.; Wilson, M.R.; Thompson, V.; Yurtsever, S.; Halkka, A.; Borges, P.A.V.; Quartau, J.A.; et al. New Mitochondrial and Nuclear Evidences Support Recent Demographic Expansion and an Atypical Phylogeographic Pattern in the Spittlebug Philaenus spumarius (Hemiptera, Aphrophoridae). PLoS ONE 2014, 9, e98375. [Google Scholar] [CrossRef] [Green Version]
- Gargani, E.; Benvenuti, C.; Marianelli, L.; Roversi, P.F.; Ricciolini, M.; Scarpelli, I.; Sacchetti, P.; Nencioni, A.; Rizzo, D.; Strangi, A.; et al. A five-year survey in Tuscany (Italy) and detection of Xylella fastidiosa subspecies multiplex in potential insect vectors, collected in Monte Argentario. J. Zool. 2021, 104, 75–88. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Marchi, G.; Cinelli, T.; Rizzo, D.; Stefani, L.; Goti, E.; DELLA Bartola, M.; Luvisi, A.; Panattoni, A.; Materazzi, A. Occurrence of different phytoplasma infections in wild herbaceous dicots growing in vineyards affected by bois noir in Tuscany (Italy). Phytopathol. Mediterr. 2015, 54, 504–515. [Google Scholar] [CrossRef]
- Rizzo, D.; Taddei, A.; Da Lio, D.; Nugnes, F.; Barra, E.; Stefani, L.; Bartolini, L.; Griffo, R.; Spigno, P.; Cozzolino, L.; et al. Identification of the Red-Necked Longhorn Beetle Aromia bungii (Faldermann, 1835) (Coleoptera: Cerambycidae) with Real-Time PCR on Frass. Sustainability 2020, 12, 6041. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. A simple method to control over-alignment in the MAFFT multiple sequence alignment program. Bioinformatics 2016, 32, 1933–1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- PM 7/98 (5) Specific requirements for laboratories preparing accreditation for a plant pest diagnostic activity. EPPO Bull. 2021, 51, 468–498. [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Adults (n * = 12) |
|---|---|
| DNA conc (ng/µL) ± SD | 78.00 ± 3.20 |
| (A260/280) | 1.85 ± 0.24 |
| Cq (18S) | 19.75 ± 1.80 |
| Dilutions 1:5 (ng/µL) | Adult |
|---|---|
| Cq Means ± SD | |
| 10 | 27.15 ± 0.07 |
| 2.0 | 29.47 ± 0.29 |
| 0.4 | 32.09 ± 0.06 |
| 0.08 | 34.81 ± 0.55 |
| 0.016 | 35.40 ± 0.14 |
| 3.2 × 10−3 | - |
| 6.4 × 10−4 | - |
| 1.28 × 10−4 | - |
| 2.56 × 10−5 | - |
| N° | Repeatability | Reproducibility | ||
|---|---|---|---|---|
| Assay 1 (Average ± SD) | Assay 2 (Average ± SD) | Assay 1 (Average ± SD) | Assay 2 (Average ± SD) | |
| 1 | 26.88 ± 0.15 | 26.94 ± 0.24 | 26.58 ± 0.21 | 26.98 ± 0.28 |
| 2 | 26.63 ± 0.07 | 26.57 ± 0.01 | 26.88 ± 0.05 | 26.80 ± 0.06 |
| 3 | 26.60 ± 0.14 | 26.70 ± 0.13 | 26.49 ± 0.24 | 26.52 ± 0.24 |
| 4 | 26.74 ± 0.16 | 26.81 ± 0.16 | 26.45 ± 0.06 | 26.48 ± 0.06 |
| 5 | 26.71 ± 0.19 | 26.87 ± 0.07 | 26.45 ± 0.20 | 26.43 ± 0.20 |
| 6 | 26.62 ± 0.15 | 26.69 ± 0.14 | 26.50 ± 0.11 | 26.49 ± 0.11 |
| 7 | 26.68 ± 0.14 | 26.75 ± 0.21 | 26.52 ± 0.02 | 26.40 ± 0.02 |
| 8 | 26.71 ± 0.09 | 26.76 ± 0.03 | 26.47 ± 0.04 | 26.63 ± 0.04 |
| 9 | 26.66 ± 0.09 | 26.72 ± 0.06 | 26.63 ± 0.10 | 26.43 ± 0.10 |
| 10 | 26.74 ± 0.11 | 26.77 ± 0.18 | 26.68 ± 0.03 | 26.42 ± 0.03 |
| Sampling Period | P. spumarius | P. italosignus | ||||
|---|---|---|---|---|---|---|
| Females | Males | Total | Females | Males | Total | |
| December 2021 | 10 | 5 | 15 | 2 | 0 | 2 |
| June 2022 | 0 | 1 | 1 | 14 | 11 | 25 |
| Order | Family | Species | Origin of Samples | Number of Specimens Assayed | Life Stage |
|---|---|---|---|---|---|
| Hemiptera | Pentatomidae | Rhaphigaster nebulosa (Poda, 1761) | RPS—Florence | 1 | Adult |
| Tingidae | Stephanitis lauri Rietschel, 2014 | University of Pisa | 1 | Adult | |
| Ricaniidae | Ricania speculum (Walker, 1851) | University of Pisa | 1 | Adult | |
| Cicadellidae | Cicadella viridis (Linnaeus, 1758) | RPS—Florence | 1 | Adult | |
| Synophropsis lauri (Horvath, 1897) | University of Florence | 1 | Adult | ||
| Membracidae | Stictocephala bisonia Kopp &Yonke, 1977 | University of Florence | 1 | Adult | |
| Aphrophoridae | Philaenus spumarius (Linnaeus, 1758) | University of Florence | 37 | Adult | |
| Philaenus italosignus Drosopoulos & Remane, 2000 | University of Florence | 12 | Adult | ||
| Neophilaenus campestris (Fallen, 1805) | University of Florence | 75 | Adult | ||
| Cercopidae | Lepyronia coleoptrata (Linnaeus, 1758) | University of Florence | 3 | Adult | |
| Cercopis sanguinolenta (Scopoli, 1763) | University of Florence | 3 | Adult | ||
| Cercopis vulnerata Rossi, 1807 | University of Florence | 3 | Adult | ||
| Dictyopharidae | Dictyophara europaea (Linnaeus, 1767) | RPS—Florence | 1 | Adult | |
| Aleyrodidae | Dialeurodes citri (Ashmead, 1885) | RPS—Florence | 1 | Juvenile | |
| Lepidoptera | Crambidae | Cydalima perspectalis (Walker, 1859) | RPS—Florence | 1 | Larva |
| Tortricidae | Grapholita (Aspila) molesta (Busck, 1916) | University of Florence | 1 | Adult | |
| Cydia pomonella (Linnaeus, 1758) | University of Florence | 1 | Adult | ||
| Cryptoblabes gnidiella (Millière, 1867) | University of Florence | 1 | Adult | ||
| Diptera | Tephritidae | Ceratitis capitata (Wiedemann 1824) | University of Florence | 2 | Adult |
| Ceratitis capitata (Wiedemann 1824) | University of Florence | 2 | Larva | ||
| Rhagoletis cerasi (Linnaeus, 1758) | University of Florence | 1 | Pupa | ||
| Rhagoletis completa Cresson, 1929 | RPS—Florence | 1 | Larva | ||
| Acanthiophilus helianthi (Rossi, 1794) | University of Pisa | 1 | Adult |
| Primer Name | Length (Nucleotides) | Sequence 5′–3′ | Product Size (bp) |
|---|---|---|---|
| Pital_348F | 28 | GCCTATGTAATCAATATATTTATTCAAA | 119 |
| Pital_465R | 28 | AATGGAGAAATTAATAATGAAATAGTAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizzo, D.; Bracalini, M.; Campigli, S.; Nencioni, A.; Porcelli, F.; Marchi, G.; Da Lio, D.; Bartolini, L.; Rossi, E.; Sacchetti, P.; et al. Quantitative Real-Time PCR Based on SYBR Green Technology for the Identification of Philaenus italosignus Drosopoulos & Remane (Hemiptera Aphrophoridae). Plants 2022, 11, 3314. https://doi.org/10.3390/plants11233314
Rizzo D, Bracalini M, Campigli S, Nencioni A, Porcelli F, Marchi G, Da Lio D, Bartolini L, Rossi E, Sacchetti P, et al. Quantitative Real-Time PCR Based on SYBR Green Technology for the Identification of Philaenus italosignus Drosopoulos & Remane (Hemiptera Aphrophoridae). Plants. 2022; 11(23):3314. https://doi.org/10.3390/plants11233314
Chicago/Turabian StyleRizzo, Domenico, Matteo Bracalini, Sara Campigli, Anita Nencioni, Francesco Porcelli, Guido Marchi, Daniele Da Lio, Linda Bartolini, Elisabetta Rossi, Patrizia Sacchetti, and et al. 2022. "Quantitative Real-Time PCR Based on SYBR Green Technology for the Identification of Philaenus italosignus Drosopoulos & Remane (Hemiptera Aphrophoridae)" Plants 11, no. 23: 3314. https://doi.org/10.3390/plants11233314