
Proteomic Analysis of Barley (Hordeum vulgare L.) Leaves in Response to Date Palm Waste Compost Application
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Growth Traits and Physiological Parameters
2.2. Identification of Differentially Abundant Proteins (DAPs) in Response to Date Palm Waste Compost Application
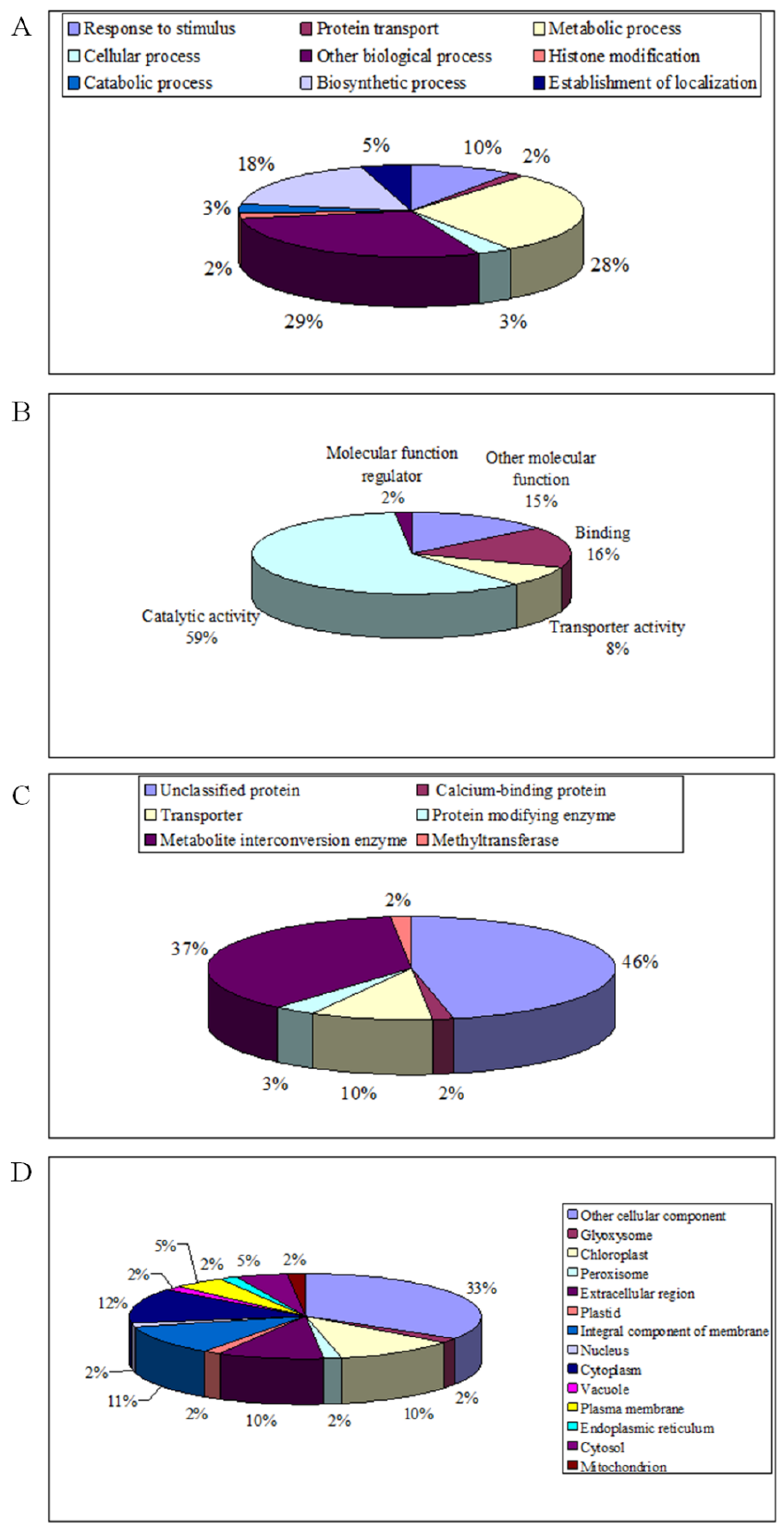
2.3. Biological Process, Molecular Function, Protein Class and Subcellular Localization of DAPs
2.4. KEGG Pathway Enrichment Analyses of DAPs
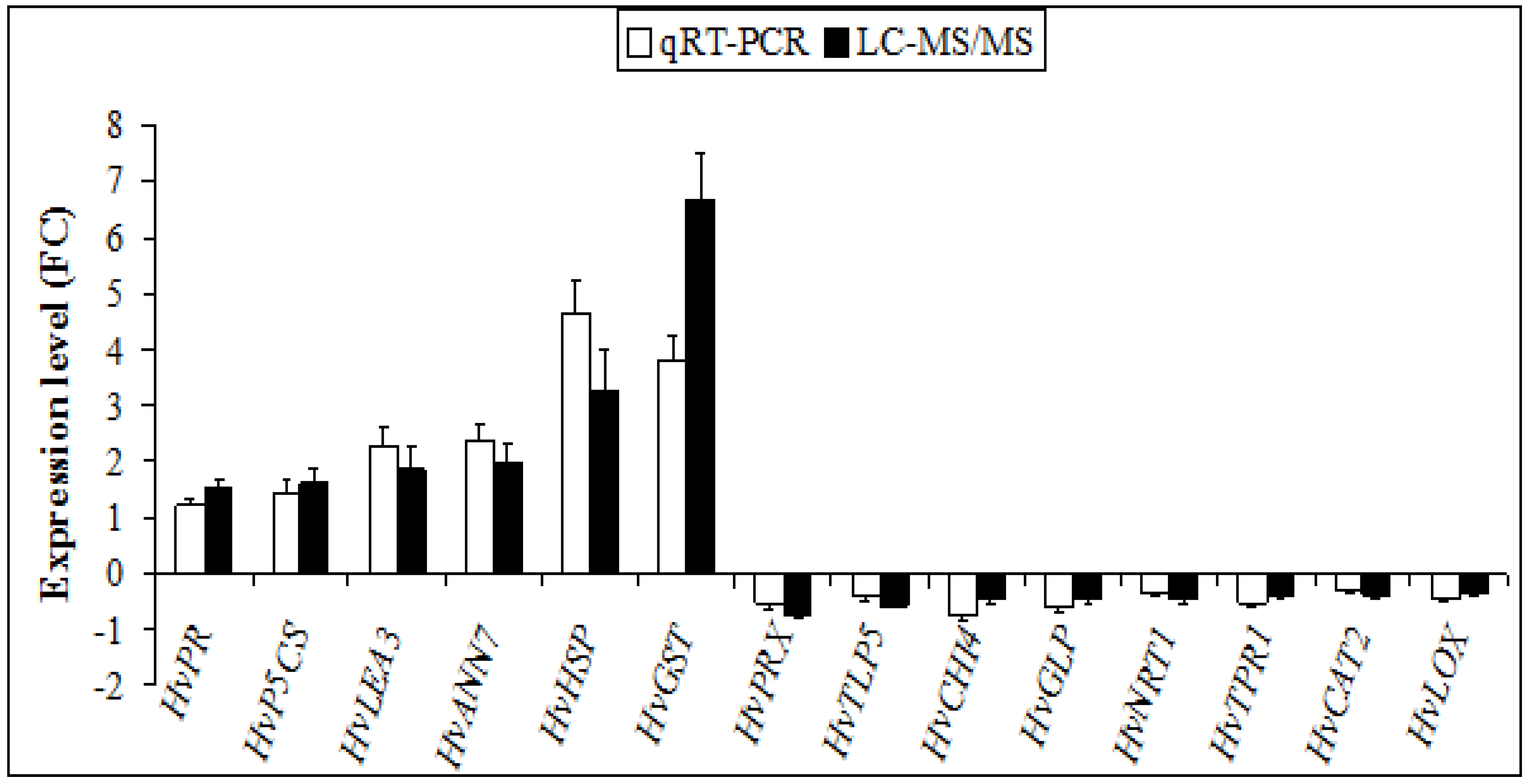
2.5. Protein Profile Validation by Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
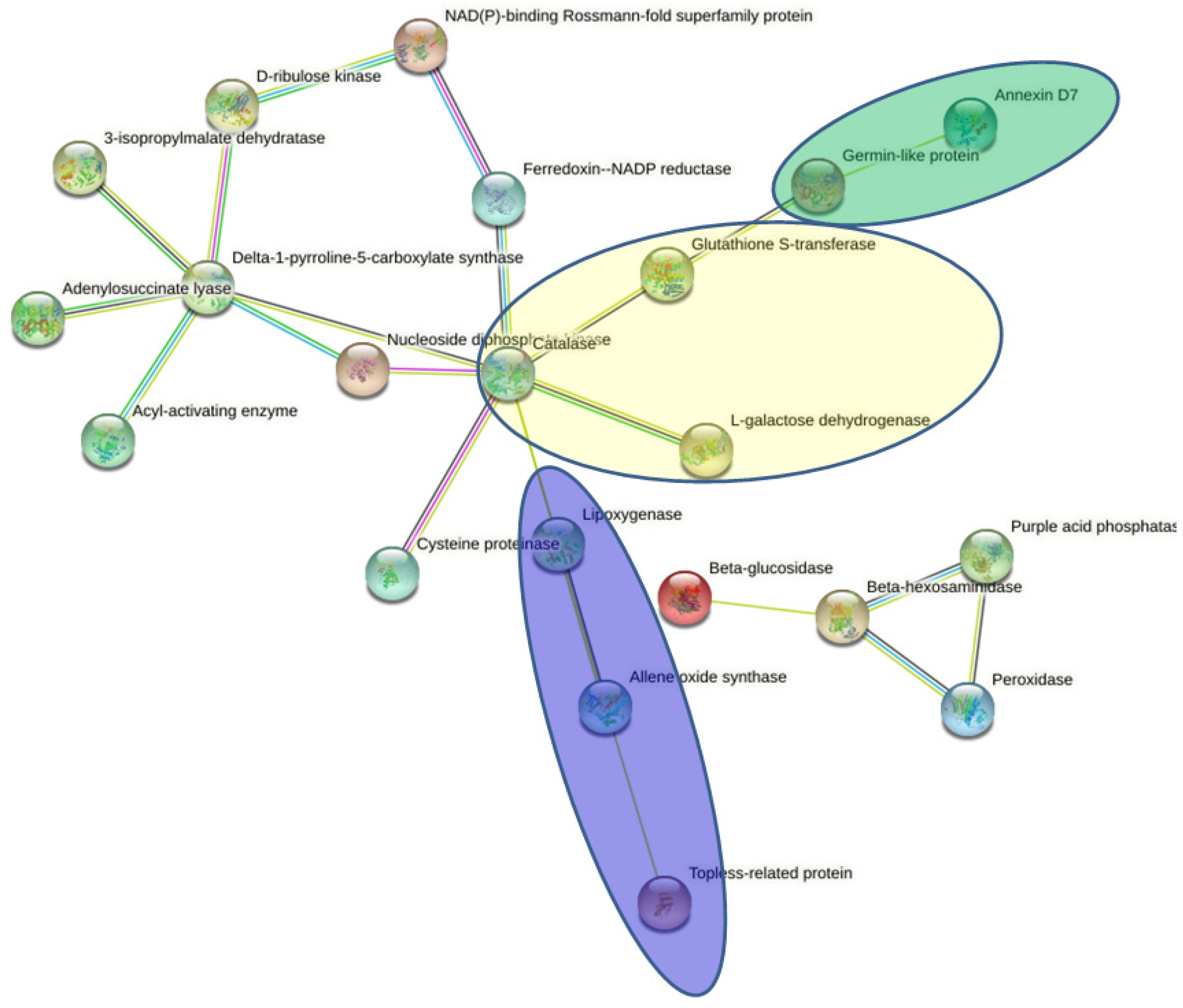
2.6. Protein–Protein Interaction (PPI) Network Analysis of DAPs
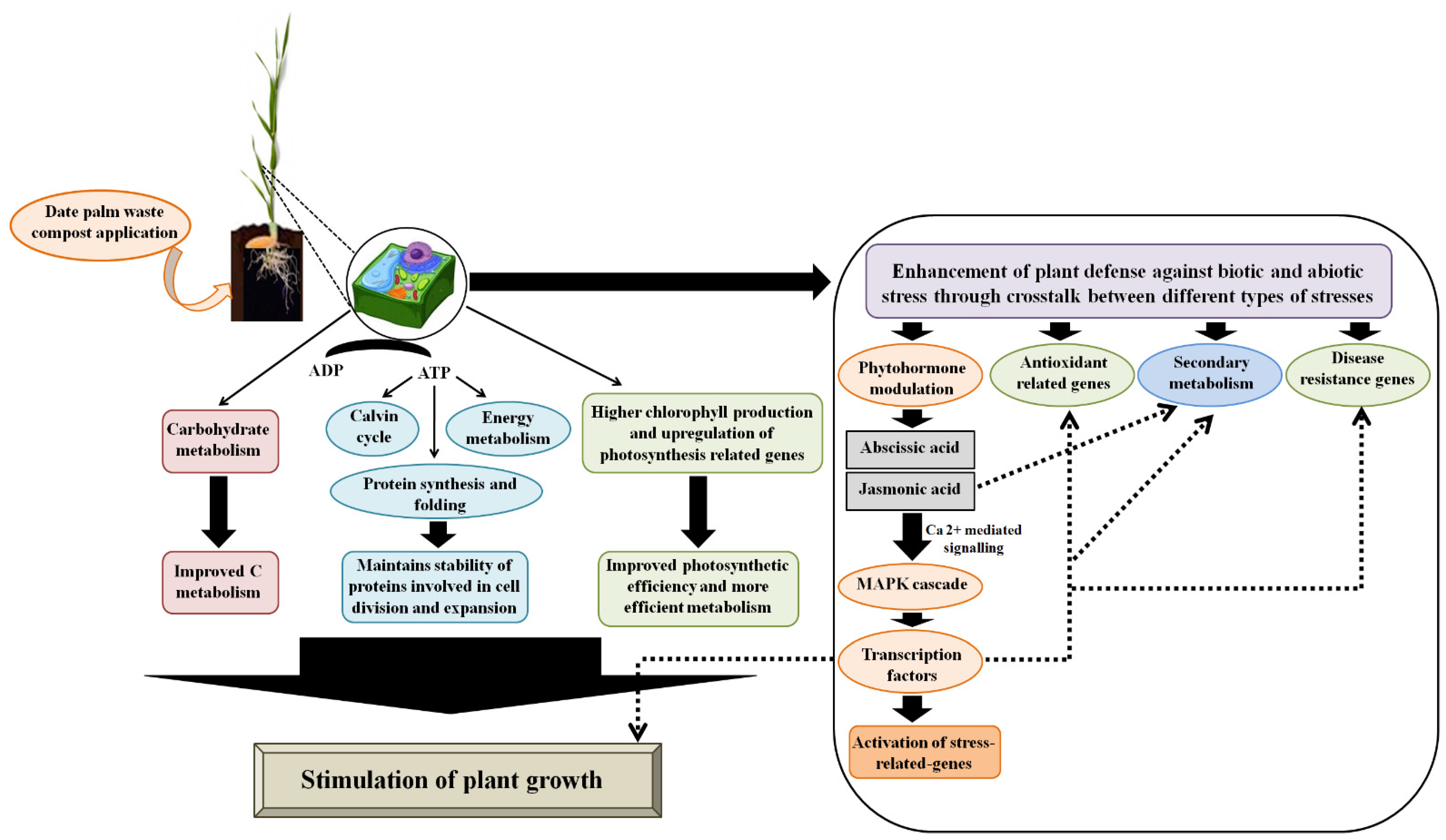
2.7. Proposed Model of a Possible Mode of Action of Compost in Barley Plants at the Tillering Stage
3. Discussion
3.1. Application of Date Palm Waste Compost Promote Growth of Barley Plants
3.2. Application of Date Palm Waste Compost Altered Barley Leaf Protein Profiles
3.3. Proteins Involved in Redox Homeostasis
3.4. Proteins Involved in Stress Response
3.5. Proteins Involved in Energy Metabolism, Photosynthesis and Chlorophyll Metabolism
3.6. Proteins Involved in Amino Acid Metabolism
3.7. Proteins Involved in Protein Metabolism
3.8. Proteins Involved in Biosynthesis of Secondary Metabolites
3.9. Proteins Involved in Hormone Metabolism
3.10. Proteins Involved in Transport
3.11. Proteins Related to Epigenetic Regulation Mechanisms
3.12. Proteins Related to Other Functional Categories
4. Materials and Methods
4.1. Experimental Design and Operations of the Experiment
4.2. Agronomic and Physiological Parameters
4.3. Total Protein Extraction and Digestion
4.4. Protein LC-MS/MS Analyses
4.5. Protein Identification and Quantification
4.6. Bioinformatics and Data Analysis
4.7. Total RNA Extraction, cDNA Synthesis and Quantitative Real-Time PCR (qRT-PCR) Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vukovic, A.; Velki, M.; Ecimovic, S.; Vukovic, R.; Štolfa Camagajevac, I.; Loncaric, Z. Vermicomposting-facts, benefits and knowledge gaps. Agronomy 2021, 11, 1952. [Google Scholar] [CrossRef]
- Ho, T.T.K.; Tra, V.T.; Le, T.H.; Nguyen, N.K.Q.; Tran, C.S.; Nguyen, P.T.; Vo, T.D.H.; Thai, V.N.; Bui, X.T. Compost to improve sustainable soil cultivation and crop productivity. Case Stud. Chem. Environ. Eng. 2022, 6, 100211. [Google Scholar] [CrossRef]
- Yatoo, A.M.; Ali, M.N.; Baba, Z.A.; Hassan, B. Sustainable management of diseases and pests in crops by vermicompost and vermicompost tea. A review. Agron. Sustain. Dev. 2021, 41, 7. [Google Scholar]
- Omara, A.E.D.; Hafez, E.M.; Osman, H.S.; Rashwan, E.; El-Said, M.A.A.; Alharbi, K.; Abd El-Moneim, D.; Gowayed, S.M. Collaborative impact of compost and beneficial rhizobacteria on soil properties, physiological attributes, and productivity of wheat subjected to deficit irrigation in salt affected soil. Plants 2022, 11, 877. [Google Scholar] [CrossRef] [PubMed]
- Sayara, T.; Basheer-Salimia, R.; Hawamde, F.; Sánchez, A. Recycling of organic wastes through composting: Process performance and compost application in agriculture. Agronomy 2020, 10, 1838. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Miras Moreno, M.B.; Reynaud, H.; Canaguier, R.; Trtílek, M.; et al. Combined phenotypic and metabolomic approach for elucidating the biostimulant action of a plant-derived protein hydrolysate on tomato grown under limited water availability. Front. Plant Sci. 2019, 10, 493. [Google Scholar] [CrossRef]
- González-Morales, S.; Solís-Gaona, S.; Valdés-Caballero, M.V.; Juárez-Maldonado, A.; Loredo-Treviño, A.; Benavides-Mendoza, A. Transcriptomics of biostimulation of plants under abiotic stress. Front. Genet. 2021, 12, 583888. [Google Scholar] [CrossRef]
- Roomi, S.; Masi, A.; Conselvan, G.B.; Trevisan, S.; Quaggiotti, S.; Pivato, M.; Arrigoni, G.; Yasmin, T.; Carletti, P. Protein profiling of Arabidopsis roots treated with humic substances: Insights into the metabolic and interactome networks. Front. Plant Sci. 2018, 9, 1812. [Google Scholar] [CrossRef] [Green Version]
- Ghouili, E.; Hidri, Y.; Cheikh M’Hamed, H.; Somenahally, A.; Xue, Q.; El Akram Znaïdi, I.; Jebara, M.; Nefissi Ouertani, R.; Muhovski, Y.; Riahi, J.; et al. Date palm waste compost promotes plant growth and nutrient transporter genes expression in barley (Hordeum vulgare L.). S. Afr. J. Bot. 2022, 149, 247–257. [Google Scholar] [CrossRef]
- Ren, W.; Shi, Z.; Zhou, M.; Zhao, B.; Li, H.; Wang, J.; Liu, Y.; Zhao, J. iTRAQ-based quantitative proteomic analysis provides insight into the drought-stress response in maize seedlings. Sci. Rep. 2022, 12, 9520. [Google Scholar] [CrossRef]
- Benabderrahim, M.A.; Elfalleh, W.; Belayadi, H.; Haddad, M. Effect of date palm waste compost on forage alfalfa growth, yield, seed yield and minerals uptake. Int. J. Recycl. Org. Waste Agric. 2018, 7, 1–9. [Google Scholar] [CrossRef]
- Abid, W.; Magdich, S.; Ben Mahmoud, I.; Medhioub, K.; Ammar, E. Date palm wastes co-composted product: An efficient substrate for tomato (Solanum lycopercicum L.) seedling production. Waste Biomass Valor. 2016, 9, 45–55. [Google Scholar] [CrossRef]
- Głąba, T.; Żabiński, A.; Sadowska, U.; Gondek, K.; Kopećb, M.; Mierzwa-Hersztek, M.; Tabor, S.; Stanek-Tarkowska, J. Fertilization effects of compost produced from maize, sewage sludge and biochar on soil water retention and chemical properties. Soil Tillage Res. 2020, 197, 104493. [Google Scholar] [CrossRef]
- Rekaby, S.A.; Awad, M.Y.M.; Hegab, S.A.; Eissa, M.A. Effect of some organic amendments on barley plants under saline condition. J. Plant Nutr. 2020, 43, 1840–1851. [Google Scholar] [CrossRef]
- Bashir, A.; Rizwan, M.; Zia ur Rehman, M.; Zubair, M.; Riaz, M.; Qayyum, M.F.; Alharby, H.F.; Bamagoos, A.A.; Ali, S. Application of co-composted farm manure and biochar increased the wheat growth and decreased cadmium accumulation in plants under different water regimes. Chemosphere 2020, 246, 125809. [Google Scholar] [CrossRef]
- Li, J.; Ali, E.F.; Majrashi, A.; Eissa, M.A.; Ibrahim, O.H.M. Compost enhances forage yield and quality of river saltbush in arid conditions. Agriculture 2021, 11, 595. [Google Scholar] [CrossRef]
- Ebinezer, L.B.; Franchin, C.; Trentin, A.R.; Carletti, P.; Trevisan, S.; Agrawal, G.K.; Rakwal, R.; Quaggiotti, S.; Arrigoni, G.; Masi, A. Quantitative proteomics of maize roots treated with a protein hydrolysate: A comparative study with transcriptomics highlights the molecular mechanisms responsive to biostimulants. J. Agric. Food Chem. 2020, 28, 7541–7553. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-wide identification of differentially expressed genes in Solanum lycopersicon L. in response to an alfalfa-protein hydrolysate using microarrays. Front. Plant Sci. 2017, 8, 1159. [Google Scholar] [CrossRef] [Green Version]
- Trevisan, S.; Botton, A.; Vaccaro, S.; Vezzaro, A.; Quaggiotti, S.; Nardi, S. Humic substances affect Arabidopsis physiology by altering the expression of genes involved in primary metabolism, growth and development. Environ. Exp. Bot. 2011, 74, 45–55. [Google Scholar] [CrossRef]
- Goñi, O.; Fort, A.; Quille, P.; McKeown, P.C.; Spillane, C.; O’Connell, S. Comparative transcriptome analysis of two ascophyllum nodosum extract biostimulants: Same seaweed but different. J. Agric. Food Chem. 2016, 64, 2980–2989. [Google Scholar] [CrossRef]
- Nunes, R.O.; Domiciano, G.A.; Alves, W.S.; Melo, A.C.A.; Nogueira, F.C.S.; Canellas, L.P.; Olivares, F.L.; Zingali, R.B.; Soares, M.R. Evaluation of the effects of humic acids on maize root architecture by label-free proteomics analysis. Sci. Rep. 2019, 9, 12019. [Google Scholar] [CrossRef] [PubMed]
- Martinez Esteso, M.J.; Vilella-Anton, M.; Selles-Marchart, S.; Martinez-Marquez, A.; Botta-Catala, A.; Pinol-Dastis, R.; BruMartinez, R. A DIGE proteomic analysis of wheat flag leaf treated with TERRA-SORB foliar, a free amino acid high content biostimulant. J. Integr. OMICS 2016, 6, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS Regulation of Plant Development and Stress Responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- García, A.C.; van Tol de Castro, T.A.; Santos, L.A.; Tavares, O.C.H.; Castro, R.N.; Berbara, R.L.L.; García-Mina, J.M. Structure-property-function relationship of humic substances in modulating the root growth of plants: A review. J. Environ. Qual. 2019, 48, 1622–1632. [Google Scholar] [CrossRef]
- Naboulsi, I.; Ben Mrid, R.; Ennoury, A.; Zouaoui, Z.; Nhiri, M.; Ben Bakrim, W.; Yasri, A.; Aboulmouhajir, A. Crataegus oxyacantha extract as a biostimulant to enhance tolerance to salinity in tomato plants. Plants 2022, 11, 1283. [Google Scholar] [CrossRef]
- Baltazar, M.; Correia, S.; Guinan, K.J.; Sujeeth, N.; Bragança, R.; Gonçalves, B. Recent advances in the molecular effects of biostimulants in plants: An overview. Biomolecules 2021, 11, 1096. [Google Scholar] [CrossRef]
- Franzoni, G.; Cocetta, G.; Prinsi, B.; Ferrante, A.; Espen, L. Biostimulants on crops: Their impact under abiotic stress conditions. Horticulturae 2022, 8, 189. [Google Scholar] [CrossRef]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for plant growth and mitigation of abiotic stresses: A metabolomics perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef]
- Pichereauxa, C.; Laurentc, E.A.; Gargarosb, A.; Viudesc, A.; Durieue, C.; Lamazed, T.; Grieuc, P.; Burlet-Schiltz, O. Analysis of durum wheat proteome changes under marine and fungal biostimulant treatments using large-scale quantitative proteomics: A useful dataset of durum wheat proteins. J. Proteom. 2019, 200, 28–39. [Google Scholar] [CrossRef]
- Jacob, P.; Hirt, H.; Bendahmane, A. The heat-shock protein/chaperone network and multiple stress resistance. Plant Biotechnol. J. 2017, 15, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Hong-Bo, S.; Zong-Suo, L.; Ming-An, S. LEA proteins in higher plants: Structure, function, gene expression and regulation. Colloids Surf. B Biointerfaces 2005, 45, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Jami, S.K.; Clark, G.B.; Ayele, B.T.; Ashe, P.; Kirti, P.B. Genomewide comparative analysis of annexin superfamily in plants. PLoS ONE 2012, 7, e47801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szalonek, M.; Sierpien, B.; Rymaszewski, W.; Gieczewska, K.; Garstka, M.; Lichocka, M.; Sass, L.; Paul, K.; Vass, I.; Vankova, R.; et al. Potato annexin STANN1 promotes drought tolerance and mitigates light Ssress in transgenic Solanum tuberosum L. plants. PLoS ONE 2015, 10, e0132683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jami, S.K.; Clark, G.B.; Turlapati, S.A.; Handley, C.; Roux, S.J.; Kirti, P.B. Ectopic expression of an annexin from Brassica juncea confers tolerance to abiotic and biotic stress treatments in transgenic tobacco. Plant Physiol. Biochem. 2008, 46, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.Y.; Kang, S.H.; Ali, I.; Lee, S.C.; Ji, M.G.; Jeong, S.Y.; Shin, G.I.; Kim, M.G.; Jeon, J.R.; Kim, W.Y. Humic acid enhances heat stress tolerance via transcriptional activation of Heat-Shock Proteins in Arabidopsis. Sci. Rep. 2020, 10, 15042. [Google Scholar] [CrossRef]
- Pereira, R.V.; Filgueiras, C.C.; Dória, J.; Peñaflor, M.F.G.V.; Willett, D.S. The effects of biostimulants on induced plant defense. Front. Agron. 2021, 3, 630596. [Google Scholar] [CrossRef]
- Zribi, I.; Ghorbel, M.; Brini, F. Pathogenesis related proteins (PRs): From cellular mechanisms to plant defense. Curr. Protein Pept. Sci. 2021, 22, 396–412. [Google Scholar] [CrossRef]
- Vasyukova, N.I.; Zinoveva, S.V.; Il’inskaya, L.I.; Perekhod, E.A.; Chalenko, G.I.; Gerasimova, N.G.; Il’ina, A.V.; Varlamov, V.P.; Ozeretskovskaya, O.L. Modulation of plant resistance to diseases by water-soluble chitosan. Appl. Biochem. Microbiol. 2001, 37, 103–109. [Google Scholar] [CrossRef]
- Jayaraj, J.; Wan, A.; Rahman, M.; Punja, Z.K. Seaweed extract reduces foliar fungal diseases on carrot. Crop Prot. 2008, 27, 1360–1366. [Google Scholar] [CrossRef]
- Gunupuru, L.R.; Patel, J.S.; Sumarah, M.W.; Renaud, J.B.; Mantin, E.G.; Prithiviraj, B. A plant biostimulant made from the marine brown algae Ascophyllum nodosum and chitosan reduce Fusarium head blight and mycotoxin contamination in wheat. PLoS ONE 2019, 14, e0220562. [Google Scholar] [CrossRef] [Green Version]
- Oikawa, A.; Yamashita, T.; Taira, H.; Ejiri, S.; Kidou, S. Jasmonate-induced 23kD protein, JIP-23, is involved in seed development of barley. Plant Biotechnol. 2007, 24, 217–220. [Google Scholar] [CrossRef]
- Lotan, T.; Ori, N.; Fluhr, R. Pathogenesis-related proteins are developmentally regulated in tobacco flowers. Plant Cell. 1989, 1, 881–887. [Google Scholar] [PubMed] [Green Version]
- Banerjee, J.; Maiti, M.K. Functional role of rice germin-like protein1 in regulation of plant height and disease resistance. Biochem. Biophys. Res. Commun. 2010, 394, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Y.; Xiao, N.; Zhang, G.; Wang, F.; Chen, X.; Fang, R. Rice GERMIN-LIKE PROTEIN 2-1 functions in seed dormancy under the control of abscisic acid and gibberellic acid signaling pathways. Plant Physiol. 2020, 183, 1157–1170. [Google Scholar] [CrossRef] [PubMed]
- Perveen, S.; Khan, T.A.; Shaheen, H.; Naz, R.; Hyder, M.Z.; Ijaz, B.; Naqvi, S.M.S.; Yasmin, T. Morpho-physiological investigations in transgenic tomato (Solanum lycopersicum L.) over expressing OsRGLP1 gene. Vitr. Cell Dev. Biol.-Plant 2021, 57, 907–922. [Google Scholar] [CrossRef]
- Liao, L.; Hu, Z.; Liu, S.; Yang, Y.; Zhou, Y. Characterization of germin-like proteins (GLPs) and their expression in response to abiotic and biotic stresses in cucumber. Horticulturae 2021, 7, 412. [Google Scholar] [CrossRef]
- Banerjee, J.; Gantait, S.; Maiti, M.K. Physiological role of rice germin-like protein 1 (OsGLP1) at early stages of growth and development in indica rice cultivar under salt stress condition. Plant Cell Tissue Organ Cult. 2017, 131, 127–137. [Google Scholar] [CrossRef]
- Ertani, A.; Sambo, P.; Nicoletto, C.; Santagata, S.; Schiavon, M.; Nardi, S. The use of organic biostimulants in hot pepper plants to help low input sustainable agriculture. Chem. Biol. Technol. Agric. 2015, 2, 11. [Google Scholar] [CrossRef] [Green Version]
- Kavi Kishor, P.B.; Hima Kumari, P.; Sunita, M.S.L.; Sreenivasulu, N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, F.; Kopka, J.; Haskell, D.W.; Zhao, W.; Schiller, K.C.; Gatzke, N.; Sung, D.Y.; Guy, C.L. Exploring the temperature-stress metabolome of Arabidopsis. Plant Physiol. 2004, 136, 4159–4168. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Wang, Z.; Yu, W.; Liu, Y.; Huang, B. Differential metabolic responses of perennial grass Cynodon transvaalensis × Cynodon dactylon (C4) and Poa pratensis (C3) to heat stress. Physiol. Plant. 2011, 141, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Arruda, P.; Barreto, P. Lysine Catabolism Through the saccharopine pathway: Enzymes and intermediates involved in plant responses to abiotic and biotic stress. Front. Plant Sci. 2020, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Alfosea-Simón, M.; Zavala-Gonzalez, E.A.; Camara-Zapata, J.M.; Martínez-Nicolás, J.J.; Simón, I.; Simón-Grao, S.; García-Sánchez, F. Effect of foliar application of amino acids on the salinity tolerance of tomato plants cultivated under hydroponic system. Sci. Hortic. 2020, 272, 109509. [Google Scholar] [CrossRef]
- Novaes, E.; Kirst, M.; Chiang, V.; Winter-Sederoff, H.; Sederoff, R. Lignin and biomass: A negative correlation for wood formation and lignin content in trees. Plant Physiol. 2010, 154, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Mertz, I.T.; Christians, N.E.; Thoms, A.W. Utilizing branched-chain amino acids for increasing shoot density and establishment rate in creeping bentgrass. HortTechnology 2020, 30, 102–106. [Google Scholar] [CrossRef] [Green Version]
- Casadesus, A.; Perez-Llorca, M.; Munne-Bosch, S.; Polo, J. An enzymatically hydrolyzed animal protein-based biostimulant (pepton) increases salicylic acid and promotes growth of tomato roots under temperature and nutrient stress. Front. Plant Sci. 2020, 11, 953. [Google Scholar] [CrossRef]
- Pratelli, R.; Pilot, G. Regulation of amino acid metabolic enzymes and transporters in plants. J. Exp. Bot. 2014, 65, 5535–5556. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nesi, A.N.; Araujo, W.L.; Braun, H.P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.; Zang, X.; Cong, X.; Hou, L.; He, B.; Ding, Y.; Dong, M.; Sun, D.; Guo, Y.; Zhang, F.; et al. Cloning of nitrite reductase gene from Haematococcus pluvialis and transcription and enzymatic activity analysis at different nitrate and phosphorus concentration. Gene 2019, 697, 123–130. [Google Scholar] [CrossRef]
- Wang, W.; Xu, M.; Wang, G.; Galili, G. New insights into the metabolism of aspartate-family amino acids in plant seeds. Plant Rep. 2018, 31, 203–211. [Google Scholar] [CrossRef]
- Zhang, J.; Yuan, H.; Yang, Y.; Fish, T.; Lyi, S.M.; Thannhauser, T.W.; Zhang, L.; Li, L. Plastid ribosomal protein S5 is involved in photosynthesis, plant development, and cold stress tolerance in Arabidopsis. J. Exp. Bot. 2016, 67, 2731–2744. [Google Scholar] [CrossRef]
- Luengo, T.M.; Mayer, M.P.; Rüdige, S.G.D. The Hsp70–Hsp90 chaperone cascade in protein folding. Trends Cell Biol. 2019, 29, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Larkin, R.M. Tetrapyrrole signaling in plants. Front. Plant Sci. 2015, 7, 1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, M.; Li, Z.; Zhu, L.; Wang, J.; Zhang, B.; Zheng, F.; Zhao, B.; Zhang, H.; Wang, Y.; Zhang, Z. The multiple roles of ascorbate in the abiotic stress response of plants: Antioxidant, cofactor, and regulator. Front. Plant Sci. 2021, 12, 598173. [Google Scholar] [CrossRef] [PubMed]
- Zdyb, A.; Salgado, M.G.; Demchenko, K.N.; Brenner, W.G.; Płaszczyca, M.; Stumpe, M.; Herrfurth, C.; Feussner, I.; Pawlowski, K. Allene oxide synthase, allene oxide cyclase and jasmonic acid levels in Lotus japonicus nodules. PLoS ONE 2018, 13, e0190884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, S.Y.; Zhang, X.F.; Lu, Z.; Xin, Q.; Wu, Z.; Jiang, T.; Lu, Y.; Wang, X.F.; Zhang, D.F. Roles of the different components of magnesium chelatase in abscisic acid signal transduction. Plant Mol. Biol. 2012, 80, 519–537. [Google Scholar] [CrossRef] [Green Version]
- Vishwakarma, K.; Mishra, M.; Patil, G.; Mulkey, S.; Ramawat, N.; Singh, V.P.; Deshmukh, R.; Tripathi, D.K.; Nguyen, H.T.; Sharma, S. Avenues of the membrane transport system in adaptation of plants to abiotic stresses. Crit. Rev. Biotechnol. 2019, 39, 861–883. [Google Scholar] [CrossRef]
- Lephatsi, M.; Nephali, L.; Meyer, V.; Piater, L.A.; Buthelezi, N.; Dubery, I.A.; Opperman, H.; Brand, M.; Huyser, J.; Tugizimana, F. Molecular mechanisms associated with microbial biostimulant-mediated growth enhancement, priming and drought stress tolerance in maize plants. Sci. Rep. 2022, 12, 10450. [Google Scholar] [CrossRef]
- De Palma, M.; Salzano, M.; Villano, C.; Aversano, R.; Lorito, M.; Ruocco, M.; Docimo, T.; Piccinelli, A.L.; D’Agostino, N.; Tucc, M. Transcriptome reprogramming, epigenetic modifications and alternative splicing orchestrate the tomato root response to the beneficial fungus Trichoderma harzianum. Hortic. Res. 2019, 6, 5. [Google Scholar] [CrossRef]
- Yang, D.; Huang, Z.; Jin, W.; Xia, P.; Jia, Q.; Yang, Z.; Hou, Z.; Zhang, H.; Ji, W.; Han, R. DNA methylation: A new regulator of phenolic acids biosynthesis in Salvia miltiorrhiza. Ind. Crops Prod. 2018, 124, 402–411. [Google Scholar] [CrossRef]
- Jannin, L.; Arkoun, M.; Ourry, A.; Laîné, P.; Goux, D.; Garnica, M.; Fuentes, M.; Francisco, S.S.; Baigorri, R.; Cruz, F.; et al. Microarray analysis of humic acid effects on Brassica napus growth: Involvement of N, C and S metabolisms. Plant Soil 2012, 359, 297–319. [Google Scholar] [CrossRef]
- Pandian, B.A.; Sathishraj, R.; Djanaguiraman, M.; Prasad, P.V.V.; Jugulam, M. Role of cytochrome P450 enzymes in plant stress response. Antioxidants 2020, 9, 454. [Google Scholar] [CrossRef]
- Zhao, J.; Qiu, Z.; Ruan, B.; Kang, S.; He, L.; Zhang, S.; Dong, G.; Hu, J.; Zeng, D.; Zhang, G.; et al. Functional inactivation of putative photosynthetic electron acceptor ferredoxin C2 (FdC2) induces delayed heading date and decreased photosynthetic rate in rice. PLoS ONE 2015, 10, e0143361. [Google Scholar] [CrossRef]
- Alam, J.; Whitaker, R.A.; Krogmann, D.W.; Curtis, S.E. Isolation and sequence of the gene for ferredoxin i from the cyanobacterium anabaena sp. strain pcc 7120. J. Bacteriol. 1986, 168, 1265–1271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Sun, Q.; Wu, J.; Zhao, P.; Sun, Y.; Guo, Z. Genome-wide identification and characterization of short-chain dehydrogenase/reductase (SDR) gene family in medicago truncatula. Int. J. Mol. Sci. 2021, 22, 9498. [Google Scholar] [CrossRef] [PubMed]
- Arnon, D.I. Copper enzymes in isolated chloroplasts, polyphenol oxidase in Beta vulgaris. Plant. Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Faurobert, M.; Pelpoir, E.; Chaïb, J. Phenol extraction of proteins for proteomic studies of recalcitrant plant tissues. Methods Mol. Biol. 2007, 355, 9–14. [Google Scholar]
- Belouah, I.; Benard, C.; Denton, A.; Blein-Nicolas, M.; Balliau, T.; Teyssier, E.; Gallusci, P.; Bouchez, O.; Usadel, B.; Zivy, M.; et al. Transcriptomic and proteomic data in developing tomato fruit. Data Brief 2020, 28, 105015. [Google Scholar] [CrossRef]
- Kessner, D.; Chambers, M.; Burke, R.; Agus, D.; Mallick, P. ProteoWizard: Open source software for rapid proteomics tools development. Bioinformatics 2008, 24, 2534–2536. [Google Scholar] [CrossRef] [Green Version]
- Valot, B.; Langella, O.; Nano, E.; Zivy, M. MassChroQ: A versatile tool for mass spectrometry quantification. Proteomics 2011, 11, 3572–3577. [Google Scholar] [CrossRef] [Green Version]
- Balliau, T.; Duruflé, H.; Blanchet, N.; Blein-Nicolas, M.; Langlade, N.B.; Zivy, M. Proteomic data from leaves of twenty-four sunflower genotypes under water deficit. OCL 2021, 28, 12. [Google Scholar] [CrossRef]
- Wang, H.; Wu, K.K.; Liu, Y.; Wu, Y.F.; Wang, X.F. Integrative proteomics to understand the transmission mechanism of Barley yellow dwarf virus-GPV by its insect vector Rhopalosiphum padi. Sci. Rep. 2015, 5, 10971. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; von Mering, C.; et al. STRINGv9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, 808–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Rozen, S.; Skaletsky, H.J. Primer3 on the WWW for general users and for biologist programmers. In Bioinformatics Methods and Protocols: Methods in Molecular Biology; Krawetz, S., Misener, S., Eds.; Humana Press: Totowa, NJ, USA, 2000; pp. 365–386. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Control | Compost |
|---|---|---|
| SL (cm) | 39.65 ± 0.49 b | 51.20 ± 1.8 3 a |
| SDW (mg plant−1) | 329.10 ± 7.07 b | 536.06 ± 21.22 a |
| Chla (mg g−1 FW) | 10.47 ± 0.41 b | 12.67 ± 0.37 a |
| Chlb (mg g−1 FW) | 2.84 ± 0.13 b | 3.28 ± 0.21 a |
| Chlt (mg g−1 FW) | 13.32 ± 0.55 b | 15.67 ± 0.15 a |
| Cart (mg g−1 FW) | 0.38 ± 0.02 b | 0.83 ± 0.08 a |
| Ci (µmol CO2 mol–1) | 282.00 ± 2.82 b | 298.00 ± 4.24 a |
| E (mmol H2O m–2 S–1) | 0.49 ± 0.02 b | 0.82 ± 0.01 a |
| gs (µmol H2O m–2 S–1) | 49.50 ± 3.53 b | 87.50 ± 4.94 a |
| Pn (µmol CO2 m–2 S–1) | 4.87 ± 0.50 b | 9.27 ± 0.62 a |
| No | Protein Name | Function | Regulated Type | Fold Change | Protein Accession | KEGG Pathway |
|---|---|---|---|---|---|---|
| 1 | Pathogenesis-related protein 1 | Defense response | Up | 1.52 | P16273 | MAPK signaling pathway |
| 2 | Ribosome-binding factor PSRP1 | Protein biosynthesis | Up | 1.52 | F2DV60 | Other pathways |
| 3 | Nucleoside diphosphate kinase 1 | Nucleotide metabolism | Up | 1.52 | A0A287NWM2 | Metabolic pathways |
| 4 | Magnesium-protoporphyrin IX monomethyl ester (oxidative) cyclase | Chlorophyll metabolism | Up | 1.53 | F2D891 | Biosynthesis of secondary metabolites |
| 5 | Adenylosuccinate lyase | Nucleotide metabolism | Up | 1.54 | A0A287P260 | Biosynthesis of secondary metabolites |
| 6 | Phenylacetaldehyde reductase | Catalytic activity | Up | 1.59 | F2DCH1 | Other pathways |
| 7 | Delta-1-pyrroline-5-carboxylate synthase | Amino acid metabolism | Up | 1.62 | A0A224MLP5 | Biosynthesis of amino acids |
| 8 | Allene oxide synthase | Phytohormone action | Up | 1.66 | F2CWX3 | Biosynthesis of secondary metabolites |
| 9 | Photosynthetic NDH subunit of lumenal location 2. chloroplastic | Photosynthesis | Up | 1.70 | F2CTW7 | Energy metabolism |
| 10 | Sanguinarine reductase | Oxidoreductase | Up | 1.73 | M0VI12 | Metabolic pathways |
| 11 | Ribonuclease | RNA processing | Up | 1.73 | O04393 | Other pathways |
| 12 | L-galactose dehydrogenase | Redox homeostasis | Up | 1.74 | F2CWR4 | Biosynthesis of secondary metabolites |
| 13 | Mg-chelatase | Phytohormone action | Up | 1.75 | Q94C01 | Biosynthesis of secondary metabolites |
| 14 | D-ribulose kinase | ATP binding | Up | 1.79 | F2CZJ1 | Carbon metabolism |
| 15 | Bifunctional aspartokinase/homoserine dehydrogenase (AK-HSDH) | Amino acid metabolism | Up | 1.79 | F2EJ93 | Biosynthesis of amino acids |
| 16 | Phospho-2-dehydro-3-deoxyheptonate aldolase | Amino acid metabolism | Up | 1.81 | F2D2N1 | Biosynthesis of amino acids |
| 17 | Aconitase_C domain-containing protein | Clade-specific metabolism | Up | 1.84 | F2DE08 | Citrate cycle (TCA cycle) |
| 18 | 3-isopropylmalate dehydratase | Clade-specific metabolism | Up | 1.86 | F2DK00 | Biosynthesis of amino acids |
| 19 | Group3 late embryogenesis abundant protein | Stress response | Up | 1.87 | Q43478 | Metabolic pathways |
| 20 | Methyltransf_11 domain-containing protein | methyltransferase activity | Up | 1.98 | F2CR07 | Metabolic pathways |
| 21 | Annexin D7 | Stress response | Up | 1.99 | A0A287UPG1 | Other pathways |
| 22 | Purple acid phosphatase | Catalytic activity | Up | 2.45 | F2E6U9 | Metabolic pathways |
| 23 | 23 kda jasmonate-induced protein | Defense response | Up | 2.50 | P32024 | Other pathways |
| 24 | Myrosinase-binding protein 2-like | carbohydrate binding | Up | 3.14 | F2CV07 | Other pathways |
| 25 | Heat shock protein | Protein homeostasis | Up | 3.26 | F2DYT5 | Protein Processing |
| 26 | Glutathione S-transferase | Redox homeostasis | Up | 6.67 | M0V2H7 | Metabolic pathways |
| 27 | Translocase of chloroplast 34. chloroplastic-like | Protein translocation | Down | 0.06 | M0XFE2 | Other pathways |
| 28 | Ultraviolet-B receptor UVR8-like | External stimuli response | Down | 0.23 | F2CZC7 | Ubiquitin mediated proteolysis |
| 29 | Peroxidase (POX) | Response to oxidative stress | Down | 0.27 | M0Z9D1 | Biosynthesis of secondary metabolites |
| 30 | Short-chain dehydrogenase/reductase 2 | Oxidoreductase | Down | 0.39 | M0YJ05 | Metabolic pathways |
| 31 | Eceriferum-c | transferase | Down | 0.41 | M0XGE3 | Biosynthesis of secondary metabolites |
| 32 | Thaumatin-like protein (TLP5) | Defense response | Down | 0.43 | Q5MBN2 | Other pathways |
| 33 | Cysteine proteinase | Protein homeostasis | Down | 0.48 | F2DJL2 | Other pathways |
| 34 | Ferredoxin-NADP reductase | Oxidoreductase | Down | 0.52 | F2DFN1 | Energy metabolism |
| 35 | Nicotianamine synthase 9 | Nutrient uptake | Down | 0.53 | Q9XFB7 | Biosynthesis of secondary metabolites |
| 36 | Glycosyltransferase | Transferase | Down | 0.53 | F2CUZ3 | Metabolic pathways |
| 37 | Chitinase | Polysaccharide catabolic process | Down | 0.53 | A0A287IRA5 | Metabolic pathways |
| 38 | Germin-like protein | Stress response | Down | 0.54 | A0A287ERI6 | Other pathways |
| 39 | Subtilisin-like protease SBT3 | Protein homeostasis | Down | 0.54 | A0A287M050 | Other pathways |
| 40 | RHOMBOID-like protein | Protein homeostasis | Down | 0.55 | F2DXL3 | Other pathways |
| 41 | Pectinesterase | Cell wall organisation | Down | 0.55 | F2DNY3 | Metabolic pathways |
| 42 | Cytochrome P450 89A2 | Oxidoreductase | Down | 0.55 | F2DFD7 | Other pathways |
| 43 | DUF1338 domain-containing protein | Amino acid metabolism | Down | 0.55 | F2DXZ9 | Other pathways |
| 44 | Protein NRT1/PTR FAMILY | Solute transport | Down | 0.56 | F2DQ88 | Other pathways |
| 45 | Saccharopine dehydrogenase-like protein | Amino acid metabolism | Down | 0.56 | K9JGW1 | Biosynthesis of amino acids |
| 46 | Legumain | Multi-process regulation | Down | 0.57 | B4ESE3 | Metabolic pathways |
| 47 | Non-specific lipid transfer protein GPI-anchored 1 | Cell wall organisation | Down | 0.57 | F2CSC9 | Other pathways |
| 48 | Delta tonoplast intrinsic protein 2 | Solute transport | Down | 0.57 | B8R6A6 | Other pathways |
| 49 | Putative lysine/histidine transporter | Solute transport | Down | 0.57 | J7RA72 | ABC transporters |
| 50 | Abhydrolase_3 domain-containing protein | Hydrolase activity | Down | 0.58 | A0A287KEW5 | Metabolic pathways |
| 51 | Quinolinate phosphoribosyltransferase | NAD/NADP biosynthesis | Down | 0.58 | F2ECA5 | Metabolic pathways |
| 52 | Dirigent protein | plant secondary metabolism | Down | 0.60 | F2CQ16 | Other pathways |
| 53 | Lipase_3 domain-containing protein | Lipid metabolism | Down | 0.60 | A0A287K7Y5 | Other pathways |
| 54 | Topless-related protein 1 | Phytohormone action | Down | 0.61 | A0A287DXX3 | Other pathways |
| 55 | Catalase 2 | Redox homeostasis | Down | 0.61 | F2DBE3 | Biosynthesis of secondary metabolites |
| 56 | Acyl-activating enzyme 16. chloroplastic isoform X1 | Lipid metabolism | Down | 0.61 | A0A287SYP5 | Metabolic pathways |
| 57 | Lipoxygenase | Phytohormone action | Down | 0.62 | F2DFM0 | Metabolic pathways |
| 58 | Pectate_lyase_3 domain-containing protein | hydrolase | Down | 0.62 | F2D1A5 | Metabolic pathways |
| 59 | NAD(P)-binding Rossmann-fold superfamily protein | Protein neddylation | Down | 0.62 | F2DD18 | Biosynthesis of secondary metabolites |
| 60 | Haloacid dehalogenase-like hydrolase domain-containing protein Sgpp | Carbohydrate metabolism | Down | 0.65 | F2DPD5 | Other pathways |
| 61 | Beta-hexosaminidase | Protein modification | Down | 0.66 | F2E7M4 | Metabolic pathways |
| 62 | Beta-glucosidase | Hydrolase | Down | 0.66 | M0UZF4 | Biosynthesis of secondary metabolites |
| Gene | Forward and Reverse Primer 5′-3′ | Product Size (bp) |
|---|---|---|
| Pathogenesis-related protein (HvPR) | F: 5′-CTGCTGAAGGAGGTGGAGAC-3′ R: 5′-CCTCGAAGAGCTTGTTCACC-3′ | 117 |
| Delta-1-pyrroline-5-carboxylate synthase (HvP5CS) | F: 5′-GAGACAAGTCCCGTGTTGGT-3′ R: 5′-CCCACGGAGAACCTTAACAA-3′ | 134 |
| Group3 late embryogenesis abundant protein (HvLEA3) | F: 5′-GGTGGAGACGACCCAAGTTA-3′ R: 5′-GTGACAGCCTCGCTTTTCTC-3′ | 135 |
| Annexin D7 (HvANN7) | F: 5′-CCCTATCACCAAGGACCTCA-3′ R: 5′-GGGGCAAGTGAAACACCTTA-3′ | 85 |
| Heat shock protein (HvHSP) | F: 5′-CAAGATCACCATCACCAACG -3′ R: 5′-TAGTTCTCCAGCGCGTTCTT-3′ | 138 |
| Glutathione S-transferase (HvGST) | F: 5′-AAGATGTTCGTGGGCAAAAC-3′ R: 5′-AGGGCCACCTCTAGCTTCTC-3′ | 86 |
| Peroxidase (HvPRX) | F: 5′-TTCGACAAGAAGCAGCTGAA-3′ R: 5′-GTAGATGTGGTCGCGGAAGT-3′ | 99 |
| Thaumatin-like protein (HvTLP5) | F: 5′-ACTGCCCGCCAACATACTAC-3′ R: 5′-GTGCTGGTCTGGTCATCCTT-3′ | 85 |
| Chitinase IV precursor (HvCHI4) | F: 5′-GGACCAATTCCAGCTACTGC-3′ R: 5′-GACTTGATCCCGTCGAAGAA-3′ | 121 |
| Germin-like protein 5-1 (HvGLP) | F: 5′-TTCAGCATCCATCCAACAAG-3′ R: 5′-TGATCTGGCTGCTGAGAGTG-3′ | 121 |
| Protein NRT1/PTR FAMILY (HvNRT1) | F: 5′-TTCAGTCTCTCCGGCTTGAT-3′ R: 5′-GGAGCATCGTCTCCGAGTAG-3′ | 93 |
| Topless-related protein 1 (HvTRP1) | F: 5′-CGGCAGTGTTTATGTCATGG-3′ R: 5′-GCTCCATTAGGAGGTGTGGA-3′ | 108 |
| Catalase 2 (HvCAT2) | F: 5′-CATGTGATGGATGGATCTGC-3′ R: 5′-TCGACCACATGATCCACAGT-3′ | 123 |
| Lipoxygenase (HvLOX) | F: 5′-ACGACGTCCAAGTCGTAACC-3′ R: 5′-CACGTCCTCCAGGAACATCT-3′ | 116 |
| Actin (HvActin) | F: 5′-CGACAATGGAACCGGAATG-3′ R: 5′-CCCTTGGCGCATCATCTC-3′ | 56 |
| Parameters | Value |
|---|---|
| Clay (%) | 5.50 |
| Silt (%) | 8.30 |
| Sand (%) | 84.40 |
| Soil texture | Sandy |
| pH | 7.50 |
| EC (dS m–1) | 4.02 |
| Organic matter (%) | 0.905 |
| Total organic carbon (%) | 0.525 |
| Total N (g kg–1 soil) | 0.28 |
| Available P (mg kg–1 soil) | 4.92 |
| Exchange K (mg kg–1 soil) | 292 |
| Total coliforms (MPN g DW–1 soil) | 24 × 102 |
| Faecal coliforms (MPN g DW–1 soil) | <0.3 |
| Escherichia coli (MPN g DW–1 soil) | <0.3 |
| Faecal Streptococci (MPN g DW–1 soil) | 39 × 102 |
| Parameters | Value |
|---|---|
| Total organic carbon (%) | 18.58 |
| Total N (%) | 1.21 |
| C/N | 15.36 |
| P (%) | 0.54 |
| K (%) | 0.95 |
| Ca (%) | 8.18 |
| Mg (%) | 1.05 |
| Na (%) | 0.42 |
| Alkalinity (% CaCO3) | 11.50 |
| Zn (mg kg–1 DW compost) | 70.10 |
| Fe (g kg–1 DW compost) | 70 |
| Mn (mg kg–1 DW compost) | 130 |
| Cu (mg kg–1 DW compost) | 11.60 |
| Cd (mg kg–1 DW compost) | 0.20 |
| Pb (mg kg–1 DW compost) | 4.15 |
| Cr (mg kg–1 DW compost) | 11.50 |
| Ni (mg kg–1 DW compost) | 5.88 |
| Total coliforms (MPN g DW−1 compost) | 143.33 ± 5.77 |
| Faecal coliforms (MPN g DW−1 compost) | 120 ± 17.32 |
| Escherichia coli (MPN g DW−1 compost) | 114 ± 23.79 |
| Faecal Streptococci (MPN g DW−1 compost) | 114.33 ± 23.8 |
| Salmonella spp. (MPN g DW−1 compost) | <0.3 |
| Shigella spp. (MPN g DW−1 compost) | <0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghouili, E.; Abid, G.; Jebara, M.; Nefissi Ouertani, R.; de Oliveira, A.C.; El Ayed, M.; Muhovski, Y. Proteomic Analysis of Barley (Hordeum vulgare L.) Leaves in Response to Date Palm Waste Compost Application. Plants 2022, 11, 3287. https://doi.org/10.3390/plants11233287
Ghouili E, Abid G, Jebara M, Nefissi Ouertani R, de Oliveira AC, El Ayed M, Muhovski Y. Proteomic Analysis of Barley (Hordeum vulgare L.) Leaves in Response to Date Palm Waste Compost Application. Plants. 2022; 11(23):3287. https://doi.org/10.3390/plants11233287
Chicago/Turabian StyleGhouili, Emna, Ghassen Abid, Moez Jebara, Rim Nefissi Ouertani, Ana Caroline de Oliveira, Mohamed El Ayed, and Yordan Muhovski. 2022. "Proteomic Analysis of Barley (Hordeum vulgare L.) Leaves in Response to Date Palm Waste Compost Application" Plants 11, no. 23: 3287. https://doi.org/10.3390/plants11233287