
Evidence from Phylogenomics and Morphology Provide Insights into the Phylogeny, Plastome Evolution, and Taxonomy of Kitagawia
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Phylogenetic Analyses
2.2. Comparative Analyses of Plastomes
2.2.1. Plastome Features of Kitagawia and Related Taxa
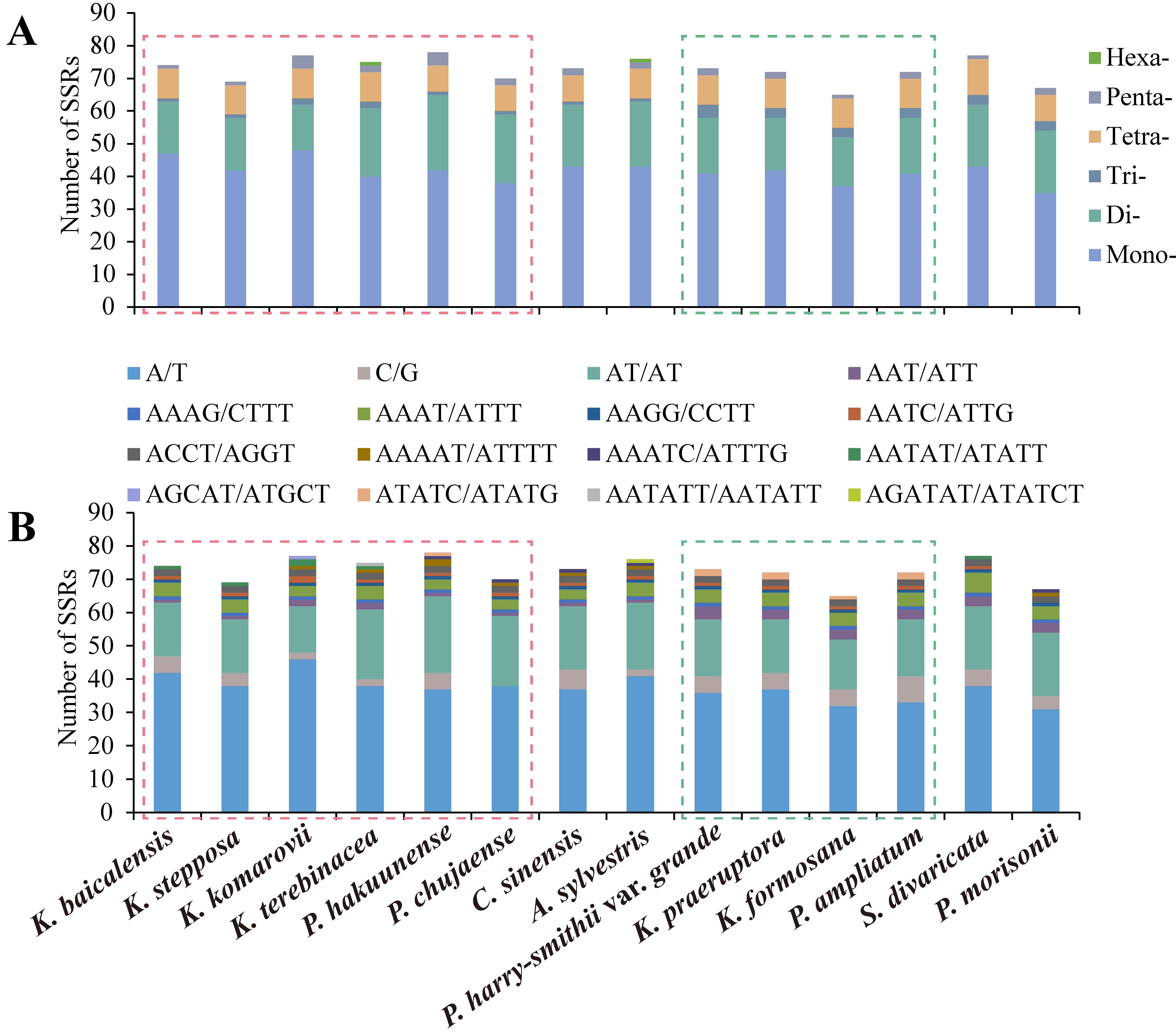
2.2.2. Simple Sequence Repeat Analysis
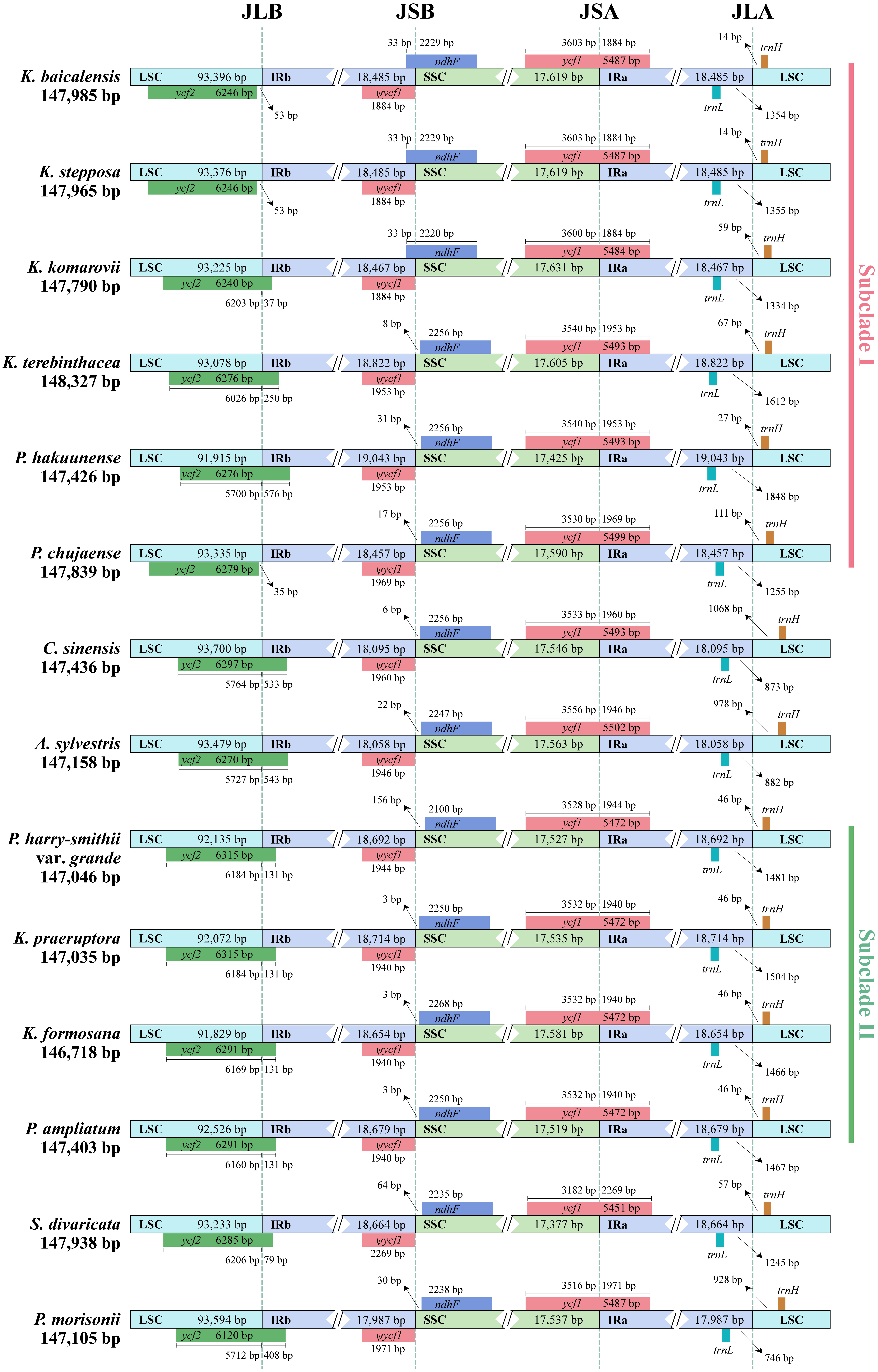
2.2.3. IR Border Analysis
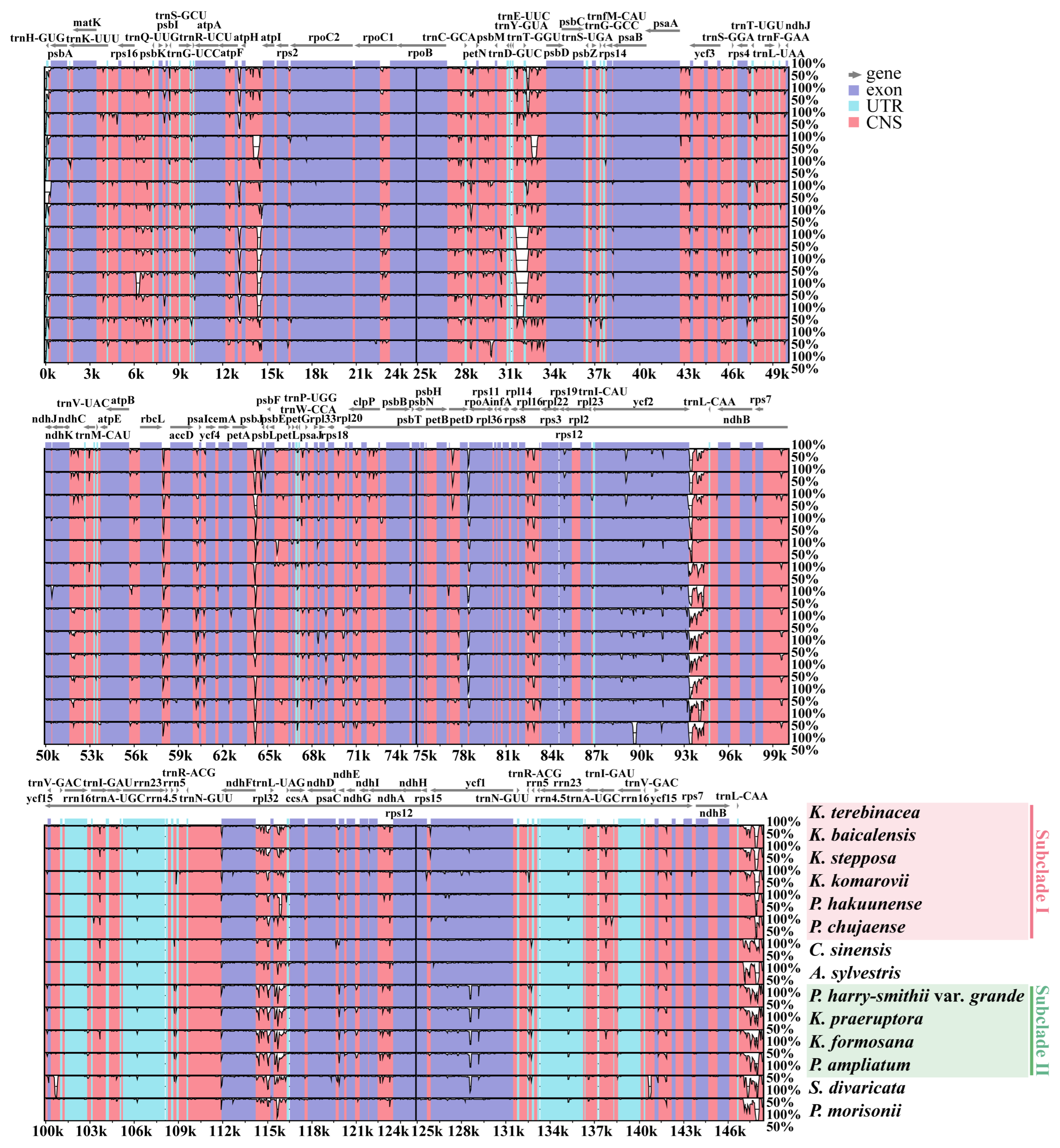
2.2.4. DNA Rearrangement and Sequence Divergence Analyses
2.3. Morphological Analyses
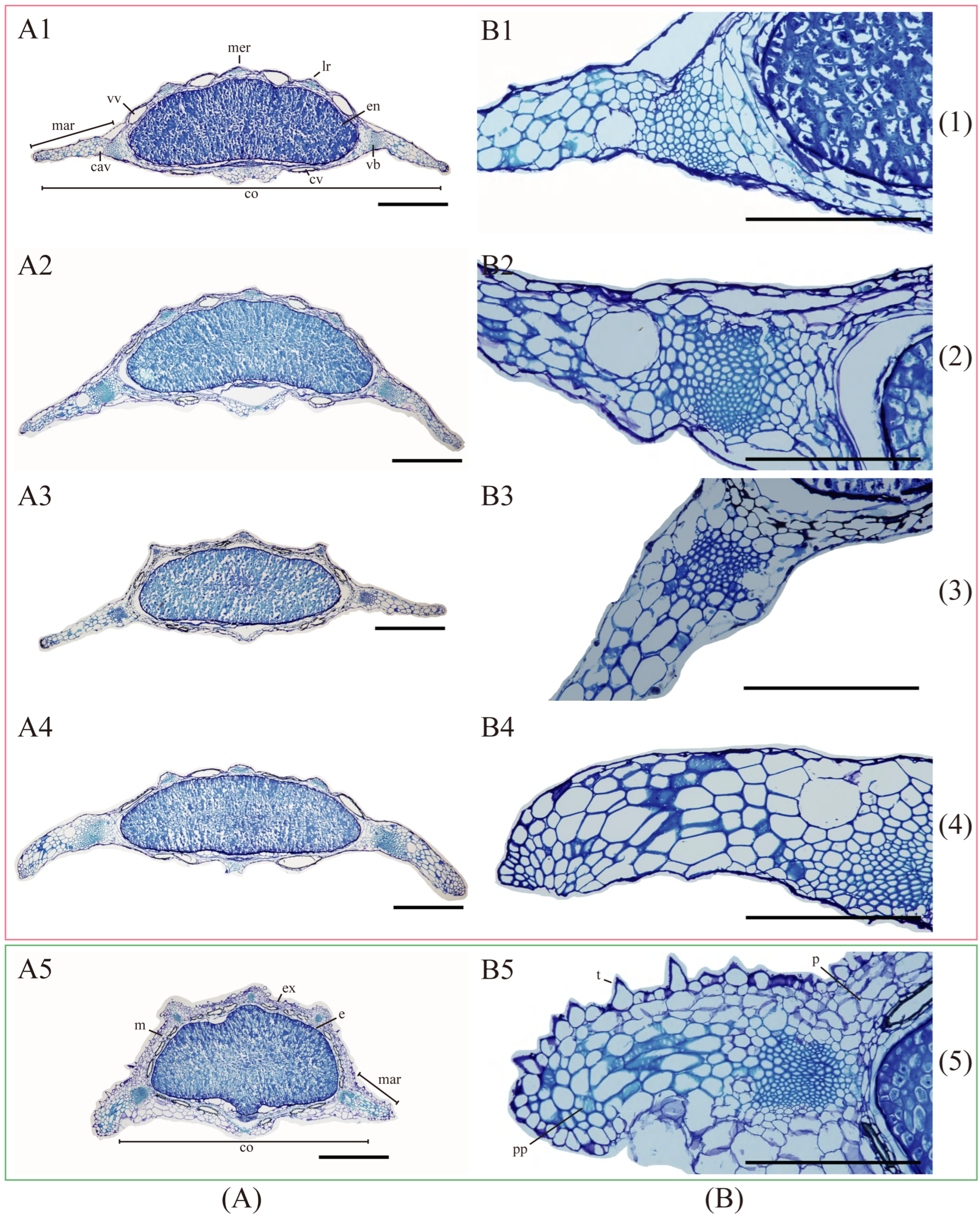
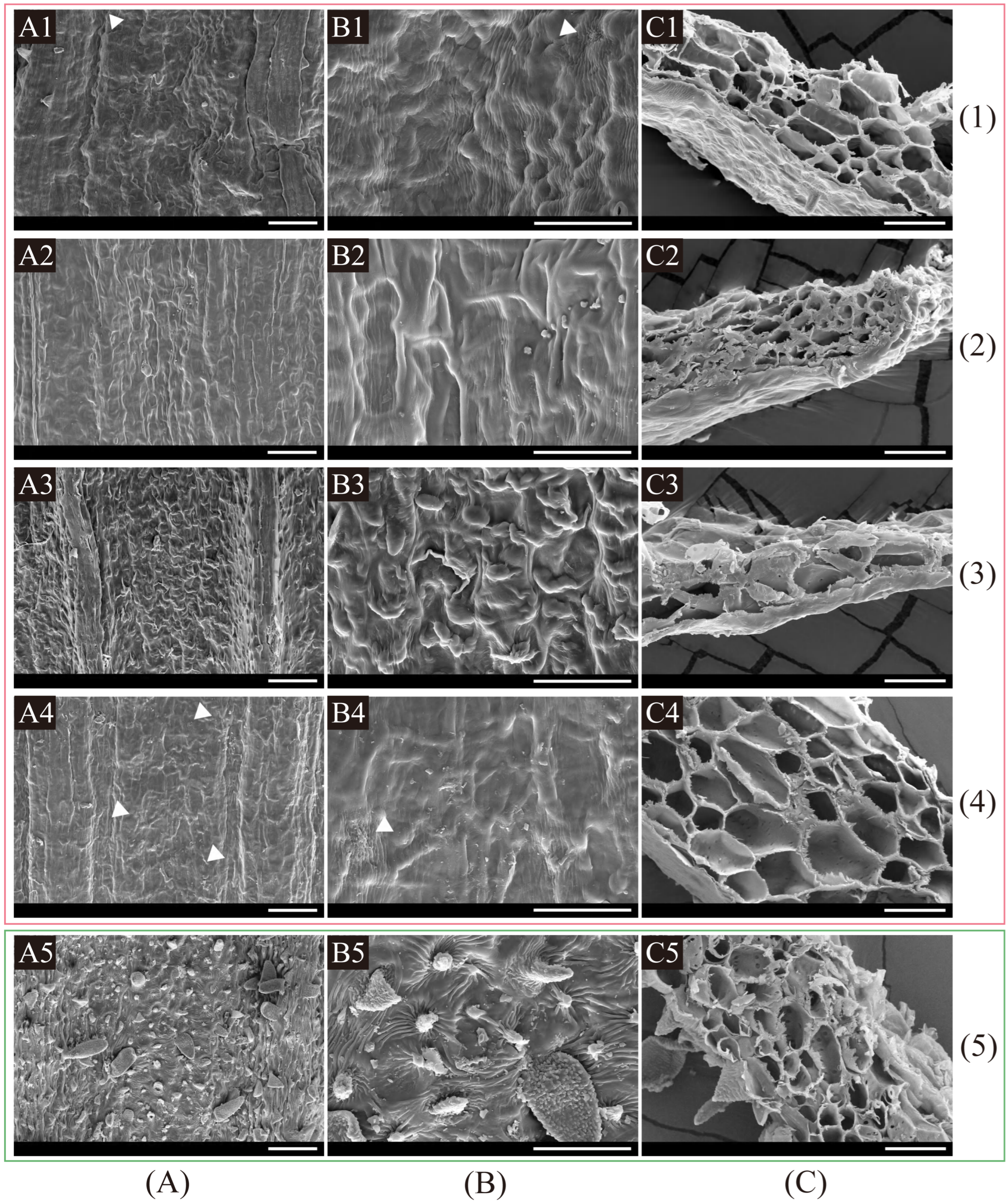
2.3.1. Fruit Anatomical and Micromorphological Examination
2.3.2. Comparison of General Morphological Characteristics
3. Discussion
3.1. Phylogenetic Position and Non-Monophyly of Kitagawia
3.2. Plastome Structure and Evolution and Their Phylogenetic Implications
3.3. Morphological Delimitations between Kitagawia and Related Taxa
3.4. Taxonomic Suggestions for Six Species from Kitagawia and Peucedanum Sensu Lato
3.5. Topologic Incongruence between Phylogenies based on Plastome and NrDNA Sequences
4. Materials and Methods
4.1. Taxon Sampling
4.2. DNA Extraction, Sequencing, Assembly, and Annotation
4.3. Phylogenetic Analyses
4.4. Comparative Analyses of Plastomes
4.5. Morphological Analyses
4.5.1. Fruit Anatomical and Micromorphological Characteristics
4.5.2. General Morphological Characteristics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pimenov, M.G.; Leonov, M.V.; Constance, L. The Genera of the Umbelliferae: A Nomenclator; Royal Botanic Gardens: Kew, UK, 1993; ISBN 978-0-947643-58-4. [Google Scholar]
- Shneyer, V.S.; Kutyavina, N.G.; Pimenov, M.G. Systematic Relationships within and between Peucedanum and Angelica (Umbelliferae–Peucedaneae) Inferred from Immunological Studies of Seed Proteins. Plant Syst. Evol. 2003, 236, 175–194. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, W.C.; Gong, X.; Liu, Z.W. Leaf Epidermal Morphology in Peucedanum L. (Umbelliferae) from China. Acta Bot. Gallica 2014, 161, 21–31. [Google Scholar] [CrossRef]
- Plunkett, G.M.; Pimenov, M.G.; Reduron, J.-P.; Kljuykov, E.V.; van Wyk, B.-E.; Ostroumova, T.A.; Henwood, M.J.; Tilney, P.M.; Spalik, K.; Watson, M.F.; et al. Apiaceae. In Flowering Plants. Eudicots: Apiales, Gentianales (Except Rubiaceae); Kadereit, J.W., Bittrich, V., Eds.; The Families and Genera of Vascular Plants; Springer International Publishing: Cham, Switzerland, 2018; Volume 15, pp. 9–206. ISBN 978-3-319-93605-5. [Google Scholar]
- Pimenov, M.G.; Ostroumova, T.A.; Degtjareva, G.V.; Samigullin, T.H. Sillaphyton, a New Genus of the Umbelliferae, Endemic to the Korean Peninsula. Bot. Pac. 2016, 5, 31–41. [Google Scholar] [CrossRef] [Green Version]
- Brullo, C.; Brullo, S.; Downie, S.R.; Danderson, C.A.; del Galdo, G.G. Siculosciadium, a New Monotypic Genus of Apiaceae from Sicily. Ann. Mo. Bot. Gard. 2013, 99, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Winter, P.J.D.; Magee, A.R.; Phephu, N.; Tilney, P.M.; Downie, S.R.; van Wyk, B.E. A New Generic Classification for African Peucedanoid Species (Apiaceae). Taxon 2008, 57, 347–364. [Google Scholar] [CrossRef]
- Spalik, K.; Reduron, J.-P.; Downie, S.R. The Phylogenetic Position of Peucedanum Sensu Lato and Allied Genera and Their Placement in Tribe Selineae (Apiaceae, Subfamily Apioideae). Plant Syst. Evol. 2004, 243, 189–210. [Google Scholar] [CrossRef]
- Ostroumova, T.A.; Pimenov, M.G.; Degtjareva, G.V.; Samigullin, T.H. Taeniopetalum Vis. (Apiaceae), a Neglected Segregate of Peucedanum, L., Supported as a Remarkable Genus by Morphological and Molecular Data. Skvortsovia 2016, 3, 20–44. [Google Scholar]
- Pimenov, M.G. Kitagawia—A New Asiatic Genus of the Family Umbelliferae. Bot. Zhurnal 1986, 71, 942–949. [Google Scholar]
- Sheh, M.L. Peucedanum. In Flora Reipublicae Popularis Sinicae; Science Press: Beijing, China, 1992; Volume 55, pp. 127–158. [Google Scholar]
- Sheh, M.L.; Watson, M.F. Peucedanum Linnaeus. In Flora of China; Science Press & Missouri Botanical Garden Press: Beijing, China; St. Louis, MO, USA, 2005; Volume 14, pp. 182–192. [Google Scholar]
- Yamazaki, T. Umbelliferae in Japan II. J. Jpn. Bot. 2001, 76, 283–285. [Google Scholar]
- Chang, C.S.; Kim, H.; Chang, K.S. Provisional Checklist of Vascular Plants for the Korea Peninsula Flora (KPF); Designpost: Pajo, Philippines, 2014; ISBN 978-89-968648-2-0. [Google Scholar]
- Kim, K.; Kim, C.S.; Oh, S.H.; Park, C.W. A New Species of Peucedanum (Apiaceae) from Korea. Phytotaxa 2019, 393, 75. [Google Scholar] [CrossRef]
- Kim, K.; Suh, H.-J.; Song, J.-H. Two New Endemic Species, Peucedanum miroense and P. tongkangense (Apiaceae), from Korea. PhytoKeys 2022, 210, 35–52. [Google Scholar] [CrossRef]
- Pimenov, M.G.; Ostroumova, T.A. Umbelliferae of Russia; Association of Scientific Publications: Moscow, Russia, 2012; ISBN 978-5-87317-813-1. [Google Scholar]
- Pimenov, M.G. Updated Checklist of Chinese Umbelliferae: Nomenclature, Synonymy, Typification, Distribution. Turczaninowia 2017, 20, 106–239. [Google Scholar] [CrossRef] [Green Version]
- Drude, O. Umbelliferae. In Die Natürlichen Pflanzenfamilien; Wilhelm Engelman: Leipzig, Germany, 1898; Volume 3, pp. 63–150, 271. [Google Scholar]
- Kljuykov, E.V.; Zakharova, E.A.; Ostroumova, T.A.; Tilney, P.M. Most Important Carpological Anatomical Characters in the Taxonomy of Apiaceae. Bot. J. Linn. Soc. 2021, 195, 532–544. [Google Scholar] [CrossRef]
- Liu, M.; van Wyk, B.-E.; Tilney, P.M. The Taxonomic Value of Fruit Structure in the Subfamily Saniculoideae and Related African Genera (Apiaceae). Taxon 2003, 52, 261–270. [Google Scholar] [CrossRef]
- Morison, R. Plantarum Umbelliferarum Distributio Nova; Theatro Sheldoniano: Oxford, UK, 1672. [Google Scholar]
- Ostroumova, T.A. Fruit Micromorphology in the Umbelliferae of the Russian Far East. Bot. Pac. J. Plant Sci. Conserv. 2018, 7, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Ostroumova, T.A. Fruit Micromorphology in the Umbelliferae in Siberia and Patterns of Morphological Diversity in the Family. Probl. Bot. South Sib. Mong. 2020, 19, 44–48. [Google Scholar] [CrossRef]
- Ostroumova, T.A. Fruit Micromorphology of Siberian Apiaceae and Its Value for Taxonomy of the Family. Turczaninowia 2021, 24, 120–143. [Google Scholar] [CrossRef]
- Lee, C.; Kim, J.; Darshetkar, A.M.; Choudhary, R.K.; Park, S.H.; Lee, J.; Choi, S. Mericarp Morphology of the Tribe Selineae (Apiaceae, Apioideae) and Its Taxonomic Implications in Korea. Bangladesh J. Plant Taxon. 2018, 25, 175–186. [Google Scholar] [CrossRef]
- Downie, S.R.; Watson, M.F.; Spalik, K.; Katz-Downie, D.S. Molecular Systematics of Old World Apioideae (Apiaceae): Relationships among Some Members of Tribe Peucedaneae Sensu Lato, the Placement of Several Island-Endemic Species, and Resolution within the Apioid Superclade. Can. J. Bot. 2000, 78, 506–528. [Google Scholar] [CrossRef]
- Downie, S.R.; Plunkett, G.M.; Watson, M.F.; Spalik, K.; Katz-Downie, D.S.; Valiejo-Roman, C.M.; Terentieva, E.I.; Troitsky, A.V.; Lee, B.-Y.; Lahham, J.; et al. Tribes and Clades within Apiaceae Subfamily Apioideae: The Contribution of Molecular Data. Edinb. J. Bot. 2001, 58, 301–330. [Google Scholar] [CrossRef]
- Feng, T.; Downie, S.R.; Yu, Y.; Zhang, X.M.; Chen, W.W.; He, X.J.; Liu, S. Molecular Systematics of Angelica and Allied Genera (Apiaceae) from the Hengduan Mountains of China Based on NrDNA ITS Sequences: Phylogenetic Affinities and Biogeographic Implications. J. Plant Res. 2009, 122, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Downie, S.R.; Spalik, K.; Katz-Downie, D.S.; Reduron, J.P. Major Clades within Apiaceae Subfamily Apioideae as Inferred by Phylogenetic Analysis of NrDNA ITS Sequences. Plant Divers. Evol. 2010, 128, 111. [Google Scholar] [CrossRef] [Green Version]
- Pimenov, M.; Degtjareva, G.; Ostroumova, T.; Samigullin, T.; Averyanov, L.V. Xyloselinum laoticum (Umbelliferae), a New Species from Laos, and Taxonomic Placement of the Genus in the Light of NrDNA ITS Sequence Analysis. Phytotaxa 2016, 244, 248. [Google Scholar] [CrossRef]
- Zhou, J.; Gong, X.; Downie, S.R.; Peng, H. Towards a More Robust Molecular Phylogeny of Chinese Apiaceae Subfamily Apioideae: Additional Evidence from NrDNA ITS and CpDNA Intron (Rpl16 and Rps16) Sequences. Mol. Phylogenet. Evol. 2009, 53, 56–68. [Google Scholar] [CrossRef]
- Zhou, J.; Gao, Y.Z.; Wei, J.; Liu, Z.W.; Downie, S.R. Molecular Phylogenetics of Ligusticum (Apiaceae) Based on NrDNA ITS Sequences: Rampant Polyphyly, Placement of the Chinese Endemic Species, and a Much-Reduced Circumscription of the Genus. Int. J. Plant Sci. 2020, 181, 306–323. [Google Scholar] [CrossRef]
- Corriveau, J.L.; Coleman, A.W. Rapid Screening Method to Detect Potential Biparental Inheritance of Plastid DNA and Results for over 200 Angiosperm Species. Am. J. Bot. 1988, 75, 1443–1458. [Google Scholar] [CrossRef]
- Mandel, J.R.; Ramsey, A.J.; Holley, J.M.; Scott, V.A.; Mody, D.; Abbot, P. Disentangling Complex Inheritance Patterns of Plant Organellar Genomes: An Example from Carrot. J. Hered. 2020, 111, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Erixon, P.; Oxelman, B. Whole-Gene Positive Selection, Elevated Synonymous Substitution Rates, Duplication, and Indel Evolution of the Chloroplast ClpP1 Gene. PLoS ONE 2008, 3, e1386. [Google Scholar] [CrossRef] [Green Version]
- Korpelainen, H. The Evolutionary Processes of Mitochondrial and Chloroplast Genomes Differ from Those of Nuclear Genomes. Sci. Nat. 2004, 91, 505–518. [Google Scholar] [CrossRef]
- Gou, W.; Jia, S.B.; Price, M.; Guo, X.L.; Zhou, S.D.; He, X.J. Complete Plastid Genome Sequencing of Eight Species from Hansenia, Haplosphaera and Sinodielsia (Apiaceae): Comparative Analyses and Phylogenetic Implications. Plants 2020, 9, 1523. [Google Scholar] [CrossRef]
- Guo, X.L.; Zheng, H.Y.; Price, M.; Zhou, S.D.; He, X.J. Phylogeny and Comparative Analysis of Chinese Chamaesium Species Revealed by the Complete Plastid Genome. Plants 2020, 9, 965. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yu, Y.; Liu, Y.M.; Xie, D.F.; He, X.J.; Zhou, S.D. Comparative Chloroplast Genomics of Fritillaria (Liliaceae), Inferences for Phylogenetic Relationships between Fritillaria and Lilium and Plastome Evolution. Plants 2020, 9, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Cai, J.; Qin, H.H.; Price, M.; Zhang, Z.; Yu, Y.; Xie, D.F.; He, X.J.; Zhou, S.D.; Gao, X.F. Phylogeny, Age, and Evolution of Tribe Lilieae (Liliaceae) Based on Whole Plastid Genomes. Front. Plant Sci. 2021, 12, 699226. [Google Scholar] [CrossRef]
- Parks, M.; Cronn, R.; Liston, A. Increasing Phylogenetic Resolution at Low Taxonomic Levels Using Massively Parallel Sequencing of Chloroplast Genomes. BMC Biol. 2009, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Ren, T.; Li, Z.X.; Xie, D.F.; Gui, L.J.; Peng, C.; Wen, J.; He, X.J. Plastomes of Eight Ligusticum Species: Characterization, Genome Evolution, and Phylogenetic Relationships. BMC Plant Biol. 2020, 20, 519. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Wicke, S.; Wang, H.; Jin, J.J.; Chen, S.Y.; Zhang, S.D.; Li, D.Z.; Yi, T.S. Plastid Genome Evolution in the Early-Diverging Legume Subfamily Cercidoideae (Fabaceae). Front. Plant Sci. 2018, 9, 138. [Google Scholar] [CrossRef] [Green Version]
- Yao, G.; Jin, J.J.; Li, H.T.; Yang, J.B.; Mandala, V.S.; Croley, M.; Mostow, R.; Douglas, N.A.; Chase, M.W.; Christenhusz, M.J.M. Plastid Phylogenomic Insights into the Evolution of Caryophyllales. Mol. Phylogenet. Evol. 2019, 134, 74–86. [Google Scholar] [CrossRef]
- Wen, J.; Xie, D.F.; Price, M.; Ren, T.; Deng, Y.Q.; Gui, L.J.; Guo, X.L.; He, X.J. Backbone Phylogeny and Evolution of Apioideae (Apiaceae): New Insights from Phylogenomic Analyses of Plastome Data. Mol. Phylogenet. Evol. 2021, 161, 107183. [Google Scholar] [CrossRef]
- Liu, C.K.; Lei, J.Q.; Jiang, Q.P.; Zhou, S.D.; He, X.J. The Complete Plastomes of Seven Peucedanum Plants: Comparative and Phylogenetic Analyses for the Peucedanum Genus. BMC Plant Biol. 2022, 22, 101. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.F.; Yu, H.X.; Price, M.; Xie, C.; Deng, Y.Q.; Chen, J.P.; Yu, Y.; Zhou, S.D.; He, X.J. Phylogeny of Chinese Allium Species in Section Daghestanica and Adaptive Evolution of Allium (Amaryllidaceae, Allioideae) Species Revealed by the Chloroplast Complete Genome. Front. Plant Sci. 2019, 10, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Wang, N.; Cheng, G.; Shu, X.; Wang, T.; Zhuang, W.; Lu, R.; Wang, Z. Comparative Chloroplast Genomes of Four Lycoris Species (Amaryllidaceae) Provides New Insight into Interspecific Relationship and Phylogeny. Biology 2021, 10, 715. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Xie, D.; Peng, C.; Gui, L.; Price, M.; Zhou, S.; He, X. Molecular Evolution and Phylogenetic Relationships of Ligusticum (Apiaceae) Inferred from the Whole Plastome Sequences. BMC Ecol. Evol. 2022, 22, 55. [Google Scholar] [CrossRef] [PubMed]
- Gou, W.; Guo, X.L.; Zhou, S.D.; He, X.J. Phylogeny and Taxonomy of Meeboldia, Sinodielsia and Their Relatives (Apiaceae: Apioideae) Inferred from NrDNA ITS, Plastid DNA Intron (Rpl16 and Rps16) Sequences and Morphological Characters. Phytotaxa 2021, 482, 121–142. [Google Scholar] [CrossRef]
- Lyskov, D.F.; Samigullin, T.H.; Pimenov, M.G. Molecular and Morphological Data Support the Transfer of the Monotypic Iranian Genus Alococarpum to Prangos (Apiaceae). Phytotaxa 2017, 299, 223–233. [Google Scholar] [CrossRef]
- Pimenov, M.; Degtjareva, G.; Ostroumova, T.; Samigullin, T.; Zakharova, E. Polyphyletic Trachyspermum (Umbelliferae) Revisited: A Contribution of Molecular and Carpological Data. Plant Biosyst. 2022, 156, 722–742. [Google Scholar] [CrossRef]
- Xiao, Y.P.; Guo, X.L.; Price, M.; Gou, W.; Zhou, S.D.; He, X.J. New Insights into the Phylogeny of Sinocarum (Apiaceae, Apioideae) Based on Morphological and Molecular Data. Phytokeys 2021, 175, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.X.; Guo, X.L.; Price, M.; Zhou, S.D.; He, X.J. Phylogenetic Position of Ligusticopsis (Apiaceae, Apioideae): Evidence from Molecular Data and Carpological Characters. AoB Plants 2022, 14, plac008. [Google Scholar] [CrossRef] [PubMed]
- Mohanta, T.K.; Mishra, A.K.; Khan, A.; Hashem, A.; Abd_Allah, E.F.; Al-Harrasi, A. Gene Loss and Evolution of the Plastome. Genes 2020, 11, 1133. [Google Scholar] [CrossRef] [PubMed]
- Abdullah; Mehmood, F.; Heidari, P.; Rahim, A.; Ahmed, I.; Poczai, P. Pseudogenization of the Chloroplast Threonine (TrnT-GGU) Gene in the Sunflower Family (Asteraceae). Sci. Rep. 2021, 11, 21122. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.-M.; Staub, J.E.; Chen, J.-F. Molecular Phylogeny of Cucumis Species as Revealed by Consensus Chloroplast SSR Marker Length and Sequence Variation. Genome 2006, 49, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Powell, W.; Morgante, M.; Andre, C.; McNicol, J.W.; Machray, G.C.; Doyle, J.J.; Tingey, S.V.; Rafalski, J.A. Hypervariable Microsatellites Provide a General Source of Polymorphic DNA Markers for the Chloroplast Genome. Curr. Biol. 1995, 5, 1023–1029. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Panesar, P.S.; Bera, M.B.; Kaur, V. Simple Sequence Repeat Markers in Genetic Divergence and Marker-Assisted Selection of Rice Cultivars: A Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xie, D.; He, X.; Yang, Y.; Li, X. Comparative Analysis of the Complete Chloroplast Genomes in Allium Section Bromatorrhiza Species (Amaryllidaceae): Phylogenetic Relationship and Adaptive Evolution. Genes 2022, 13, 1279. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Price, M.; Su, D.M.; Zhang, Z.; Yu, Y.; Xie, D.F.; Zhou, S.D.; He, X.J.; Gao, X.F. Phylogeny and Comparative Analysis for the Plastid Genomes of Five Tulipa (Liliaceae). BioMed Res. Int. 2021, 2021, 6648429. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Song, J.; Gao, H.; Zhu, Y.; Xu, J.; Pang, X.; Yao, H.; Sun, C.; Li, X.; Li, C.; et al. The Complete Chloroplast Genome Sequence of the Medicinal Plant Salvia miltiorrhiza. PLoS ONE 2013, 8, e57607. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.J.; Cheng, C.L.; Chang, C.C.; Wu, C.L.; Su, T.M.; Chaw, S.M. Dynamics and Evolution of the Inverted Repeat-Large Single Copy Junctions in the Chloroplast Genomes of Monocots. BMC Evol. Biol. 2008, 8, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yildirim, H.; Duman, H. Peucedanum guvenianum (Apiaceae), a New Species from West Anatolia, Turkey. Doga Turk. J. Bot. 2017, 41, 600–608. [Google Scholar] [CrossRef]
- Dunn, S.T. Carlesia Dunn. In Hooker’s Icones Plantarum; Ser. 4; Icones Plantarum; London, UK, 1902; Volume 28, p. 2739. [Google Scholar]
- Dihoru, G.; Comanescu, M.; Nicoara, R. Analysis of the Characters on Some Angelica Taxa. Rom. J. Biol.-Plant Biol. 2011, 56, 79–89. [Google Scholar]
- Randall, R.E.; Thornton, G. Peucedanum officinale L. J. Ecol. 1996, 84, 475–485. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, W.; Liu, M.; Liu, Z. Molecular Authentication of the Traditional Medicinal Plant Peucedanum praeruptorum and Its Substitutes and Adulterants by DNA-Barcoding Technique. Pharmacogn. Mag. 2014, 10, 385–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, M.T. Peucedanum L. In Flora of Taiwan, 2nd ed.; Huang, T.C., Ed.; Department of Botany, National Taiwan University: Taipei, Taiwan, 1993; Volume 3, pp. 1034–1037. [Google Scholar]
- Liou, T.N. Saposhnikovia Schischk. In Flora Plantarum Herbacearum Chinae Boreali-Orientalis; Science Press: Beijing, China, 1977; Volume 6, pp. 213–216. [Google Scholar]
- Nakai, T. Notulae ad Plantas Asiae Orientalis (XI). J. Jpn. Bot. 1939, 15, 740–741. [Google Scholar]
- Park, C.; Lee, B.; Song, J.-H.; Kim, K. Peucedanum L. In Flora of Korea Volume 5c Rosidae: Rhamnaceae to Apiaceae; National Institute of Biological Resources: Incheon, Republic of Korea, 2017; ISBN 978-89-6811-276-8. [Google Scholar]
- Stull, G.W.; Soltis, P.S.; Soltis, D.E.; Gitzendanner, M.A.; Smith, S.A. Nuclear Phylogenomic Analyses of Asterids Conflict with Plastome Trees and Support Novel Relationships among Major Lineages. Am. J. Bot. 2020, 107, 790–805. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J. Gene Trees and Species Trees: Molecular Systematics as One-Character Taxonomy. Syst. Bot. 1992, 17, 144–163. [Google Scholar] [CrossRef]
- Smith, S.A.; Moore, M.J.; Brown, J.W.; Yang, Y. Analysis of Phylogenomic Datasets Reveals Conflict, Concordance, and Gene Duplications with Examples from Animals and Plants. BMC Evol. Biol. 2015, 15, 150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelser, P.B.; Kennedy, A.H.; Tepe, E.J.; Shidler, J.B.; Nordenstam, B.; Kadereit, J.W.; Watson, L.E. Patterns and Causes of Incongruence between Plastid and Nuclear Senecioneae (Asteraceae) Phylogenies. Am. J. Bot. 2010, 97, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.X.; Zeng, C.X.; Li, D.Z. Complex Evolution in Arundinarieae (Poaceae: Bambusoideae): Incongruence between Plastid and Nuclear GBSSI Gene Phylogenies. Mol. Phylogenet. Evol. 2012, 63, 777–797. [Google Scholar] [CrossRef]
- Lavretsky, P.; Hernández-Baños, B.E.; Peters, J.L. Rapid Radiation and Hybridization Contribute to Weak Differentiation and Hinder Phylogenetic Inferences in the New World Mallard Complex (Anas spp.). Auk 2014, 131, 524–538. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.; Ezcurra, C.; Calviño, C.I. Chloroplast and ITS Phylogenies to Understand the Evolutionary History of Southern South American Azorella, Laretia and Mulinum (Azorelloideae, Apiaceae). Mol. Phylogenet. Evol. 2017, 108, 1–21. [Google Scholar] [CrossRef]
- Milne, R.I.; Abbott, R.J. The Origin and Evolution of Tertiary Relict Floras. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2002; Volume 38, pp. 281–314. ISBN 0065-2296. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols, A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Baldwin, B.G.; Markos, S. Phylogenetic Utility of the External Transcribed Spacer (ETS) of 18S–26S RDNA: Congruence of ETS and ITS Trees of Calycadenia (Compositae). Mol. Phylogenet. Evol. 1998, 10, 449–463. [Google Scholar] [CrossRef] [PubMed]
- Logacheva, M.D.; Valiejo-Roman, C.M.; Degtjareva, G.V.; Stratton, J.M.; Downie, S.R.; Samigullin, T.H.; Pimenov, M.G. A Comparison of NrDNA ITS and ETS Loci for Phylogenetic Inference in the Umbelliferae: An Example from Tribe Tordylieae. Mol. Phylogenet. Evol. 2010, 57, 471–476. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; DePamphilis, C.W.; Yi, T.S.; Li, D.Z. GetOrganelle: A Fast and Versatile Toolkit for Accurate de Novo Assembly of Organelle Genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.J.; Moore, M.J.; Li, D.Z.; Yi, T.S. PGA: A Software Package for Rapid, Accurate, and Flexible Batch Annotation of Plastomes. Plant Methods 2019, 15, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greiner, S.; Lehwark, P.; Bock, R. OrganellarGenomeDRAW (OGDRAW) Version 1.3.1: Expanded Toolkit for the Graphical Visualization of Organellar Genomes. Nucleic Acids Res. 2019, 47, W59–W64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An Integrated and Scalable Desktop Platform for Streamlined Molecular Sequence Data Management and Evolutionary Phylogenetics Studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J. MrModeltest v2. Program Distributed by the Author. Evolutionary Biology Centre, Uppsala University. Available online: https://github.com/nylander/MrModeltest2 (accessed on 8 October 2021).
- Rambaut, A.; Drummond, A. FigTree v1.4.4. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 November 2021).
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-Web: A Web Server for Microsatellite Prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiryousefi, A.; Hyvönen, J.; Poczai, P. IRscope: An Online Program to Visualize the Junction Sites of Chloroplast Genomes. Bioinformatics 2018, 34, 3030–3031. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple Alignment of Conserved Genomic Sequence with Rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational Tools for Comparative Genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brudno, M.; Malde, S.; Poliakov, A.; Do, C.B.; Couronne, O.; Dubchak, I.; Batzoglou, S. Glocal Alignment: Finding Rearrangements during Alignment. Bioinformatics 2003, 19, i54–i62. [Google Scholar] [CrossRef] [PubMed]
- Feder, N.; O’Brien, T.P. Plant Microtechnique: Some Principles and New Methods. Am. J. Bot. 1968, 55, 123–142. [Google Scholar] [CrossRef]
- Kljuykov, E.V.; Liu, M.; Ostroumova, T.A.; Pimenov, M.G.; Tilney, P.M.; van Wyk, B.-E.; van Staden, J. Towards a Standardised Terminology for Taxonomically Important Morphological Characters in the Umbelliferae. S. Afr. J. Bot. 2004, 70, 488–496. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Length (bp) | Number of Unique Genes | GC Content (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genome | LSC | SSC | IR | Total | PCG | tRNA | rRNA | Total | LSC | SSC | IR | |
| K. baicalensis | 147,985 | 93,396 | 17,619 | 18,485 | 114 | 80 | 30 | 4 | 37.5 | 35.9 | 31.0 | 44.6 |
| K. formosana | 146,718 | 91,829 | 17,581 | 18,654 | 113 | 80 | 29 | 4 | 37.6 | 36.1 | 31.0 | 44.5 |
| K. komarovii | 147,790 | 93,225 | 17,631 | 18,467 | 114 | 80 | 30 | 4 | 37.5 | 35.9 | 31.1 | 44.6 |
| K. praeruptora | 147,035 | 92,072 | 17,535 | 18,714 | 113 | 80 | 29 | 4 | 37.6 | 36.1 | 31.1 | 44.5 |
| K. stepposa | 147,965 | 93,376 | 17,619 | 18,485 | 114 | 80 | 30 | 4 | 37.5 | 36.0 | 31.0 | 44.6 |
| K. terebinthacea | 148,327 | 93,078 | 17,605 | 18,822 | 114 | 80 | 30 | 4 | 37.5 | 35.9 | 30.9 | 44.3 |
| P. ampliatum | 147,403 | 92,526 | 17,519 | 18,679 | 114 | 80 | 30 | 4 | 37.6 | 36.0 | 31.0 | 44.5 |
| P. chujaense | 147,839 | 93,335 | 17,590 | 18,457 | 114 | 80 | 30 | 4 | 37.4 | 35.9 | 30.8 | 44.6 |
| P. hakuunense | 147,426 | 91,915 | 17,425 | 19,043 | 114 | 80 | 30 | 4 | 37.5 | 35.9 | 30.9 | 44.4 |
| P. harry-smithii var. grande | 147,046 | 92,135 | 17,527 | 18,692 | 113 | 80 | 29 | 4 | 37.6 | 36.0 | 31.1 | 44.5 |
| P. morisonii | 147,105 | 93,594 | 17,537 | 17,987 | 114 | 80 | 30 | 4 | 37.6 | 36.1 | 31.1 | 44.8 |
| A. sylvestris | 147,158 | 93,479 | 17,563 | 18,058 | 114 | 80 | 30 | 4 | 37.5 | 35.9 | 31.0 | 44.8 |
| C. sinensis | 147,436 | 93,700 | 17,546 | 18,095 | 114 | 80 | 30 | 4 | 37.5 | 35.9 | 31.0 | 44.7 |
| S. divaricata | 147,938 | 93,233 | 17,377 | 18,664 | 114 | 80 | 30 | 4 | 37.5 | 35.9 | 30.8 | 44.6 |
| Taxa | K. baicalensis | K. stepposa | K. komarovii | K. terebinthacea | K. praeruptora |
|---|---|---|---|---|---|
| Mericarps | Strongly compressed dorsally | Strongly compressed dorsally | Strongly compressed dorsally | Strongly compressed dorsally | Slightly compressed dorsally |
| Dorsal rib shape | Filiform, slightly prominent | Filiform, slightly prominent | Filiform, slightly prominent | Filiform, slightly prominent | Filiform, prominent |
| Marginal rib shape | Winged | Winged | Winged | Winged | Narrowly winged |
| Vallecular vittae | 1 | 1 | 1 | 1 | 3–4 |
| Commissural vittae | 2 | 2 | 2 | 2 | 6 |
| Endosperm | Slightly concave | Slightly concave | Flat | Flat | Flat |
| Layers of mesocarp cells | 1–2 | 1–2 | 1–2 | 1–2 | 3–5 |
| Mesocarp parenchyma | Lignified parenchyma with pitted walls | Lignified parenchyma with pitted walls | Lignified parenchyma with pitted walls | Lignified parenchyma with pitted walls | Lignified parenchyma with pitted walls |
| Average of marw/fw (%) | 42.3 | 48.8 | 44.6 | 50.3 | 39.8 |
| Average of cow/fw (%) | 96.4 | 94.0 | 95.4 | 94.1 | 85.2 |
| Cell borders (outer surface of fruits) | Inconspicuous | Inconspicuous | Inconspicuous | Inconspicuous | Inconspicuous |
| Fruit surfaces | Undulate | Rugate | Tuberculate | Undulate | Raised |
| Cuticular foldings | Striate and rugulate | Smooth and striate | Smooth and striate | Striate and rugulate | Rugulate and tuberculate |
| Epicuticular wax | Absent | Absent | Absent | Absent | Absent |
| Inner fruit structure | Parenchyma cells with pitted walls | Parenchyma cells with pitted walls | Parenchyma cells with pitted walls | Parenchyma cells with pitted walls | Parenchyma cells with pitted walls |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, J.-Q.; Liu, C.-K.; Cai, J.; Price, M.; Zhou, S.-D.; He, X.-J. Evidence from Phylogenomics and Morphology Provide Insights into the Phylogeny, Plastome Evolution, and Taxonomy of Kitagawia. Plants 2022, 11, 3275. https://doi.org/10.3390/plants11233275
Lei J-Q, Liu C-K, Cai J, Price M, Zhou S-D, He X-J. Evidence from Phylogenomics and Morphology Provide Insights into the Phylogeny, Plastome Evolution, and Taxonomy of Kitagawia. Plants. 2022; 11(23):3275. https://doi.org/10.3390/plants11233275
Chicago/Turabian StyleLei, Jia-Qing, Chang-Kun Liu, Jing Cai, Megan Price, Song-Dong Zhou, and Xing-Jin He. 2022. "Evidence from Phylogenomics and Morphology Provide Insights into the Phylogeny, Plastome Evolution, and Taxonomy of Kitagawia" Plants 11, no. 23: 3275. https://doi.org/10.3390/plants11233275