
Increased Invasion Risk of Tagetes minuta L. in China under Climate Change: A Study of the Potential Geographical Distributions
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
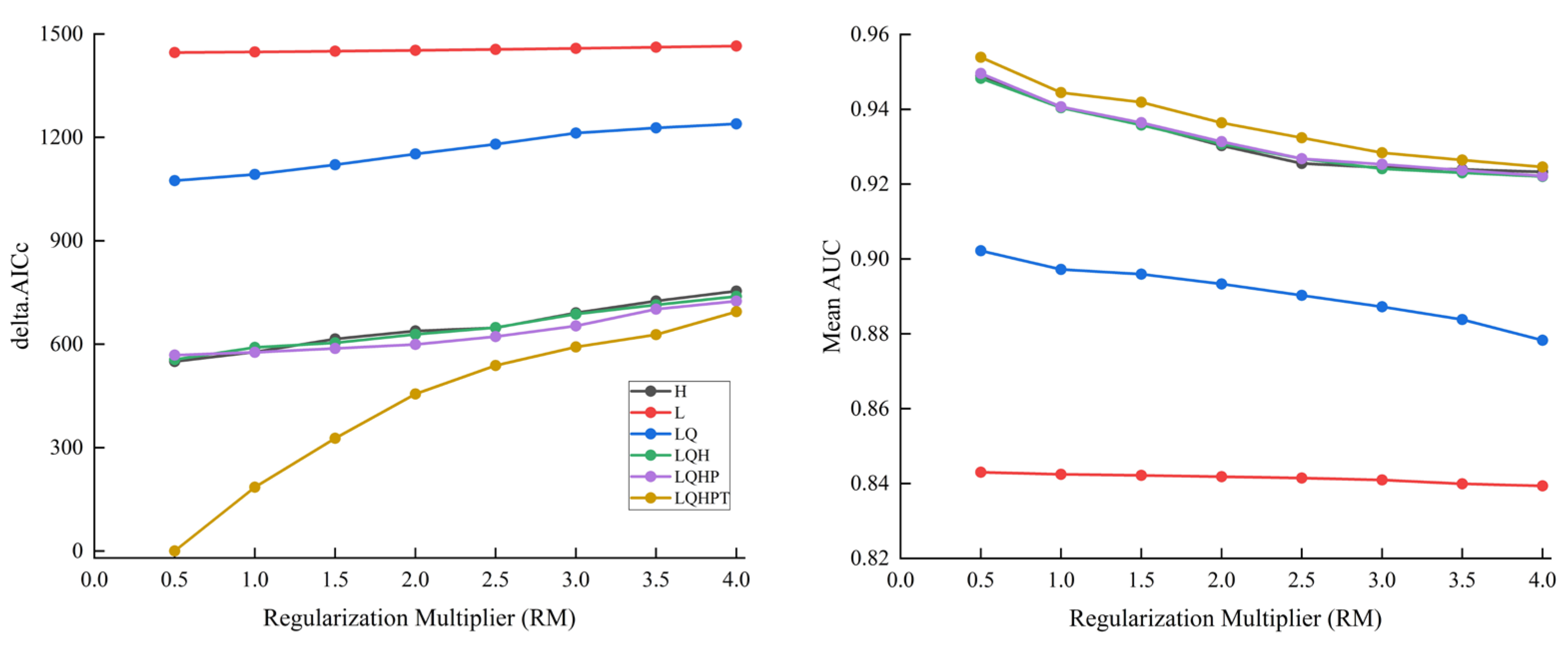
2.1. Optimization Model
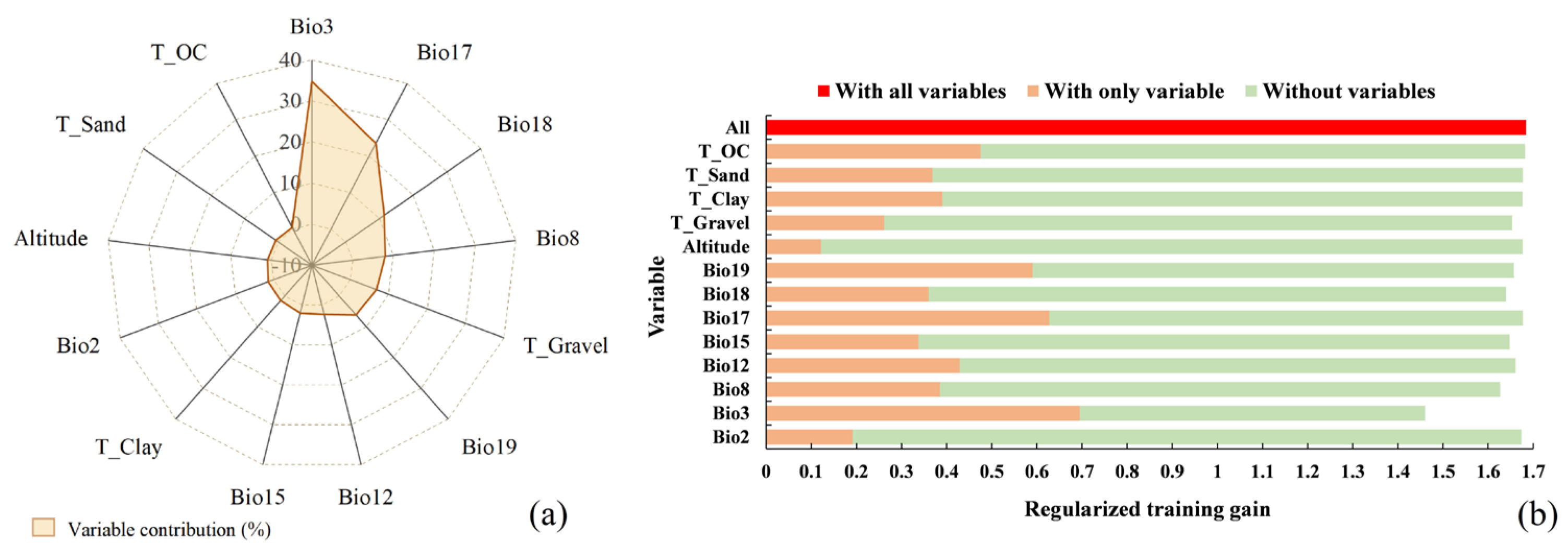
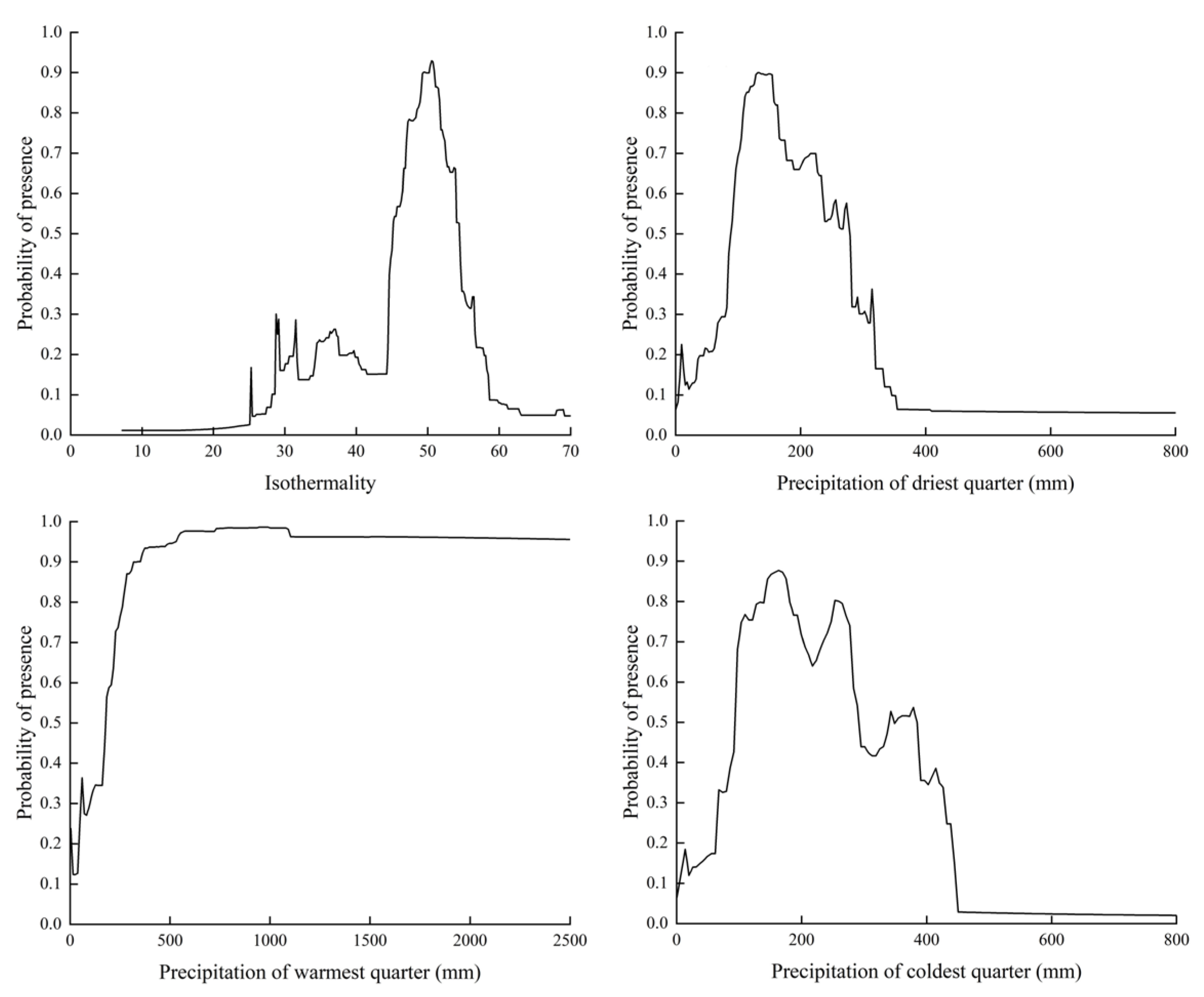
2.2. Significant Environmental Variables
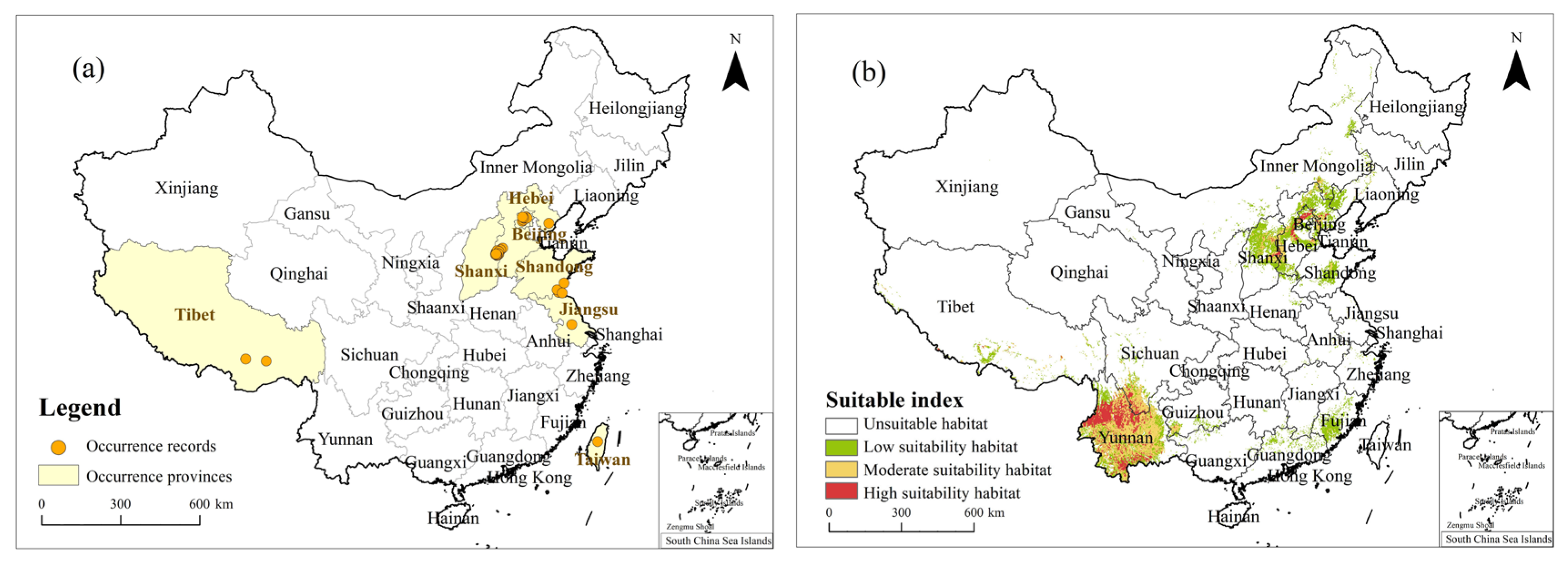
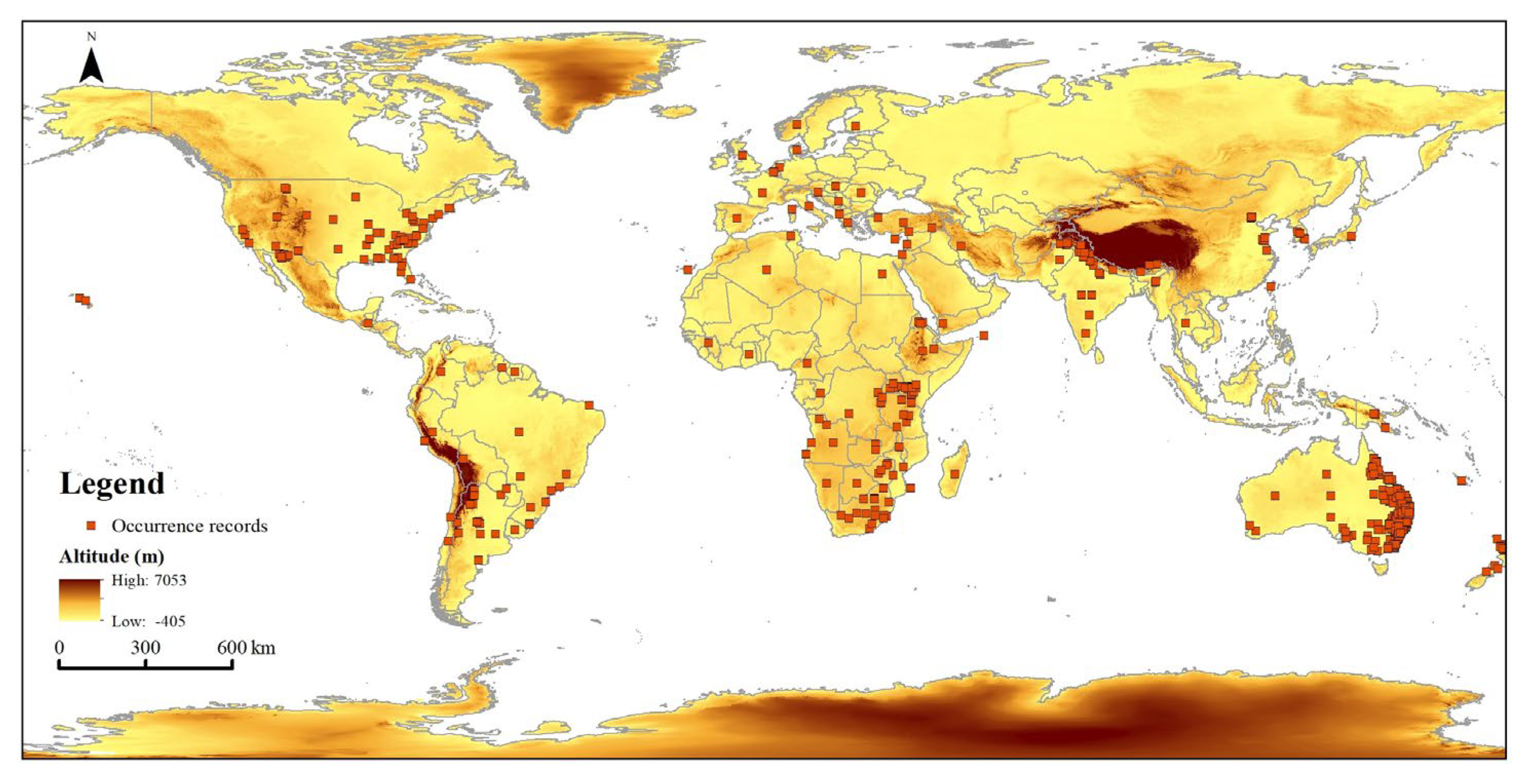
2.3. Occurrence Records and PGDs in China under the Current Climate
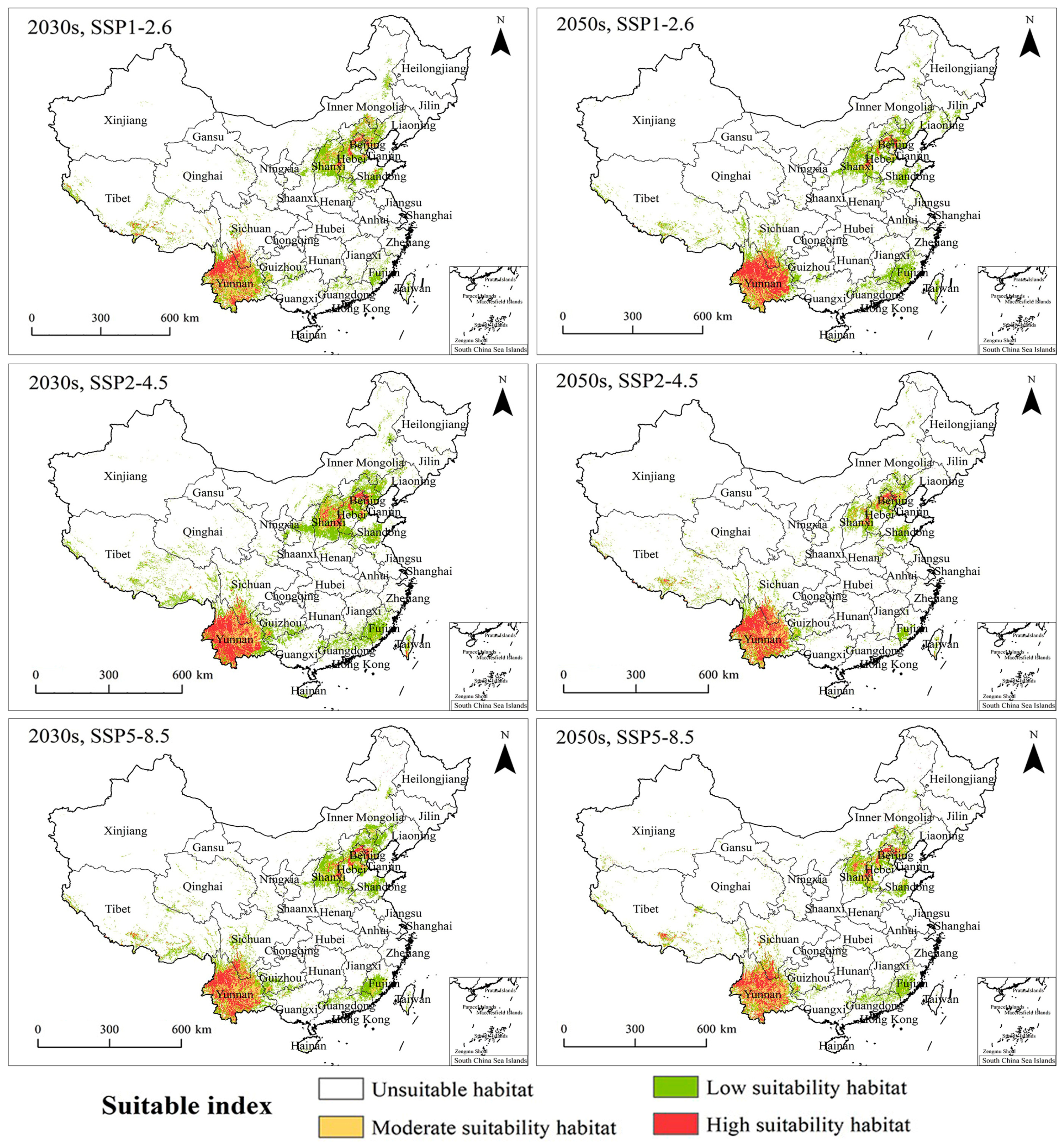
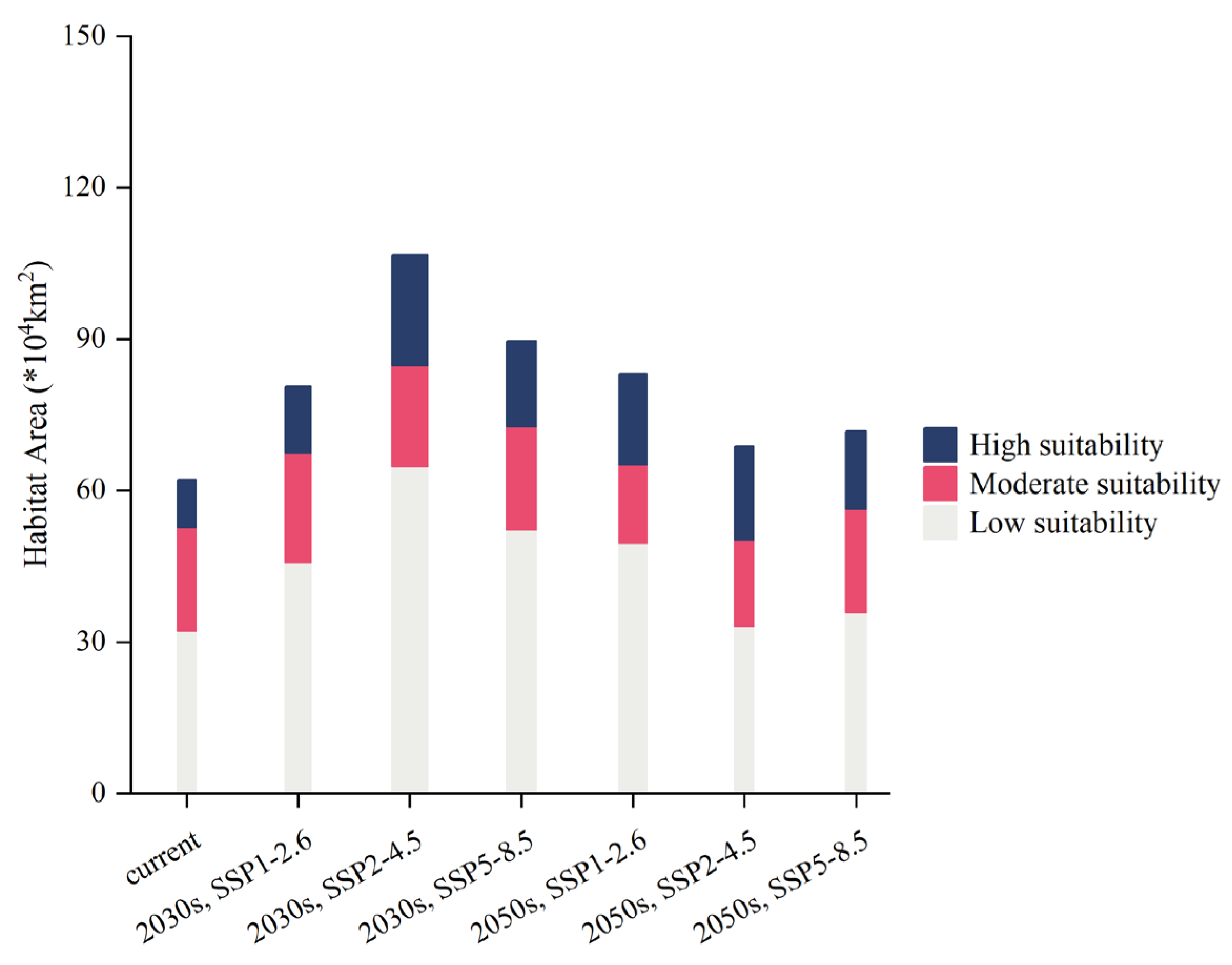
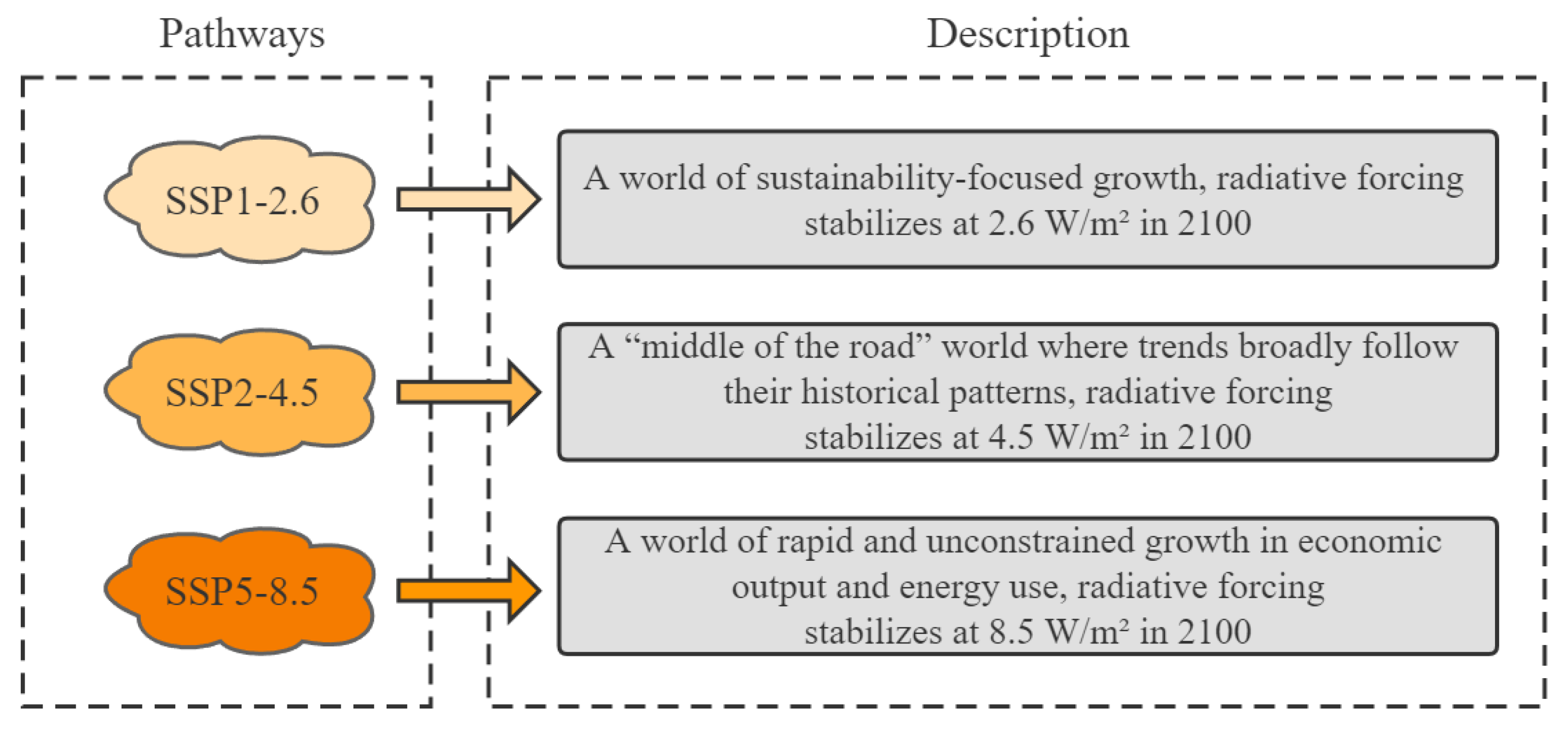
2.4. PGDs and Changes under Climate Change
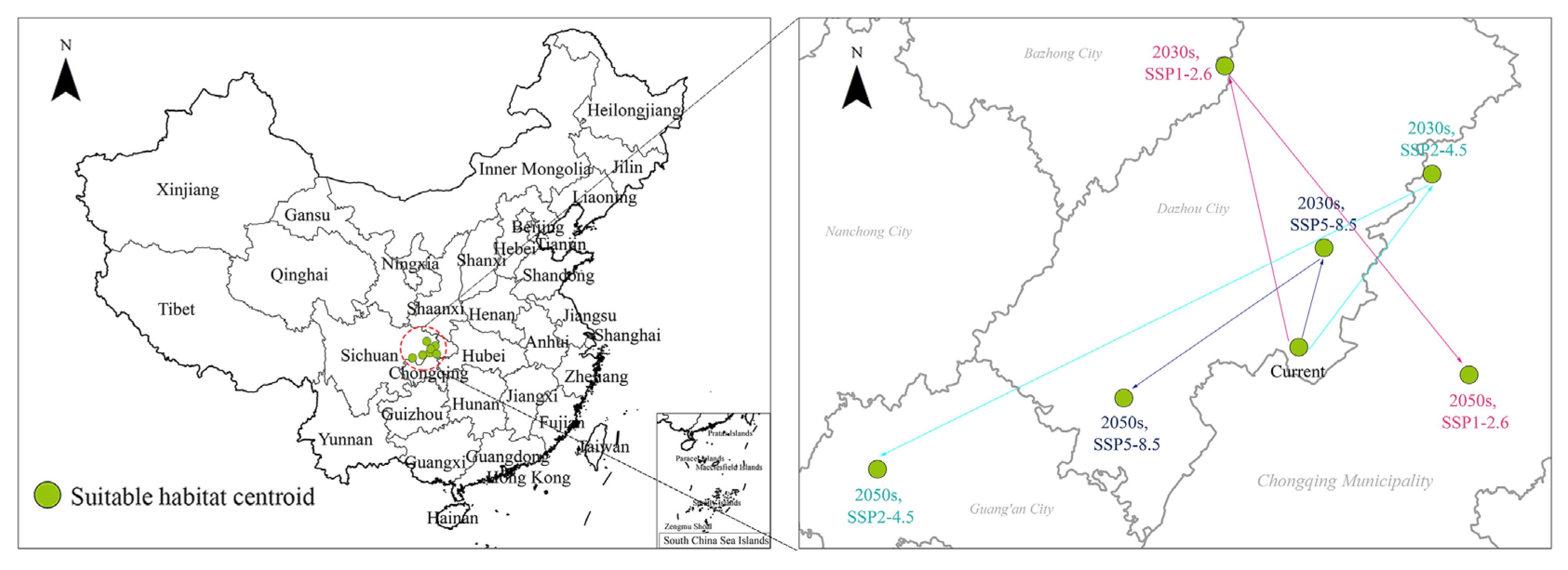
2.5. Centroid Distributional Shifts under Climate Change
3. Discussion
3.1. Environmental Variables Influenced PGDs of T. minuta
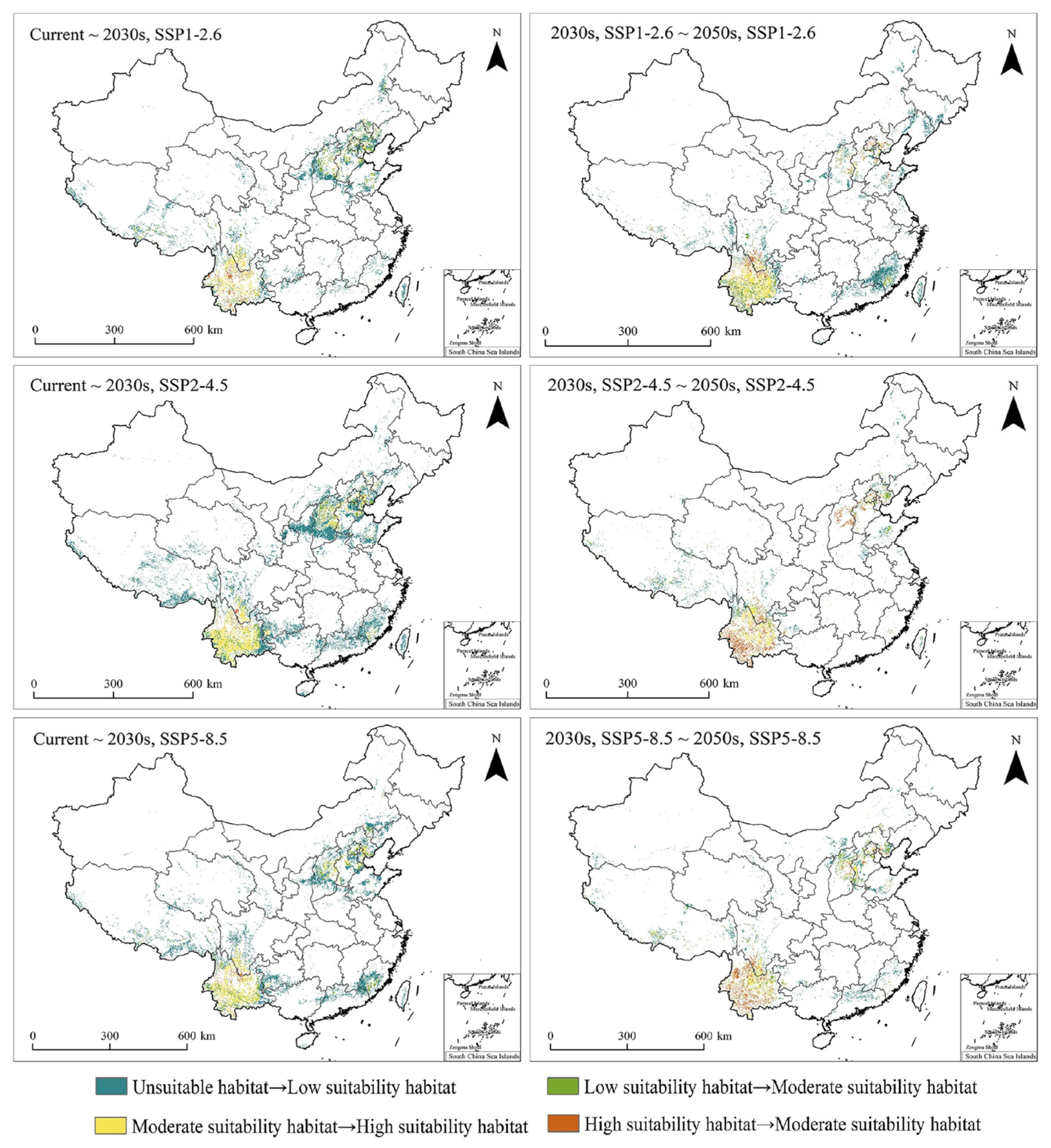
3.2. Changes in T. minuta PGDs
3.3. Strategies for Early Warning of T. minuta Invasion
4. Materials and Methods
4.1. Occurrence Records of T. minuta
4.2. Environmental Variables
4.3. Model Settings and Evaluation
4.4. Suitable Habitat Classification
4.5. Centroid of PGDs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wynne, J.J.; Howarth, F.G.; Mammola, S.; Ferreira, R.L.; Cardoso, P.; Lorenzo, T.D.; Galassi, D.M.P.; Medellin, R.A.; Miller, B.W.; Sánchez-Fernández, D.; et al. A conservation roadmap for the subterranean biome. Conserv. Lett. 2021, 14, e12834. [Google Scholar] [CrossRef]
- Tayeh, A.; Hufbauer, R.A.; Estoup, A.; Ravigné, V.; Frachon, L.; Facon, B. Biological invasion and biological control select for different life histories. Nat. Commun. 2015, 6, 7268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochocki, B.M.; Miller, T.E.X. Rapid evolution of dispersal ability makes biological invasions faster and more variable. Nat. Commun. 2017, 8, 14315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diagne, C.; Leroy, B.; Vaissière, A.-C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Sardain, A.; Sardain, E.; Leung, B. Global forecasts of shipping traffic and biological invasions to 2050. Nat. Sustain. 2019, 2, 274–282. [Google Scholar] [CrossRef]
- Zhang, L.; Rohr, J.; Cui, R.; Xin, Y.; Han, L.; Yang, X.; Gu, S.; Du, Y.; Liang, J.; Wang, X.; et al. Biological invasions facilitate zoonotic disease emergences. Nat. Commun. 2022, 13, 1762. [Google Scholar] [CrossRef]
- Rai, P.K.; Singh, J.S. Invasive alien plant species: Their impact on environment, ecosystem services and human health. Ecol. Indic. 2020, 111, 106020. [Google Scholar] [CrossRef]
- Omer, A.; Fristoe, T.; Yang, Q.; Razanajatovo, M.; Weigelt, P.; Kreft, H.; Dawson, W.; Dullinger, S.; Essl, F.; Pergl, J.; et al. The role of phylogenetic relatedness on alien plant success depends on the stage of invasion. Nat. Plants 2022, 8, 906–914. [Google Scholar] [CrossRef]
- Pagad, S.; Bisset, S.; Genovesi, P.; Groom, Q.; Hirsch, T.; Jetz, W.; Ranipeta, A.; Schigel, D.; Sica, Y.V.; McGeoch, M.A. Country compendium of the global register of introduced and invasive species. Sci. Data 2022, 9, 391. [Google Scholar] [CrossRef]
- Li, Y.; Shen, Z. Roles of dispersal limit and environmental filtering in shaping the spatiotemporal patterns of invasive alien plant diversity in China. Front. Ecol. Evol. 2020, 8, 544670. [Google Scholar] [CrossRef]
- Hannula, S.E.; Heinen, R.; Huberty, M.; Steinauer, K.; De Long, J.R.; Jongen, R.; Bezemer, T.M. Persistence of plant-mediated microbial soil legacy effects in soil and inside roots. Nat. Commun. 2021, 12, 5686. [Google Scholar] [CrossRef]
- Price, J.H.; Raduski, A.R.; Brandvain, Y.; Van Tassel, D.L.; Smith, K.P. Development of first linkage map for Silphium integrifolium (Asteraceae) enables identification of sporophytic self-incompatibility locus. Heredity 2022, 128, 304–312. [Google Scholar] [CrossRef]
- Baral, S.; Adhikari, A.; Khanal, R.; Malla, Y.; Kunwar, R.; Basnyat, B.; Gauli, K.; Acharya, R.P. Invasion of alien plant species and their impact on different ecosystems of Panchase Area, Nepal. Banko Janakari 2017, 27, 31–42. [Google Scholar] [CrossRef]
- Poudel, A.S.; Jha, P.K.; Shrestha, B.B.; Muniappan, R. Biology and management of the invasive weed Ageratina adenophora (Asteraceae): Current state of knowledge and future research needs. Weed Res. 2019, 59, 79–92. [Google Scholar] [CrossRef]
- Jin, H.; Chang, L.; van Kleunen, M.; Liu, Y. Soil mesofauna may buffer the negative effects of drought on alien plant invasion. J. Ecol. 2022, 110, 2332–2342. [Google Scholar] [CrossRef]
- Qian, H.; Rejmánek, M.; Qian, S. Are Invasive Species a Phylogenetically Clustered Subset of Naturalized Species in Regional Floras? A Case Study for Flowering Plants in China. Divers. Distrib. 2022, 28, 2084–2093. [Google Scholar] [CrossRef]
- Niu, H.; Liu, W.; Wan, F.H.; Liu, B. An invasive aster (Ageratina adenophora) invades and dominates forest understories in China: Altered soil microbial communities facilitate the invader and inhibit natives. Plant Soil 2007, 294, 73–85. [Google Scholar] [CrossRef]
- Babaei, K.; Moghaddam, M.; Farhadi, N.; Pirbalouti, A.G. Morphological, Physiological and Phytochemical Responses of Mexican Marigold (Tagetes minuta L.) to Drought Stress. Sci. Hort. 2021, 284, 110116. [Google Scholar] [CrossRef]
- Kumar, A.; Gautam, R.D.; Kumar, A.; Singh, S.; Singh, S. Understanding the effect of different abiotic stresses on wild marigold (Tagetes minuta L.) and role of breeding strategies for developing tolerant lines, 12. Front. Plant Sci. 2021, 12, 754457. [Google Scholar] [CrossRef]
- Kumar, A.; Gautam, R.D.; Singh, S.; Chauhan, R.; Kumar, A.; Singh, S. Comparative study of the effects of different soluble salts on seed germination of wild marigold (Tagetes minuta L.). J. Appl. Res. Med. Aromat. Plants 2022, 32, 100421. [Google Scholar] [CrossRef]
- Zhang, J.; Lv, Y.; Bian, Y.; Liu, R.; Jiang, L. A new kind of invasive plant from mainland China. Plant Quar. 2014, 28, 65–67. (In Chinese) [Google Scholar]
- Xu, M.; Tsering, T. A newly naturalized plant in Qinghai-Tibetan Plateau. Guihaia 2015, 35, 554–555. (In Chinese) [Google Scholar]
- Yun, L.; Zhang, R.; Song, Z.; Fu, W.; Wang, R.; Wang, Z.; Zhang, G. The effect of Tagetes minuta L. on the diversity of soil bacterial community. Ecol. Environ. Sci. 2020, 29, 901–909. (In Chinese) [Google Scholar]
- Ibrahim, S.R.M.; Mohamed, G.A.A. Tagetones A and B, new cytotoxic monocyclic diterpenoids from flowers of Tagetes minuta. Chin. J. Nat. Med. 2017, 15, 546–549. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y.; Wang, L.; Wang, X.; Wang, L.; Duan, Y. Status of invasive plants on local pollination networks: A case study of Tagetes minuta in Tibet based on pollen grains from pollinators. Biodivers. Sci. 2019, 27, 306–313. [Google Scholar] [CrossRef]
- Xu, L.; Chen, J.; Qi, H.; Shi, Y. Phytochemicals and their biological activities of plants in Tagetes L. Chin. Herb. Med. 2012, 4, 103–117. [Google Scholar] [CrossRef]
- Walia, S.; Kumar, R. Nitrogen and sulfur fertilization modulates the yield, essential oil and quality traits of wild marigold (Tagetes minuta L.) in the western Himalaya. Front. Plant Sci. 2020, 11, 631154. [Google Scholar] [CrossRef]
- Luo, D.; Silva, D.P.; De Marco Júnior, P.; Pimenta, M.; Caldas, M.M. Model approaches to estimate spatial distribution of bee species richness and soybean production in the Brazilian cerrado during 2000 to 2015. Sci. Total Environ. 2020, 737, 139674. [Google Scholar] [CrossRef]
- Banerjee, A.K.; Feng, H.; Lin, Y.; Liang, X.; Wang, J.; Huang, Y. Setting the priorities straight—Species distribution models assist to prioritize conservation targets for the mangroves. Sci. Total Environ. 2022, 806, 150937. [Google Scholar] [CrossRef]
- Mukul, S.A.; Alamgir, M.; Sohel, M.S.I.; Pert, P.L.; Herbohn, J.; Turton, S.M.; Khan, M.S.I.; Munim, S.A.; Reza, A.H.M.A.; Laurance, W.F. Combined effects of climate change and sea-level rise project dramatic habitat loss of the globally endangered Bengal tiger in the Bangladesh Sundarbans. Sci. Total Environ. 2019, 663, 830–840. [Google Scholar] [CrossRef]
- Wang, G.; Wang, C.; Guo, Z.; Dai, L.; Wu, Y.; Liu, H.; Li, Y.; Li, Y.; Chen, H.; Zhang, Y.; et al. Integrating Maxent model and landscape ecology theory for studying spatiotemporal dynamics of habitat: Suggestions for conservation of endangered Red-crowned crane. Ecol. Indic. 2020, 116, 106472. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Borgelt, J.; Sicacha-Parada, J.S.; Skarpaas, O.; Verones, F. Native range estimates for red-listed vascular plants. Sci. Data 2022, 9, 117. [Google Scholar] [CrossRef]
- Padalia, H.; Srivastava, V.; Kushwaha, S.P.S. Modeling potential invasion range of alien invasive species, Hyptis suaveolens (L.) Poit. in India: Comparison of MaxEnt and GARP. Ecol. Inform. 2014, 22, 36–43. [Google Scholar] [CrossRef]
- Liu, X.; Liu, H.; Gong, H.; Lin, Z.; Lv, S. Appling the one-class classification method of Maxent to detect an invasive plant Spartina alterniflora with Time-Series Analysis. Remote Sens. 2017, 9, 1120. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Deng, X.; Xiang, W.; Chen, L.; Ouyang, S. Predicting potential suitable habitats of Chinese fir under current and future climatic scenarios based on Maxent model. Ecol. Inform. 2021, 64, 101393. [Google Scholar] [CrossRef]
- Shabani, F.; Ahmadi, M.; Kumar, L.; Solhjouy-fard, S.; Shafapour Tehrany, M.; Shabani, F.; Kalantar, B.; Esmaeili, A. Invasive weed species’ threats to global biodiversity: Future scenarios of changes in the number of invasive species in a changing climate. Ecol. Indic. 2020, 116, 106436. [Google Scholar] [CrossRef]
- Kumar, R.; Ramesh, K.; Singh, R.D.; Prasad, R. Modulation of wild marigold (Tagetes minuta L.) phenophases towards the varying temperature regimes—A field study. J. Agrometeorol. 2010, 12, 234–240. [Google Scholar] [CrossRef]
- Taylor, N.J.; Hills, P.N.; Gold, J.D.; Stirk, W.A.; Staden, J.V. Factors contributing to the regulation of thermoinhibition in Tagetes minuta L. J. Plant Physiol. 2005, 162, 1270–1279. [Google Scholar] [CrossRef]
- Rathore, S.; Walia, S.; Kumar, R. Biomass and essential oil of Tagetes minuta influenced by pinching and harvesting stage under high precipitation conditions in the western Himalayas. J. Essent. Oil Res. 2018, 30, 360–368. [Google Scholar] [CrossRef]
- Mohamed, M.A.; Harris, P.J.; Henderson, J.; Senatore, F. Effect of drought stress on the yield and composition of volatile oils of drought-tolerant and non-drought-tolerant clones of Tagetes minuta. Planta Med. 2002, 68, 472–474. [Google Scholar] [CrossRef] [PubMed]
- Graven, E.H.; Webber, L.; Benians, G.; Venter, M.; Gardner, J.B. Effect of soil type and nutrient status on the yield and composition of Tagetes oil (Tagetes minuta L.). J. Essent. Oil Res. 1991, 3, 303–307. [Google Scholar] [CrossRef]
- Peel, M.C.; Finlayson, B.L.; McMahon, T.A. Updated world map of the Koppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef]
- Karimian, P.; Kavoosi, G.; Amirghofran, Z. Anti-oxidative and anti-inflammatory effects of Tagetes minuta essential oil in activated macrophages. Asian Pac. J. Trop. Biomed. 2014, 4, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaei, F.; Jamei, R.; Heidari, R. Evaluation of volatile profile, fatty acids composition and in vitro bioactivity of Tagetes minuta growing wild in Northern Iran. Adv. Pharm. Bull. 2018, 8, 115–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aljeddani, G.S.; Al-Harbi, N.A.; Al-Qahtani, S.M.; El-Absy, K.M.; Abdullatif, B.M.; Dahan, T.E. Inventory of some introduced and invasive plant species in some governorates of the Kingdom of Saudi Arabia. Appl. Ecol. Environ. Res. 2021, 19, 4373–4388. [Google Scholar] [CrossRef]
- Cao, Y.; Xiao, Y.; Zhang, S.; Hu, W. Simulated warming enhances biological invasion of Solidago canadensis and Bidens frondosa by increasing reproductive investment and altering flowering phenology pattern. Sci. Rep. 2018, 8, 16073. [Google Scholar] [CrossRef] [Green Version]
- Brändel, M. Dormancy and germination of heteromorphic achenes of Bidens frondosa. Flora 2004, 199, 228–233. [Google Scholar] [CrossRef]
- Tankeu, S.Y.; Vermaak, I.; Viljoen, A.M.; Sandasi, M.; Kamatou, G.P.P. Essential oil variation of Tagetes minuta in South Africa–A chemometric approach. Biochem. Syst. Ecol. 2013, 51, 320–327. [Google Scholar] [CrossRef]
- Stroze, C.T.; Baida, F.C.; Balbi-Peña, M.I.; Dias-Arieira, C.R.; Santiago, D.C. Tagetes minuta propagation and interaction with nematoide. J. Agric. Sci. 2019, 11, 139. [Google Scholar] [CrossRef]
- Sadia, S.; Khalid, S.; Qureshi, R.; Bajwa, A. Tagetes minuta L., a useful underutilized plant of family Asteraceae: A review. Pak. J. Weed Sci. Res. 2013, 19, 179–189. [Google Scholar]
- Green, M.M.; Singer, J.M.; Sutherland, D.J.; Hibben, C.R. Larvicidal activity of Tagetes minuta (marigold) toward Aedes aegypti. J. Am. Mosq. Control Assoc. 1991, 7, 282–286. [Google Scholar]
- Scrivanti, L.R.; Zunino, M.P.; Zygadlo, J.A. Tagetes minuta and Schinus areira essential oils as allelopathic agents. Biochem. Syst. Ecol. 2003, 31, 563–572. [Google Scholar] [CrossRef]
- Javidan, N.; Kavian, A.; Pourghasemi, H.R.; Conoscenti, C.; Jafarian, Z.; Rodrigo-Comino, J. Evaluation of multi-hazard map produced using MaxEnt machine learning technique. Sci. Rep. 2021, 11, 6496. [Google Scholar] [CrossRef]
- Betts, M.G.; Yang, Z.; Hadley, A.S.; Smith, A.C.; Rousseau, J.S.; Northrup, J.M.; Nocera, J.J.; Gorelick, N.; Gerber, B.D. Forest degradation drives widespread avian habitat and population declines. Nat. Ecol. Evol. 2022, 6, 709–719. [Google Scholar] [CrossRef]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Olvera, L.O. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [Green Version]
- Bates, O.K.; Ollier, S.; Bertelsmeier, C. Smaller climatic niche shifts in invasive than non-invasive alien ant species. Nat. Commun. 2020, 11, 5213. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, Y.; Xian, X.; Zhao, H.; Wang, R.; Huang, H.; Zhang, Y.; Yang, M.; Liu, W. Increased Invasion Risk of Tagetes minuta L. in China under Climate Change: A Study of the Potential Geographical Distributions. Plants 2022, 11, 3248. https://doi.org/10.3390/plants11233248
Qi Y, Xian X, Zhao H, Wang R, Huang H, Zhang Y, Yang M, Liu W. Increased Invasion Risk of Tagetes minuta L. in China under Climate Change: A Study of the Potential Geographical Distributions. Plants. 2022; 11(23):3248. https://doi.org/10.3390/plants11233248
Chicago/Turabian StyleQi, Yuhan, Xiaoqing Xian, Haoxiang Zhao, Rui Wang, Hongkun Huang, Yanping Zhang, Ming Yang, and Wanxue Liu. 2022. "Increased Invasion Risk of Tagetes minuta L. in China under Climate Change: A Study of the Potential Geographical Distributions" Plants 11, no. 23: 3248. https://doi.org/10.3390/plants11233248