
The Biostimulant Effect of Hydroalcoholic Extracts of Sargassum spp. in Tomato Seedlings under Salt Stress
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Seedling Growth and Biomass
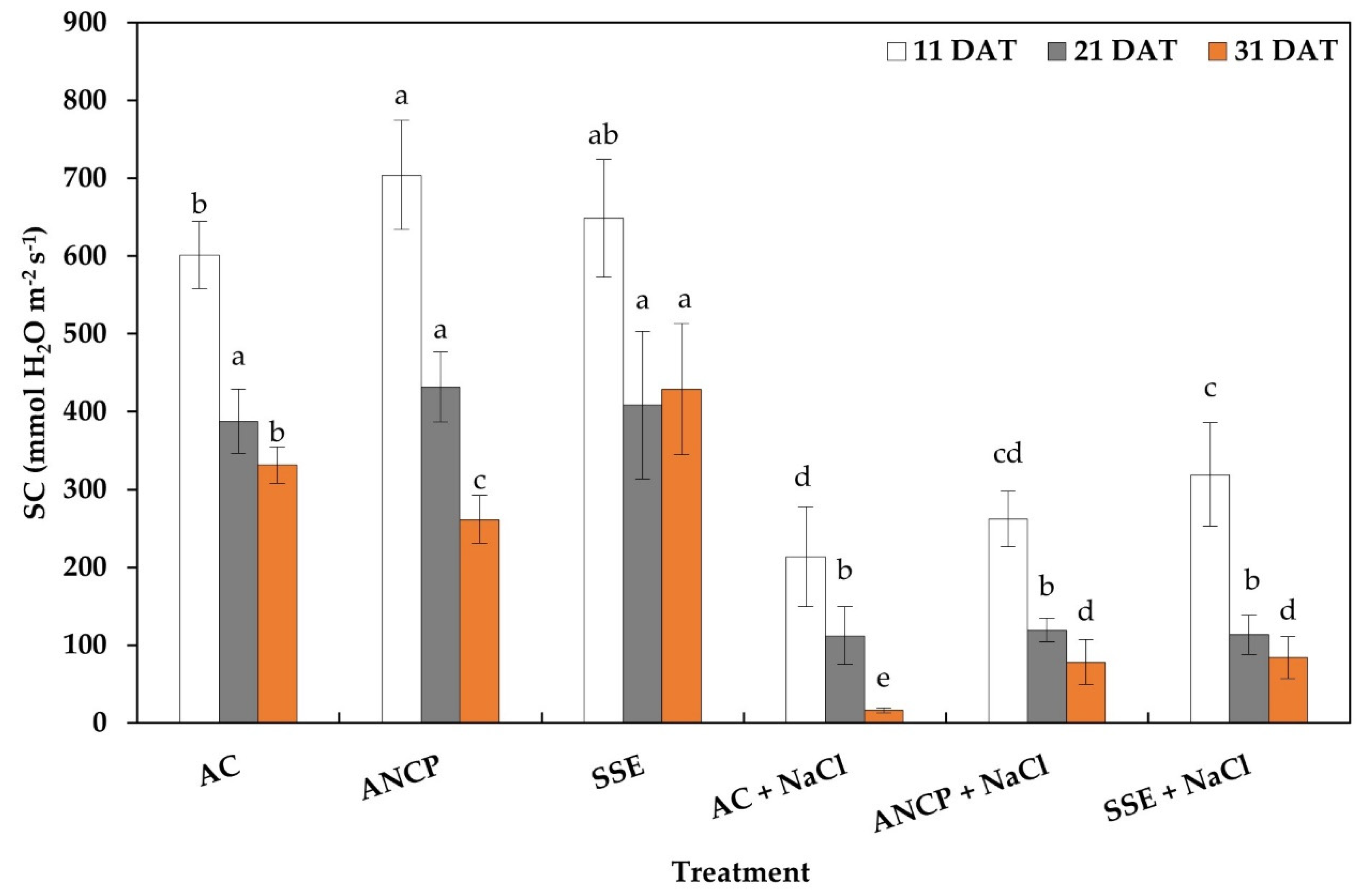
2.2. Stomatal Conductance of Leaves
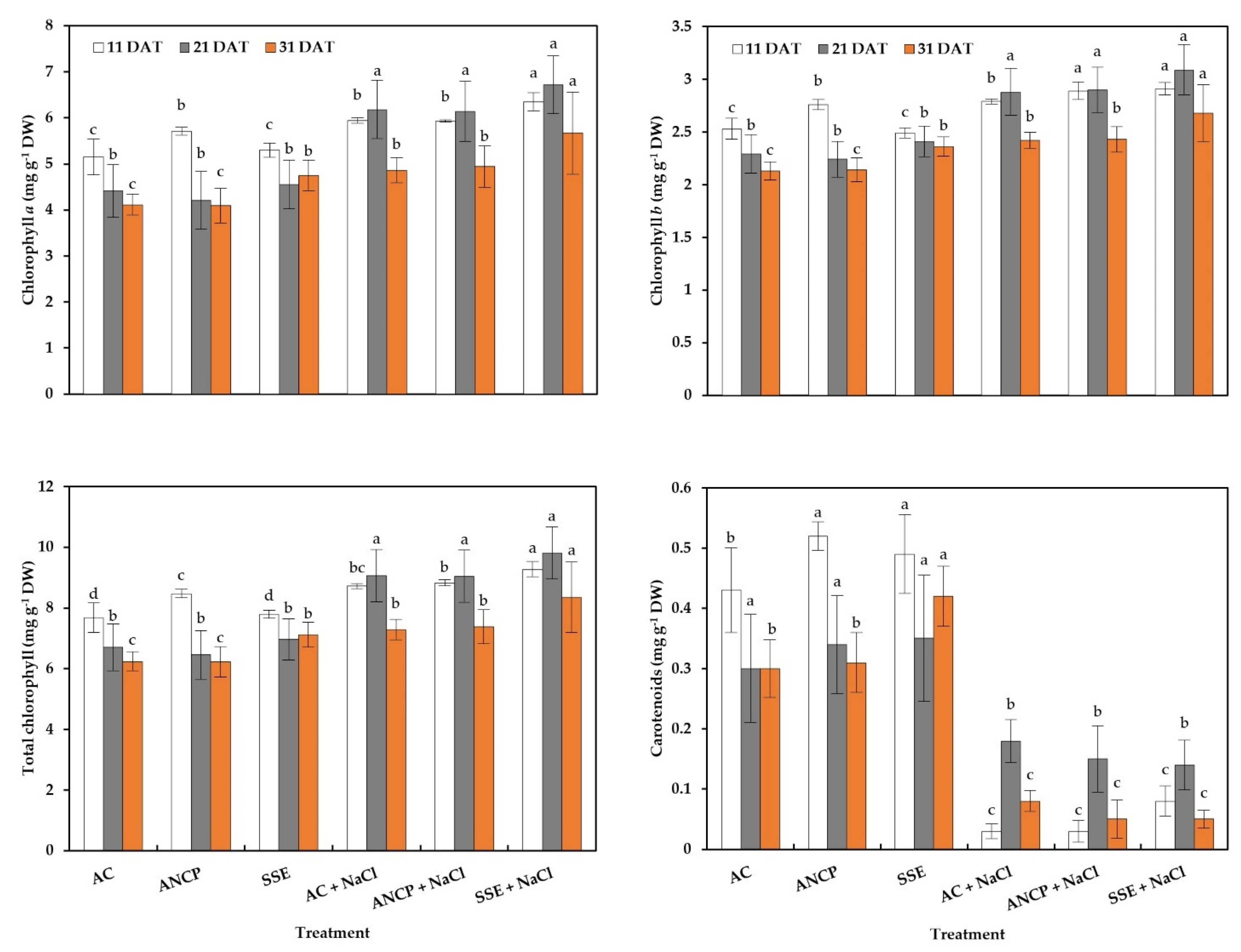
2.3. Photosynthetic Pigments
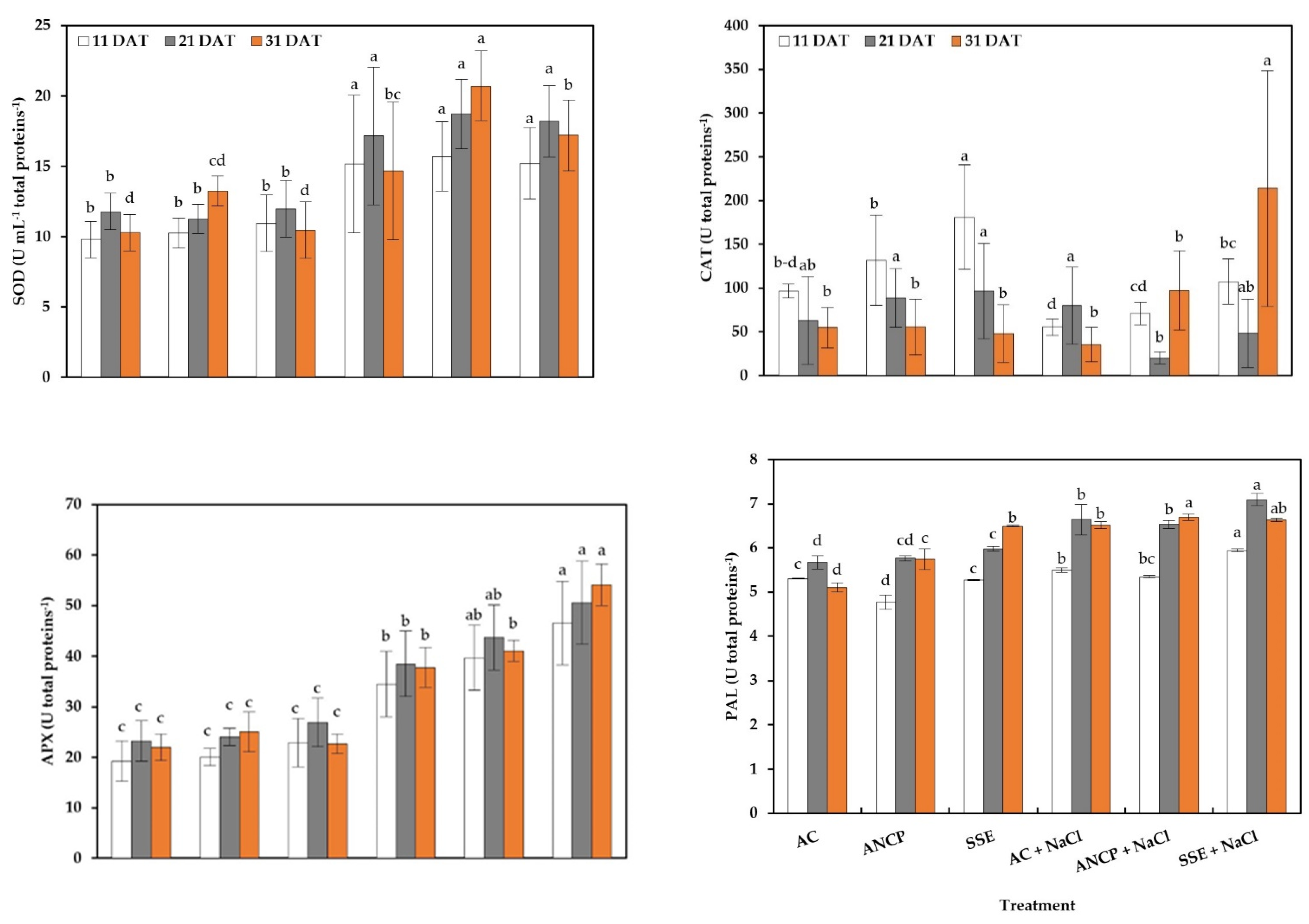
2.4. Enzymatic Activity and Total Proteins
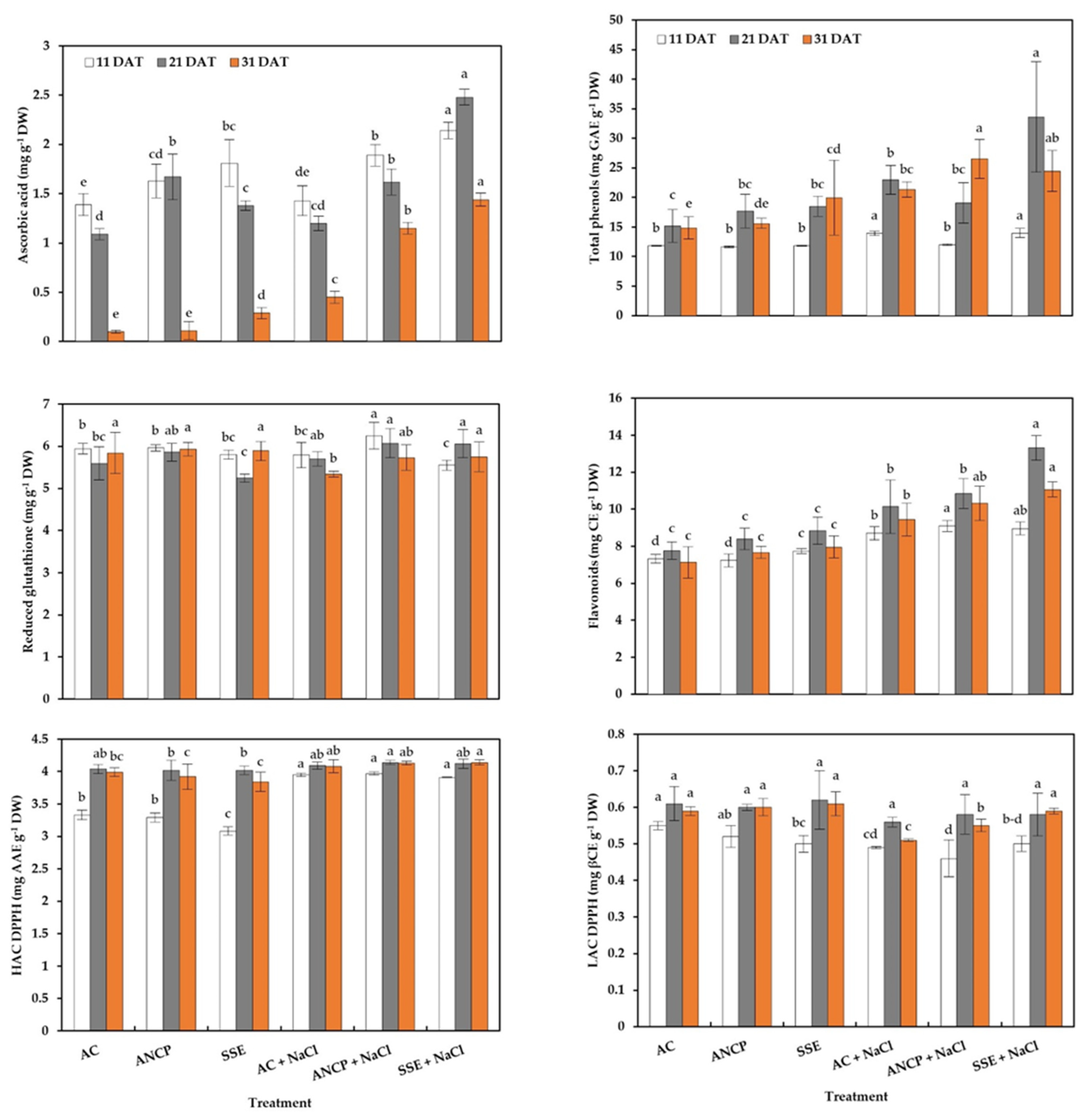
2.5. Hydrophilic Antioxidants and Antioxidant Capacity
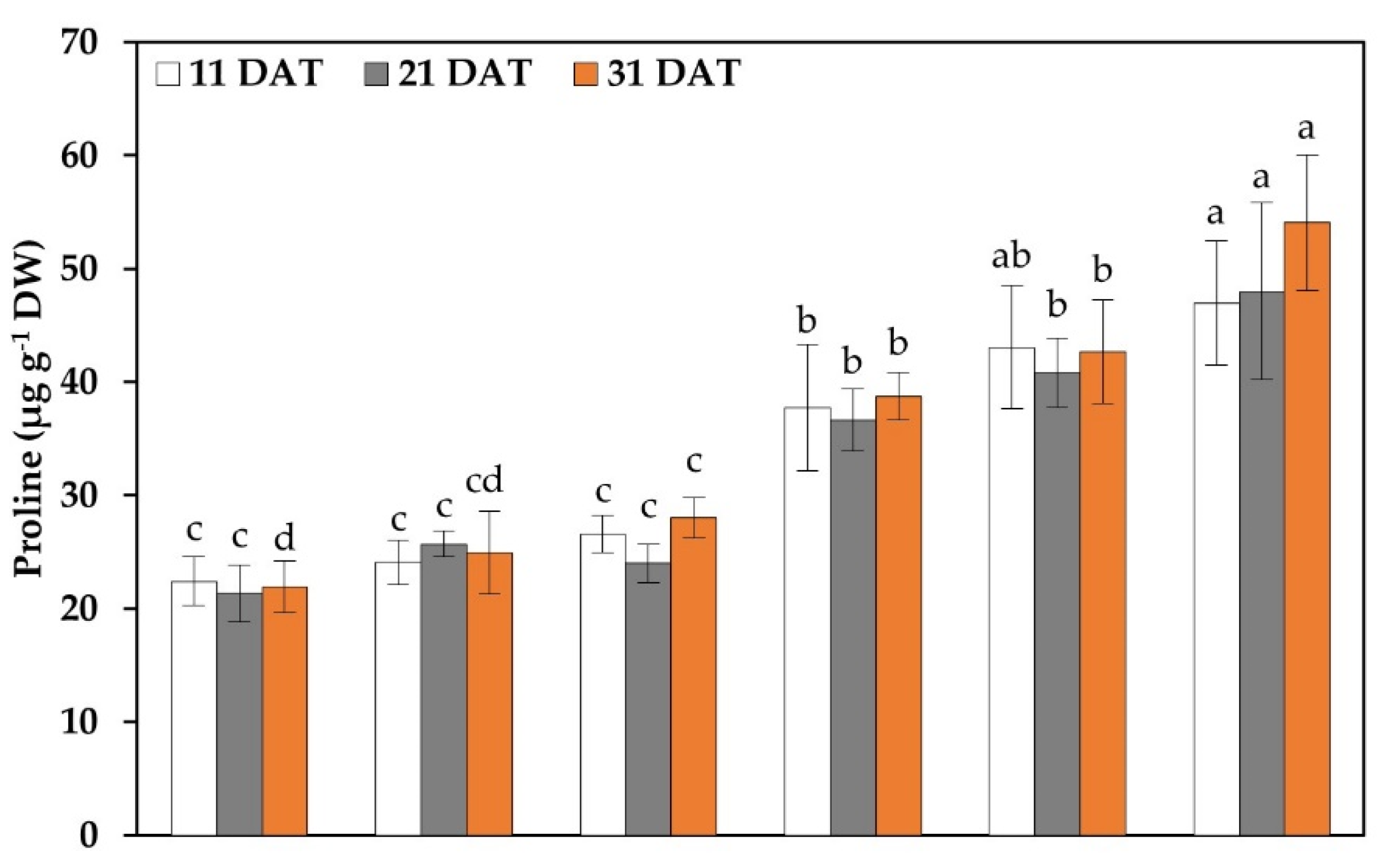
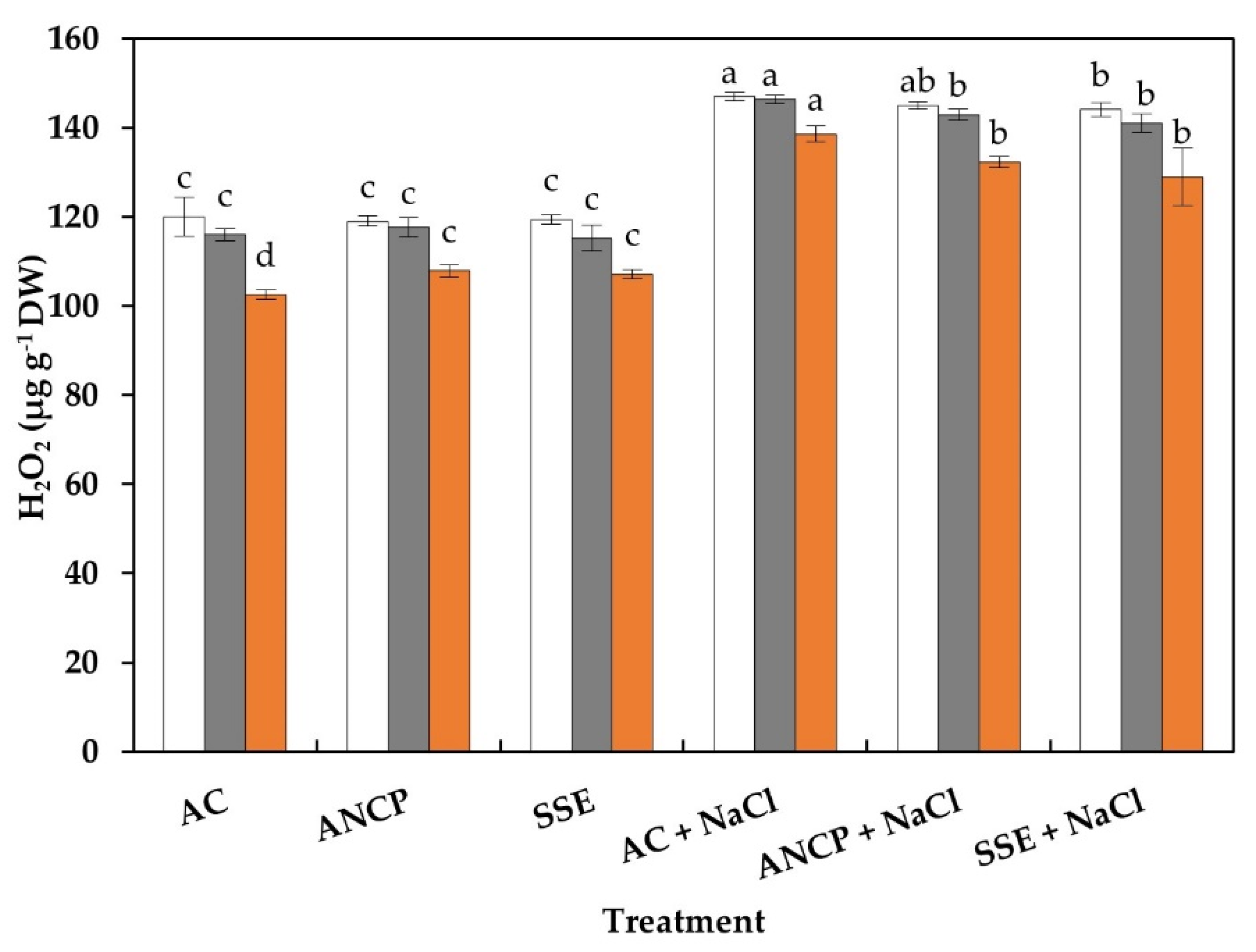
2.6. Proline and Hydrogen Peroxide
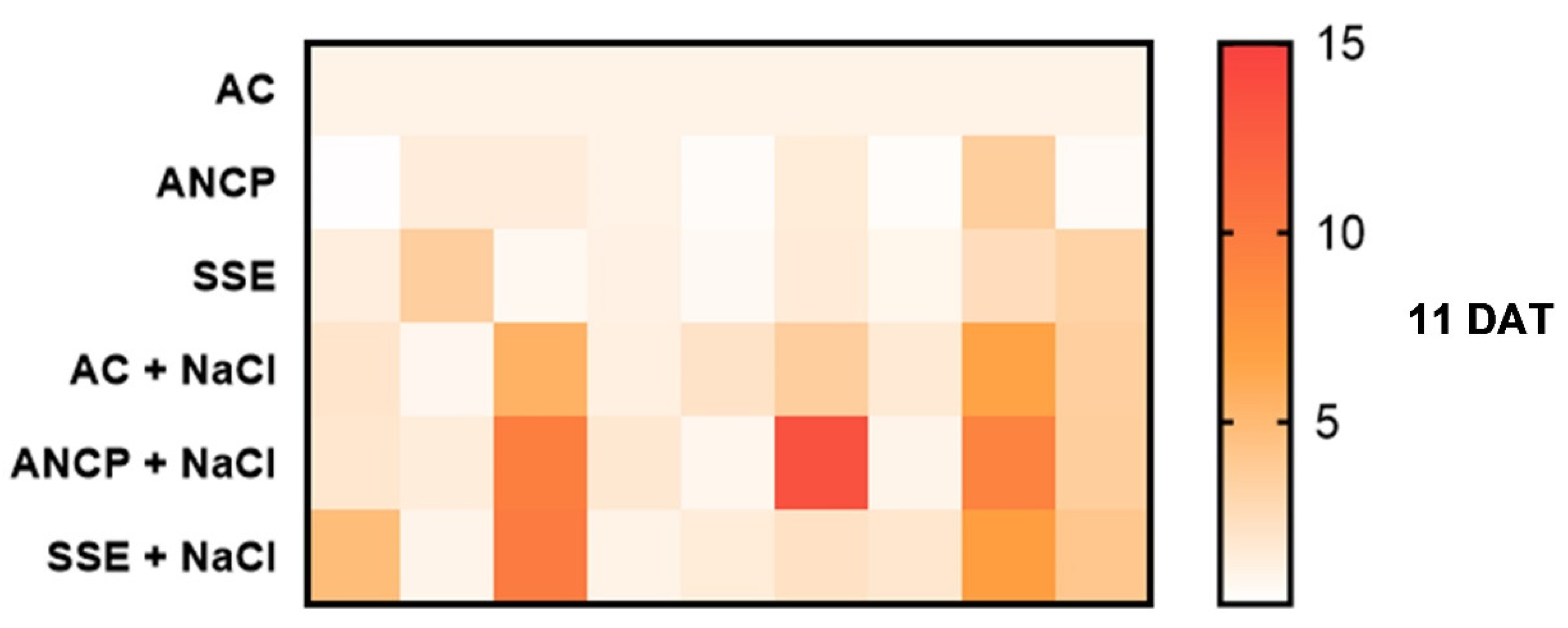
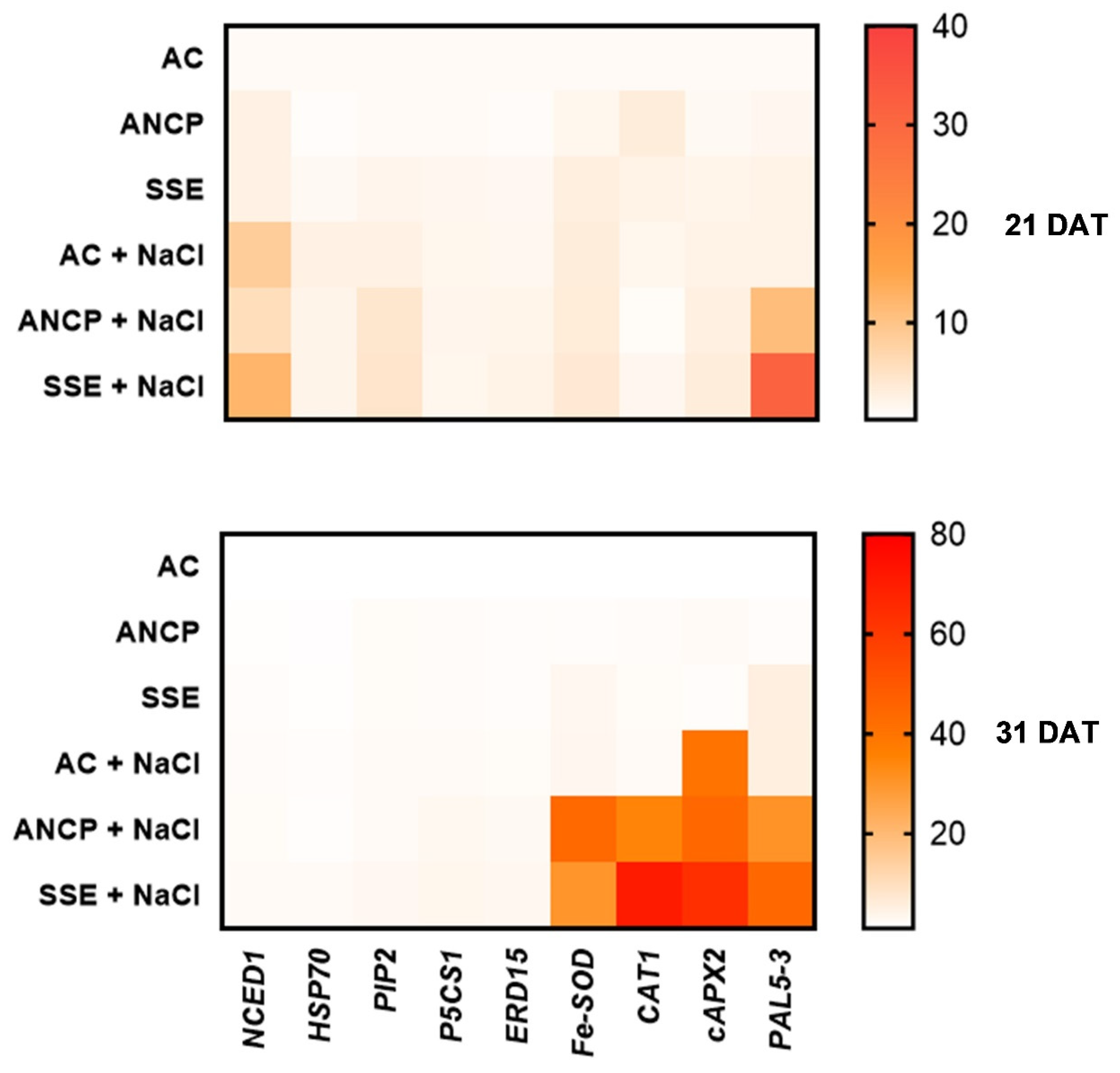
2.7. Expression of Defense Genes
3. Discussion
3.1. Seedling Biomass and Growth Parameters
3.2. Stomatal Conductance
3.3. Photosynthetic Pigments
3.4. Enzymatic Activity, Hydrophilic Antioxidants, and Antioxidant Capacity
3.5. Total Protein, Proline, and Hydrogen Peroxide
3.6. Expression of Defense Genes
4. Materials and Methods
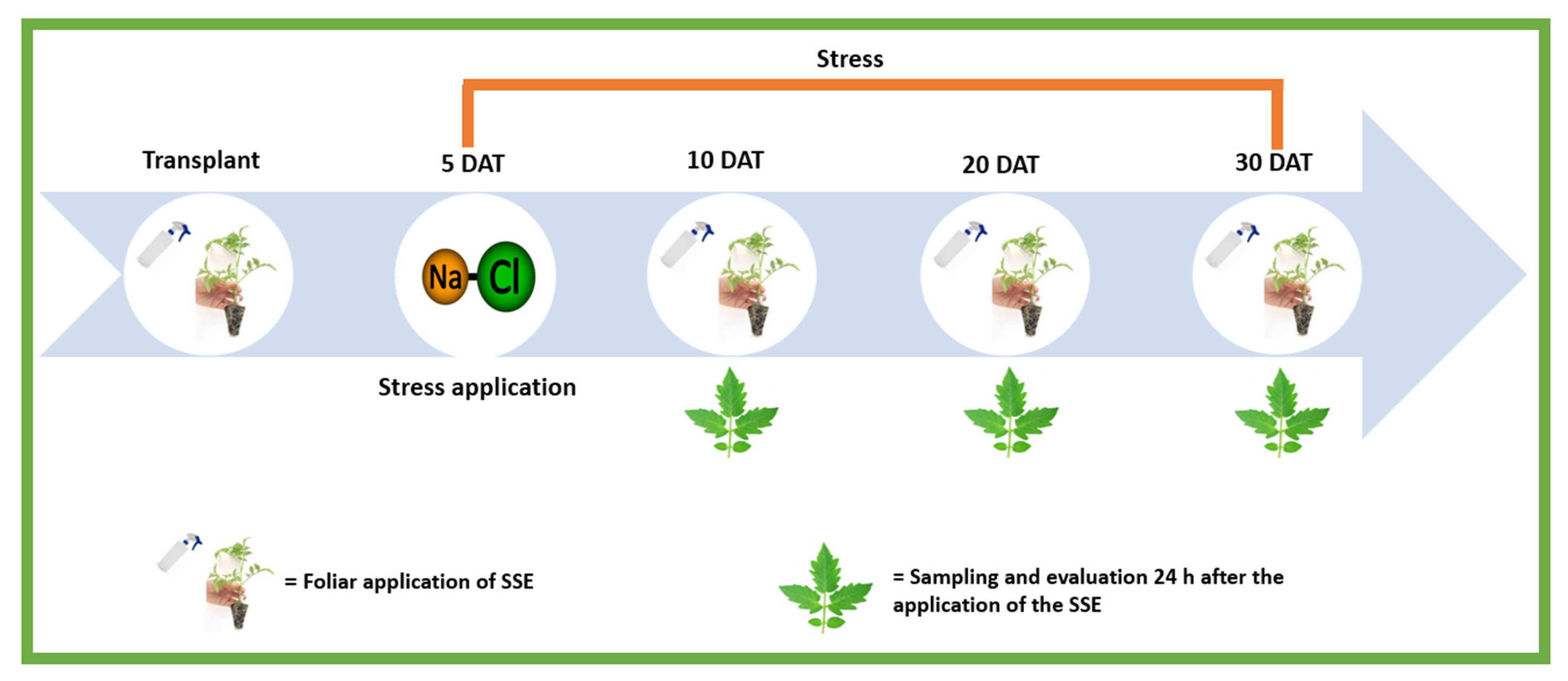
4.1. Plant Material and Experimental Conditions
4.2. Treatments
4.3. Sampling and Evaluations
4.4. Biochemical Analyses
4.5. Real-Time Reverse Transcription PCR
4.6. Experimental Design and Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jēkabsone, A.; Andersone-Ozola, U.; Karlsons, A.; Romanovs, M.; Ievinsh, G. Effect of Salinity on Growth, Ion Accumulation and Mineral Nutrition of Different Accessions of a Crop Wild Relative Legume Species, Trifolium fragiferum. Plants 2022, 11, 797. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Ahanger, M.A.; Alam, P.; Alyemeni, M.N.; Wijaya, L.; Ali, S.; Ashraf, M. Silicon (Si) Supplementation Alleviates NaCl Toxicity in Mung Bean [Vigna radiata (L.) Wilczek] Through the Modifications of Physio-biochemical Attributes and Key Antioxidant Enzymes. J. Plant Growth Regul. 2018, 38, 70–82. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach toward salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, H.; Juárez-Maldonado, A.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Sánchez-Aspeytia, D.; González-Morales, S. Chitosan-PVA and Copper Nanoparticles Improve Growth and Overexpress the SOD and JA Genes in Tomato Plants under Salt Stress. Agronomy 2018, 8, 175. [Google Scholar] [CrossRef] [Green Version]
- Raja, V.; Majeed, U.; Kang, H.; Andrabi, K.I.; John, R. Abiotic stress: Interplay between ROS, hormones and MAPKs. Environ. Exp. Bot. 2017, 137, 142–157. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Borhannuddin Bhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Hewedy, O.A.; Battaglia, M.L.; Jalal, R.S.; Alhammad, B.A.; Schillaci, C.; Ali, N.; Al-Doss, A. Field crop responses and management strategies to mitigate soil salinity in modern agriculture: A review. Agronomy 2021, 11, 2299. [Google Scholar] [CrossRef]
- Juárez-Maldonado, A.; Tortella, G.; Rubilar, O.; Fincheira, P.; Benavides-Mendoza, A. Biostimulation and toxicity: The magnitude of the impact of nanomaterials in microorganisms and plants. J. Adv. Res. 2021, 31, 113–126. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Prespective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [Green Version]
- du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- European Union (EU). Regulation of the European Parliament and of the Council Laying down Rules on the Making Available on the Market of EU Fertilizing Products ond Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003. 2019. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=OJ:L:2019:170:TOC (accessed on 3 November 2022).
- Mukherjee, A.; Patel, J.S. Seaweed extract: Biostimulator of plant defense and plant productivity. Int. J. Environ. Sci. Technol. 2020, 17, 553–558. [Google Scholar] [CrossRef]
- Benítez-García, I.; Dueñas-Ledezma, A.K.; Martínez-Montaño, E.; Salazar-Leyva, J.A.; Carrera, E.; Osuna-Ruiz, I. Identification and Quantification of Plant Growth Regulators and Antioxidant Compounds in Aqueous Extracts of Padina durvillaei and Ulva lactuca. Agronomy 2020, 10, 866. [Google Scholar] [CrossRef]
- Sariñana-Aldaco, O.; Benavides-Mendoza, A.; Juárez-Maldonado, A.; Robledo-Olivo, A.; Rodríguez-Jasso, R.M.; Preciado-Rangel, P.; Gonzalez-Morales, S. Efecto de extractos de Sargassum spp. en el crecimiento y antioxidantes de plántulas de tomate. Ecosistemas Y Recur. Agropecu. 2021, 8, e2814. [Google Scholar] [CrossRef]
- Hamed, S.M.; Abd El-Rhman, A.A.; Abdel-Raouf, N.; Ibraheem, I.B.M. Role of marine macroalgae in plant protection & improvement for sustainable agriculture technology. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 104–110. [Google Scholar] [CrossRef]
- Zou, P.; Lu, X.; Zhao, H.; Yuan, Y.; Meng, L.; Zhang, C.; Li, Y. Polysaccharides derived from the brown algae Lessonia nigrescens enhance salt stress tolerance to wheat seedlings by enhancing the antioxidant system and modulating intracellular ion concentration. Front. Plant Sci. 2019, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Diario Oficial de la Federación (DOF). NORMA Oficial Mexicana NOM-182-SSA1-2010, Etiquetado de Nutrientes Vegetales. Available online: https://www.dof.gob.mx/normasOficiales/4371/salud1a1.htm#:~:text=1.1%20Esta%20norma%20establece%20las,regladores%20de%20crecimiento%20tipo%203 (accessed on 6 October 2022).
- González-Morales, S.; Solís-Gaona, S.; Valdés-Caballero, M.V.; Juárez-Maldonado, A.; Loredo-Treviño, A.; Benavides-Mendoza, A. Transcriptomics of Biostimulation of Plants Under Abiotic Stress. Front. Genet. 2021, 12, 583888. [Google Scholar] [CrossRef]
- Chávez, V.; Uribe-Martínez, A.; Cuevas, E.; Rodríguez-Martínez, R.E.; van Tussenbroek, B.I.; Francisco, V.; Estévez, M.; Celis, L.B.; Monroy-Velázquez, V.; Leal-Bautista, R.; et al. Massive Influx of Pelagic Sargassum spp. on the Coasts of the Mexican Caribbean 2014-2020: Challenges and Opportunities. Water 2020, 12, 2908. [Google Scholar] [CrossRef]
- López-Sosa, L.B.; Morales-Máximo, M.; Anastacio-Paulino, R.; Custodio-Hernández, A.; Corral-Huacuz, J.C.; Aguilera-Mandujano, A. Electron Microscopy Characterization of Sargassum Spp. from the Mexican Caribbean for Application as a Bioconstruction Material. Microsc. Microanal. 2021, 27, 3140–3143. [Google Scholar] [CrossRef]
- Waghmode, A.V.; Narayankar, C.U.; Nimbalkar, M.S.; Gaikwad, D.K. Exploration of fucoxanthin and astaxanthin in macro alga (Sargassum sp.) by high-performance liquid chromatography. Indian Hydrobiol. 2019, 18, 40–49. [Google Scholar]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Phytoelicitor activity of Sargassum vulgare and Acanthophora spicifera extracts and their prospects for use in vegetable crops for sustainable crop production. J. Appl. Phycol. 2021, 33, 639–651. [Google Scholar] [CrossRef]
- Abdel Latef, A.A.H.; Srivastava, A.K.; Saber, H.; Alwaleed, E.A.; Tran, L.S.P. Sargassum muticum and Jania rubens regulate amino acid metabolism to improve growth and alleviate salinity in chickpea. Sci. Rep. 2017, 7, 10537. [Google Scholar] [CrossRef] [PubMed]
- Sofy, M.R.; Sharaf, A.E.M.A.; Osman, M.S.; Sofy, A.R. Physiological changes, antioxidant activity, lipid peroxidation and yield characters of salt stressed barely plant in response to treatment with Sargassum extract. Int. J. Adv. Res. Biol. Sci. 2017, 4, 90–109. [Google Scholar] [CrossRef]
- Jithesh, M.N.; Shukla, P.S.; Kant, P.; Joshi, J.; Critchley, A.T.; Prithiviraj, B. Physiological and Transcriptomics Analyses Reveal that Ascophyllum nodosum Extracts Induce Salinity Tolerance in Arabidopsis by Regulating the Expression of Stress Responsive Genes. J. Plant Growth Regul. 2019, 38, 463–478. [Google Scholar] [CrossRef]
- Shin, Y.K.; Bhandari, S.R.; Jo, J.S.; Song, J.W.; Cho, M.C.; Yang, E.Y.; Lee, J.G. Response to salt stress in lettuce: Changes in chlorophyll fluorescence parameters, phytochemical contents, and antioxidant activities. Agronomy 2020, 10, 1627. [Google Scholar] [CrossRef]
- Medrano Macías, J.; López Caltzontzit, M.G.; Rivas Martínez, E.N.; Narváez Ortiz, W.A.; Benavides Mendoza, A.; Martínez Lagunes, P. Enhancement to salt stress tolerance in strawberry plants by iodine products application. Agronomy 2021, 11, 602. [Google Scholar] [CrossRef]
- Zhu, D.; Luo, F.; Zou, R.; Liu, J.; Yan, Y. Integrated physiological and chloroplast proteome analysis of wheat seedling leaves under salt and osmotic stresses. J. Proteomics 2021, 234, 104097. [Google Scholar] [CrossRef]
- Darko, E.; Végh, B.; Khalil, R.; Marček, T.; Szalai, G.; Pál, M.; Janda, T. Metabolic responses of wheat seedlings to osmotic stress induced by various osmolytes under iso-osmotic conditions. PLoS ONE 2019, 14, e0226151. [Google Scholar] [CrossRef]
- Turk, H. Chitosan-induced enhanced expression and activation of alternative oxidase confer tolerance to salt stress in maize seedlings. Plant Physiol. Biochem. 2019, 141, 415–422. [Google Scholar] [CrossRef]
- Che-Othman, M.H.; Jacoby, R.P.; Millar, A.H.; Taylor, N.L. Wheat mitochondrial respiration shifts from the tricarboxylic acid cycle to the GABA shunt under salt stress. New Phytol. 2020, 225, 1166–1180. [Google Scholar] [CrossRef]
- Gharib, F.A.E.L.; Zeid, I.M.; Salem, O.M.A.-H.; Ahmed, E.Z. Effects of Sargassum latifolium Extract on Growth, Oil Content and Enzymatic Activities of Rosemary Plants under Salinity Stress. Life Sci. J. 2014, 11, 933–945. [Google Scholar]
- Fleurence, J. Perspectives on the Use of Algae in Agriculture and Animal Production. Phycology 2021, 1, 79–82. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Chen, C.-L.; Song, W.-L.; Sun, L.; Qin, S.; Ren, C.-G.; Yang, J.-C.; Feng, D.-W.; Liu, N.; Yan, J.; Cui, B.-B.; et al. Effect of Seaweed Extract Supplement on Rice Rhizosphere Bacterial Community in Tillering and Heading Stages. Agronomy 2022, 12, 342. [Google Scholar] [CrossRef]
- Chandra, S.; Askari, K.; Kumari, M. Optimization of indole acetic acid production by isolated bacteria from Stevia rebaudiana rhizosphere and its effects on plant growth. J. Genet. Eng. Biotechnol. 2018, 16, 581–586. [Google Scholar] [CrossRef]
- Hönig, M.; Plíhalová, L.; Husičková, A.; Nisler, J.; Doležal, K. Role of cytokinins in senescence, antioxidant defense and photosynthesis. Int. J. Mol. Sci. 2018, 19, 4045. [Google Scholar] [CrossRef] [Green Version]
- Hai, N.N.; Chuong, N.N.; Tu, N.H.C.; Kisiala, A.; Hoang, X.L.T.; Thao, N.P. Role and regulation of cytokinins in plant response to drought stress. Plants 2020, 9, 422. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ren, Z.; Zhang, J.; Chuang, C.C.; Kandaswamy, E.; Zhou, T.; Zou, L. Role of ROS and Nutritional Antioxidants in Human Diseases. Front. Physiol. 2018, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Jeandet, P.; Formela-Luboińska, M.; Labudda, M.; Morkunas, I. The Role of Sugars in Plant Responses to Stress and Their Regulatory Function during Development. Int. J. Mol. Sci. 2022, 23, 5161. [Google Scholar] [CrossRef]
- Hernández-Herrera, R.M.; Santacruz-Ruvalcaba, F.; Ruiz-López, M.A.; Norrie, J.; Hernández-Carmona, G. Effect of liquid seaweed extracts on growth of tomato seedlings (Solanum lycopersicum L.). J. Appl. Phycol. 2014, 26, 619–628. [Google Scholar] [CrossRef]
- Teixeira, W.F.; Fagan, E.B.; Soares, L.H.; Umburanas, R.C.; Reichardt, K.; Neto, D.D. Foliar and seed application of amino acids affects the antioxidant metabolism of the soybean crop. Front. Plant Sci. 2017, 8, 327. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Li, Z.; Du, Y.; Duan, J.; Zhang, T.; Wei, Z.; Guo, L.; Gong, W.; Liu, Z.; Zhang, M. The Combined Application of Urea and Fulvic Acid Solution Improved Maize Carbon and Nitrogen Metabolism. Agronomy 2022, 12, 1400. [Google Scholar] [CrossRef]
- Clavijo-Herrera, J.; van Santen, E.; Gómez, C. Growth, water-use efficiency, stomatal conductance, and nitrogen uptake of two lettuce cultivars grown under different percentages of blue and red light. Horticulturae 2018, 4, 16. [Google Scholar] [CrossRef]
- Iseki, K.; Olaleye, O. A new indicator of leaf stomatal conductance based on thermal imaging for field grown cowpea. Plant Prod. Sci. 2020, 23, 136–147. [Google Scholar] [CrossRef] [Green Version]
- Daszkowska-Golec, A.; Szarejko, I. Open or close the gate–Stomata action under the control of phytohormones in drought stress conditions. Front. Plant Sci. 2013, 4, 138. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; McAdam, S.A.M.; Carins Murphy, M.R. Xylem and stomata, coordinated through time and space. Plant Cell Environ. 2017, 40, 872–880. [Google Scholar] [CrossRef]
- Miceli, A.; Vetrano, F.; Moncada, A. Influence of Ecklonia maxima extracts on growth, yield, and postharvest quality of hydroponic leaf lettuce. Horticulturae 2021, 7, 440. [Google Scholar] [CrossRef]
- Carrillo, P.; Ciarmiello, L.F.; Woodrow, P.; Corrado, G.; Chiaiese, P.; Rouphael, Y. Enhancing Sustainability by Improving Plant Salt Tolerance through Macro- and Micro-Algal Biostimulants. Biology 2020, 9, 253. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Samuels, L.J.; Setati, M.E.; Blancquaert, E.H. Toward a Better Understanding of the Potential Benefits of Seaweed Based Biostimulants in Vitis vinifera L. Cultivars. Plants 2022, 11, 348. [Google Scholar] [CrossRef]
- Karimi, S.; Karami, H.; Mokhtassi-Bidgoli, A.; Tavallali, V.; Vahdati, K. Inducing drought tolerance in greenhouse grown Juglans regia by imposing controlled salt stress: The role of osmotic adjustment. Sci. Hortic. 2018, 239, 181–192. [Google Scholar] [CrossRef]
- Queiroz Zepka, L.; Jacob-Lopes, E.; Roca, M. Catabolism and bioactive properties of chlorophylls. Curr. Opin. Food Sci. 2019, 26, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Collini, E. Carotenoids in Photosynthesis: The Revenge of the “Accessory” Pigments. Chem 2019, 5, 494–495. [Google Scholar] [CrossRef]
- Domonkos, I.; Kis, M.; Gombos, Z.; Ughy, B. Carotenoids, versatile components of oxygenic photosynthesis. Prog. Lipid Res. 2013, 52, 539–561. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, H.; Gonzáles-Morales, S.; Benavides-Mendoza, A.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Juárez-Maldonado, A. Effects of Chitosan–PVA and Cu Nanoparticles on the Growth and Antioxidant Capacity of Tomato under Saline Stress. Molecules 2018, 23, 178. [Google Scholar] [CrossRef] [Green Version]
- Morales-Espinoza, M.C.; Cadenas-Pliego, G.; Pérez-Alvarez, M.; Hernández-Fuentes, A.D.; Cabrera de la Fuente, M.; Benavides-Mendoza, A.; Valdés-Reyna, J.; Juárez-Maldonado, A. Se Nanoparticles Induce Changes in the Growth, Antioxidant Responses, and Fruit Quality of Tomato Developed under NaCl Stress. Molecules 2019, 24, 3030. [Google Scholar] [CrossRef] [Green Version]
- Vijayanand, N.; Ramya, S.S.; Rathinavel, S. Potential of liquid extracts of Sargassum wightii on growth, biochemical and yield parameters of cluster bean plant. Asian Pacific J. Reprod. 2014, 3, 150–155. [Google Scholar] [CrossRef]
- Khoshbakht, D.; Ramin, A.A.; Baninasab, B. Effects of sodium chloride stress on gas exchange, chlorophyll content and nutrient concentrations of nine citrus rootstocks. Photosynthetica 2015, 53, 241–249. [Google Scholar] [CrossRef]
- Latsague, M.; Sáez, P.; Mora, M. Efecto de la fertilización con nitrógeno, fósforo y potasio, sobre el contenido foliar de carbohidratos, proteínas y pigmentos fotosintéticos en plantas de Berberidopsis corallina Hook.f. Gayana Bot. 2014, 71, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, E.; Ruiz, J.M.; Romero, L.; Preciado-Rangel, P.; Flores-Córdova, M.A.; Márquez-Quiroz, C. ¿Son los pigmentos fotosintéticos buenos indicadores de la relación del nitrógeno, fósforo y potasio en frijol ejotero? Ecosistemas Y Recur. Agropecu. 2018, 5, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Estaji, A.; Kalaji, H.M.; Karimi, H.R.; Roosta, H.R.; Moosavi-Nezhad, S.M. How glycine betaine induces tolerance of cucumber plants to salinity stress? Photosynthetica 2019, 57, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Morales, S.; López-Sánchez, R.C.; Juárez-Maldonado, A.; Robledo-Olivo, A.; Benavides Mendoza, A. A Transcriptomic and Proteomic View of Hydrogen Sulfide Signaling in Plant Abiotic Stress. In Hydrogen Sulfide and Plant Acclimation to Abiotic Stresses; Khan, M., Siddiqui, M., Alamri, S., Corpas, F., Eds.; Springer: Cham, Switzerland, 2021; Volume 1, pp. 161–186. ISBN 978-3-030-73678-1. [Google Scholar]
- Oberkofler, V.; Pratx, L.; Bäurle, I. Epigenetic regulation of abiotic stress memory: Maintaining the good things while they last. Curr. Opin. Plant Biol. 2021, 61, 102007. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Jeon, B.W.; Kim, J. Signaling Peptides Regulating Abiotic Stress Responses in Plants. Front. Plant Sci. 2021, 12, 704490. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Joshi, S.; Patil, S.; Khare, T.; Kumar, V. Reactive Oxygen, Nitrogen, Carbonyl and Sulfur Species and Their Roles in Plant Abiotic Stress Responses and Tolerance. J. Plant Growth Regul. 2021, 3, 1–24. [Google Scholar] [CrossRef]
- Haida, Z.; Hakiman, M. A comprehensive review on the determination of enzymatic assay and nonenzymatic antioxidant activities. Food Sci. Nutr. 2019, 7, 1555–1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defense mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-Y.; Huang, Y.-H.; Yeh, H.-H.; Hong, P.-Y.; Hsiao, C.-J.; Hsieh, L.-S. Phenylalanine, tyrosine, and DOPA are bona fide substrates for Bambusa oldhamii BoPAL4. Catalysts 2021, 11, 1263. [Google Scholar] [CrossRef]
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar] [CrossRef]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed extracts enhance salam turfgrass performance during prolonged irrigation intervals and saline shock. Front. Plant Sci. 2017, 8, 830. [Google Scholar] [CrossRef] [Green Version]
- Aitouguinane, M.; Bouissil, S.; Mouhoub, A.; Rchid, H.; Fendri, I.; Abdelkafi, S.; Ould El-Hadj, M.D.; Boual, Z.; Dubessay, P.; Gardarin, C.; et al. Induction of Natural Defenses in Tomato Seedlings by Using Alginate and Oligoalginates Derivatives Extracted from Moroccan Brown Algae. Mar. Drugs 2020, 18, 521. [Google Scholar] [CrossRef]
- Hashem, H.A.; Mansour, H.A.; El-Khawas, S.A.; Hassanein, R.A. The potentiality of marine macroalgae as biofertilizers to improve the productivity and salt stress tolerance of canola (Brassica napus L.) plants. Agronomy 2019, 9, 146. [Google Scholar] [CrossRef] [Green Version]
- Kumari, R.; Kaur, I.; Bhatnagar, A.K. Effect of aqueous extract of Sargassum johnstonii Setchell & Gardner on growth, yield and quality of Lycopersicon esculentum Mill. J. Appl. Phycol. 2011, 23, 623–633. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Alotaibi, M.; Alhammad, B.A.; Rady, M.M.; Mahdi, A.H.A. Exogenous potassium treatments elevate salt tolerance and performances of Glycine max L. by boosting antioxidant defense system under actual saline field conditions. Agronomy 2020, 10, 1741. [Google Scholar] [CrossRef]
- Taha, R.S.; Seleiman, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H.A. Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions. Agronomy 2021, 11, 74. [Google Scholar] [CrossRef]
- Shukla, P.S.; Mantin, E.G.; Adil, M.; Bajpai, S.; Critchley, A.T.; Prithiviraj, B. Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 2019, 10, 655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, J.K.; Philippe, R.N.; Noel, J.P. The rise of chemodiversity in plants. Science 2012, 336, 1667–1670. [Google Scholar] [CrossRef] [Green Version]
- Dhara, A.; Raichaudhuri, A. ABCG transporter proteins with beneficial activity on plants. Phytochemistry 2021, 184, 112663. [Google Scholar] [CrossRef]
- Prins, M.; Laimer, M.; Noris, E.; Schubert, J.; Wassenegger, M.; Tepfer, M. Strategies for antiviral resistance in transgenic plants. Mol. Plant Pathol. 2008, 9, 73–83. [Google Scholar] [CrossRef]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212–213, 29–37. [Google Scholar] [CrossRef]
- Radhakrishnan, R. Seed pretreatment with magnetic field alters the storage proteins and lipid profiles in harvested soybean seeds. Physiol. Mol. Biol. Plants 2018, 24, 343–347. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant-and seaweed-based extracts increase yield but differentially modulate nutritional quality of greenhouse spinach through biostimulant action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.T.; Bhowmik, S.D.; Long, H.; Cheng, Y.; Mundree, S.; Hoang, L.T.M. Rapid Accumulation of Proline Enhances Salinity Tolerance in Australian Wild Rice Oryza australiensis Domin. Plants 2021, 10, 2044. [Google Scholar] [CrossRef] [PubMed]
- El-Beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Role of Ascorbic acid, Glutathione and Proline Applied as Singly or in Sequence Combination in Improving Chickpea Plant through Physiological Change and Antioxidant Defense under Different Levels of Irrigation Intervals. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, P.; Yang, X.; Yuan, Y.; Jing, C.; Cao, J.; Wang, Y.; Zhang, L.; Zhang, C.; Li, Y. Purification and characterization of a fucoidan from the brown algae Macrocystis pyrifera and the activity of enhancing salt-stress tolerance of wheat seedlings. Int. J. Biol. Macromol. 2021, 180, 547–558. [Google Scholar] [CrossRef]
- Chun, S.C.; Paramasivan, M.; Chandrasekaran, M. Proline Accumulation Influenced by Osmotic Stress in Arbuscular Mycorrhizal Symbiotic Plants. Front. Microbiol. 2018, 9, 2525. [Google Scholar] [CrossRef] [Green Version]
- Arnao, M.B.; Hernández-Ruiz, J.; Cano, A.; Reiter, R.J. Melatonin and carbohydrate metabolism in plant cells. Plants 2021, 10, 1917. [Google Scholar] [CrossRef]
- Mao, Y.; Chen, N.; Cao, M.; Chen, R.; Guan, X.; Wang, D. Functional Characterization and Evolutionary Analysis of Glycine-Betaine Biosynthesis Pathway in Red Seaweed Pyropia yezoensis. Mar. Drugs 2019, 17, 70. [Google Scholar] [CrossRef] [Green Version]
- Khan, T.A.; Yusuf, M.; Fariduddin, Q. Hydrogen peroxide in regulation of plant metabolism: Signaling and its effect under abiotic stress. Photosynthetica 2018, 56, 1237–1248. [Google Scholar] [CrossRef]
- Aerts, N.; Pereira Mendes, M.; Van Wees, S.C.M. Multiple levels of crosstalk in hormone networks regulating plant defense. Plant J. 2021, 105, 489–504. [Google Scholar] [CrossRef]
- MacKelprang, R.; Lemaux, P.G. Genetic Engineering and Editing of Plants: An Analysis of New and Persisting Questions. Annu. Rev. Plant Biol. 2020, 71, 659–687. [Google Scholar] [CrossRef] [Green Version]
- Medda, S.; Fadda, A.; Mulas, M. Influence of Climate Change on Metabolism and Biological Characteristics in Perennial Woody Fruit Crops in the Mediterranean Environment. Horticulturae 2022, 8, 273. [Google Scholar] [CrossRef]
- Drira, M.; Ben Mohamed, J.; Ben Hlima, H.; Hentati, F.; Michaud, P.; Abdelkafi, S.; Fendri, I. Improvement of Arabidopsis thaliana salt tolerance using a polysaccharidic extract from the brown algae Padina pavonica. Algal Res. 2021, 56, 102324. [Google Scholar] [CrossRef]
- Al-Ghamdi, A.A.; Elansary, H.O. Synergetic effects of 5-aminolevulinic acid and Ascophyllum nodosum seaweed extracts on Asparagus phenolics and stress related genes under saline irrigation. Plant Physiol. Biochem. 2018, 129, 273–284. [Google Scholar] [CrossRef]
- Tuteja, N.; Sopory, S.K. Chemical signaling under abiotic stress environment in plants. Plant Signal. Behav. 2008, 3, 525–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keisham, M.; Mukherjee, S.; Bhatla, S.C. Mechanisms of sodium transport in plants—Progresses and challenges. Int. J. Mol. Sci. 2018, 19, 647. [Google Scholar] [CrossRef] [Green Version]
- Guan, C.; Huang, Y.-H.; Cen, H.-F.; Cui, X.; Tian, D.-Y.; Zhang, Y.-W. Overexpression of the Lolium perenne L. Delta1-pyrroline 5-carboxylate synthase (LpP5CS) gene results in morphological alterations and salinity tolerance in switchgrass (Panicum virgatum L.). PLoS ONE 2019, 14, e0219669. [Google Scholar] [CrossRef] [Green Version]
- Changan, S.S.; Ali, K.; Kumar, V.; Garg, N.K.; Tyagi, A. Abscisic acid biosynthesis under water stress: Anomalous behavior of the 9-cis-epoxycarotenoid dioxygenase1 (NCED1) gene in rice. Biol. Plant. 2018, 62, 663–670. [Google Scholar] [CrossRef]
- ul Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.-X.; Zhang, H.-X.; Wei, A.-M.; Gong, Z.-H. Heat shock proteins: Dynamic biomolecules to counter plant biotic and abiotic stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef] [Green Version]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Ziaf, K.; Munis, M.F.H.; Samin, G.; Zhang, X.; Li, J.; Zhang, J.; Ye, Z. Characterization of ERD15 gene from cultivated tomato (Solanum lycopersicum). Pakistan J. Agric. Sci. 2016, 53, 27–33. [Google Scholar] [CrossRef]
- Steiner, A.A. A universal method for preparing nutrient solutions of a certain desired composition. Plant Soil 1961, 15, 134–154. [Google Scholar] [CrossRef] [Green Version]
- Ramya, S.S.; Nagaraj, S.; Vijayanand, N. Influence of Seaweed Liquid Extracts on Growth, Biochemical and Yield Characteristics of Cyamopsis tetragonolaba (L.) Taub. J. Phytol. 2011, 3, 37–41. [Google Scholar]
- Kasim, W.A.; Hamada, E.A.M.; El-Din, N.G.S.; Eskander, S. Influence of seaweed extracts on the growth, some metabolic activities and yield of wheat grown under drought stress. Int. J. Agron. Agric. Res. 2015, 7, 173–189. [Google Scholar]
- Ramya, S.S.; Vijayanand, N.; Rathinavel, S. Foliar application of liquid biofertilizer of brown alga Stoechospermum marginatum on growth, biochemical and yield of Solanum melongena. Int. J. Recycl. Org. Waste Agric. 2015, 4, 167–173. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Purification of ascorbate peroxidase in spinach chloroplasts; its inactivation in ascorbate-depleted medium and reactivation by monodehydroascorbate radical. Plant Cell Physiol. 1987, 28, 131–140. [Google Scholar] [CrossRef]
- Sykłowska-Baranek, K.; Pietrosiuk, A.; Naliwajski, M.R.; Kawiak, A.; Jeziorek, M.; Wyderska, S.; Łojkowska, E.; Chinou, I. Effect of L-phenylalanine on PAL activity and production of naphthoquinone pigments in suspension cultures of Arnebia euchroma (Royle) Johnst. Vitro Cell. Dev. Biol. Plant 2012, 48, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Nour, V.; Trandafir, I.; Ionica, M.E. HPLC Organic Acid Analysis in Different Citrus Juices under Reversed Phase Conditions. Not. Bot. Horti Agrobot. Cluj-Napoca 2010, 38, 44–48. [Google Scholar] [CrossRef]
- Xue, T.; Hartikainen, H.; Piironen, V. Antioxidative and growth-promoting effect of selenium on senescing lettuce. Plant Soil 2001, 237, 55–61. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Bates, L.S. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Antoniou, C.; Savvides, A.; Georgiadou, E.C.; Fotopoulos, V. Spectrophotometric Quantification of Reactive Oxygen, Nitrogen and Sulfur Species in Plant Samples. In Polyamines: Methods and Protocols, Methods in Molecular Biology; Alcázar, R., Tiburcio, A.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1694, pp. 155–161. ISBN 9781493973989. [Google Scholar]
- Yemm, E.; Cocking, E. The Determination of Amino-acids with Ninhydrin. Analyst 1955, 80, 209–214. [Google Scholar] [CrossRef]
- Bosco, R.; Caser, M.; Vanara, F.; Scariot, V. Development of a rapid LC-DAD/FLD method for the simultaneous determination of auxins and abscisic acid in plant extracts. J. Agric. Food Chem. 2013, 61, 10940–10947. [Google Scholar] [CrossRef]
- Rivas-Martínez, E.N.; Foroughbakhch-Pournavab, R.; Reyes-Valdés, M.H.; Benavides-Mendoza, A. Determinación de giberelina A4 y trans zeatina ribósido en diferentes órganos de Dasylirion cedrosanum. Rev. Mex. Cienc. Agrícolas 2016, 7, 2063–2069. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Microwave-assisted extraction of sulfated polysaccharides (fucoidan) from brown seaweed. Carbohydr. Polym. 2011, 86, 1137–1144. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Tao, X.; Xie, Y.; Fauquet, C.M.; Zhou, X. A DNAβ Associated with Tomato Yellow Leaf Curl China Virus Is Required for Symptom Induction. J. Virol. 2004, 78, 13966–13974. [Google Scholar] [CrossRef] [Green Version]
- Mascia, T.; Santovito, E.; Gallitelli, D.; Cillo, F. Evaluation of reference genes for quantitative reverse-transcription polymerase chain reaction normalization in infected tomato plants. Mol. Plant Pathol. 2010, 11, 805–816. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Evaluation | Treatment | Seedling Height (cm) | Stem Diameter (mm) | Number of Leaves |
|---|---|---|---|---|
| AC | 12.00 ± 0.73 bc | 3.18 ± 0.23 bc | 6.00 ± 0.00 b | |
| ANCP | 12.66 ± 1.05 b | 3.22 ± 0.27 b | 6.60 ± 0.55 b | |
| 11 DAT | SSE | 14.04 ± 1.43 a | 3.54 ± 0.32 a | 7.40 ± 0.89 a |
| AC + NaCl | 11.18 ± 0.60 c | 2.71 ± 0.14 d | 6.00 ± 0.71 b | |
| ANCP + NaCl | 11.50 ± 0.94 bc | 2.91 ± 0.15 cd | 6.20 ± 0.45 b | |
| SSE + NaCl | 11.72 ± 0.91 bc | 3.11 ± 0.18 bc | 6.40 ± 0.55 b | |
| AC | 24.02 ± 0.84 a | 3.81 ± 0.15 bc | 8.20 ± 0.45 c | |
| ANCP | 25.40 ± 1.91 a | 4.05 ± 0.25 ab | 8.80 ± 0.45 b | |
| 21 DAT | SSE | 26.20 ± 2.79 a | 4.28 ± 0.30 a | 9.60 ± 0.55 a |
| AC + NaCl | 19.18 ± 1.27 b | 3.09 ± 0.14 d | 7.80 ± 0.45 c | |
| ANCP + NaCl | 20.78 ± 1.71 b | 3.16 ± 0.18 d | 8.00 ± 0.00 c | |
| SSE + NaCl | 20.97 ± 0.67 b | 3.55 ± 0.16 c | 8.00 ± 0.00 c | |
| AC | 35.60 ± 0.45 b | 4.51 ± 0.15 c | 10.20 ± 0.45 b | |
| ANCP | 36.80 ± 2.68 ab | 4.83 ± 0.35 b | 11.00 ± 0.71 a | |
| 31 DAT | SSE | 38.38 ± 2.62 a | 5.18 ± 0.30 a | 11.40 ± 0.55 a |
| AC + NaCl | 24.66 ± 1.09 c | 3.49 ± 0.14 e | 8.80 ± 0.45 c | |
| ANCP + NaCl | 26.00 ± 0.60 c | 3.56 ± 0.18 e | 9.00 ± 0.00 c | |
| SSE + NaCl | 26.44 ± 1.26 c | 4.05 ± 0.16 d | 9.20 ± 0.45 c |
| Treatment | FAB | FRB | TFB | DAB | DRB | TDB |
|---|---|---|---|---|---|---|
| (g Plant−1) | (g Plant−1) | (g Plant−1) | (g Plant−1) | (g Plant−1) | (g Plant−1) | |
| AC | 19.58 ± 1.34 b | 3.95 ± 2.24 a–c | 23.53 ± 3.14 b | 3.52 ± 0.14 b | 0.84 ± 0.09 b | 4.36 ± 0.21 b |
| ANCP | 20.49 ± 2.43 b | 5.91 ± 3.55 a | 26.40 ± 5.54 ab | 3.69 ± 0.44 b | 0.90 ± 0.13 b | 4.59 ± 0.57 b |
| SSE | 23.02 ± 1.51 a | 5.04 ± 1.71 ab | 28.06 ± 3.10 a | 4.03 ± 0.33 a | 1.12 ± 0.17 a | 5.15 ± 0.43 a |
| AC + NaCl | 9.32 ± 0.93 c | 1.04 ± 0.57 d | 10.36 ± 1.48 c | 1.55 ± 0.14 c | 0.37 ± 0.09 d | 1.92 ± 0.23 c |
| ANCP + NaCl | 10.04 ± 0.85 c | 2.22 ± 1.41 cd | 12.27 ± 2.23 c | 1.61 ± 0.15 c | 0.43 ± 0.07 cd | 2.04 ± 0.21 c |
| SSE + NaCl | 11.06 ± 1.11 c | 2.43 ± 1.35 b–d | 13.49 ± 2.02 c | 1.72 ± 0.10 c | 0.56 ± 0.15 c | 2.28 ± 0.20 c |
| CV (%) | 9.37 | 59.09 | 16.82 | 9.36 | 17.43 | 10 |
| Composition | Concentration |
|---|---|
| pH | 5.60 ± 0.10 |
| EC (dS m−1) | 0.83 ± 0.01 |
| Total proteins (mg g−1 DW) | 3.47 ± 0.08 |
| GSH (mg g−1 DW) | 3.29 ± 0.02 |
| Amino acids (mg g−1 DW) | 0.43 ± 0.008 |
| Total phenols (mg EAG g−1 DW) | 8.43 ± 0.79 |
| Flavonoids (mg EC g−1 DW) | 2.83 ± 0.04 |
| IAA (mg kg−1 DW) | 0.57 ± 0.07 |
| tZ (µg g−1 DW) | 175.99 ± 7.49 |
| Glucose (mg 100 g−1 DW) | 107.87 ± 0.004 |
| Galactose (mg 100 g−1 DW) | 74.01 ± 0.49 |
| Fucose (mg 100 g−1 DW) | 258.37 ± 9.82 |
| Mannitol (mg 100 g−1 DW) | 29.96 ± 0.40 |
| ACTE DPPH (mg g−1 DW) | 53.54 ± 1.70 |
| Gene | Forward Primer 5′-3′ | Reverse Primer 5′-3′ |
|---|---|---|
| ACT | CCCAGGCACACAGGTGTTA | CAGGAGCAACTCGAAGCTC |
| NCED1 | CTTATTTGGCTATCGCTGAACC | CCTCCAACTTCAAACTCATTGC |
| HSP70 | TGCTGGAGGTGTTATGACCA | GACTCCTCTTGGTGCTGGAG |
| PIP2 | CTGCACCGTTGCTCGATTTT | GCGACAGTGACGTAGAGGAA |
| P5CS1 | CTGTTGTGGCTCGAGCTGAT | GACGACCAACACCTACAGCA |
| ERD15 | AGGCATCAAGTCATCACTCTCTGGT | GAGGTAAATGTGAGTAAGAACCAACG |
| Fe-SOD | CTGGGAATCTATGAAGCCCAACGGA | CAAATTGTGTTGCTGCAGCTGCCTT |
| CAT1 | TCGCGATGGTGCTATGAACA | CTCCCCTGCCTGTTTGAAGT |
| cAPX2 | GTGACCACTTGAGGGACGTGTTTGT | ACCAGAACGCTCCTTGTGGCATCTT |
| PAL5-3 | GGAGGAGAATTTGAAGAATGCTGTG | TCCCTTTCCACCACTTGTAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sariñana-Aldaco, O.; Benavides-Mendoza, A.; Robledo-Olivo, A.; González-Morales, S. The Biostimulant Effect of Hydroalcoholic Extracts of Sargassum spp. in Tomato Seedlings under Salt Stress. Plants 2022, 11, 3180. https://doi.org/10.3390/plants11223180
Sariñana-Aldaco O, Benavides-Mendoza A, Robledo-Olivo A, González-Morales S. The Biostimulant Effect of Hydroalcoholic Extracts of Sargassum spp. in Tomato Seedlings under Salt Stress. Plants. 2022; 11(22):3180. https://doi.org/10.3390/plants11223180
Chicago/Turabian StyleSariñana-Aldaco, Oscar, Adalberto Benavides-Mendoza, Armando Robledo-Olivo, and Susana González-Morales. 2022. "The Biostimulant Effect of Hydroalcoholic Extracts of Sargassum spp. in Tomato Seedlings under Salt Stress" Plants 11, no. 22: 3180. https://doi.org/10.3390/plants11223180