
Biochemical and Transcriptional Responses in Cold-Acclimated and Non-Acclimated Contrasting Camelina Biotypes under Freezing Stress

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
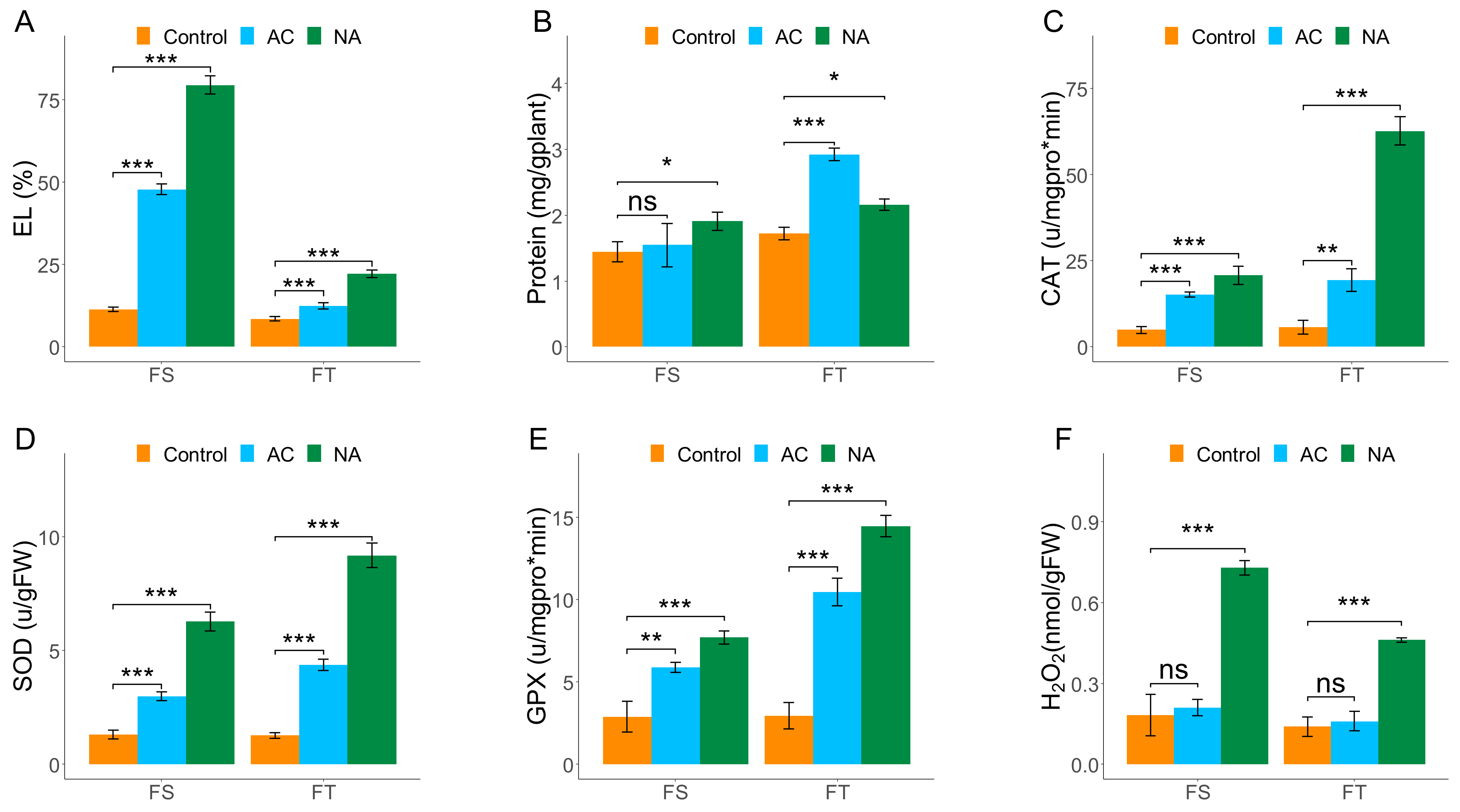
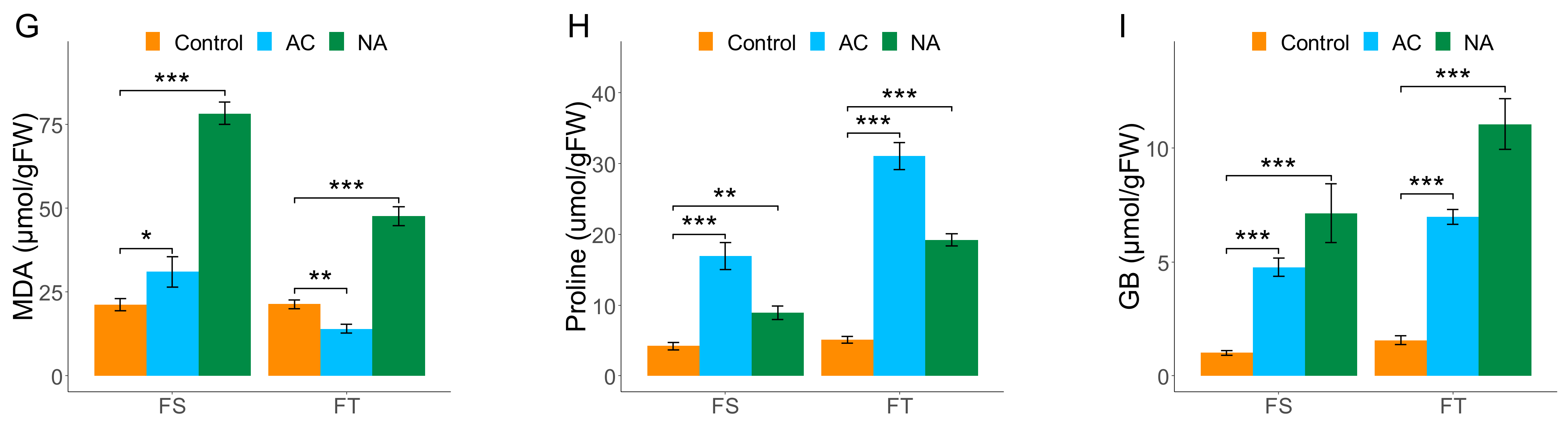
2.1. Biochemical Assessments
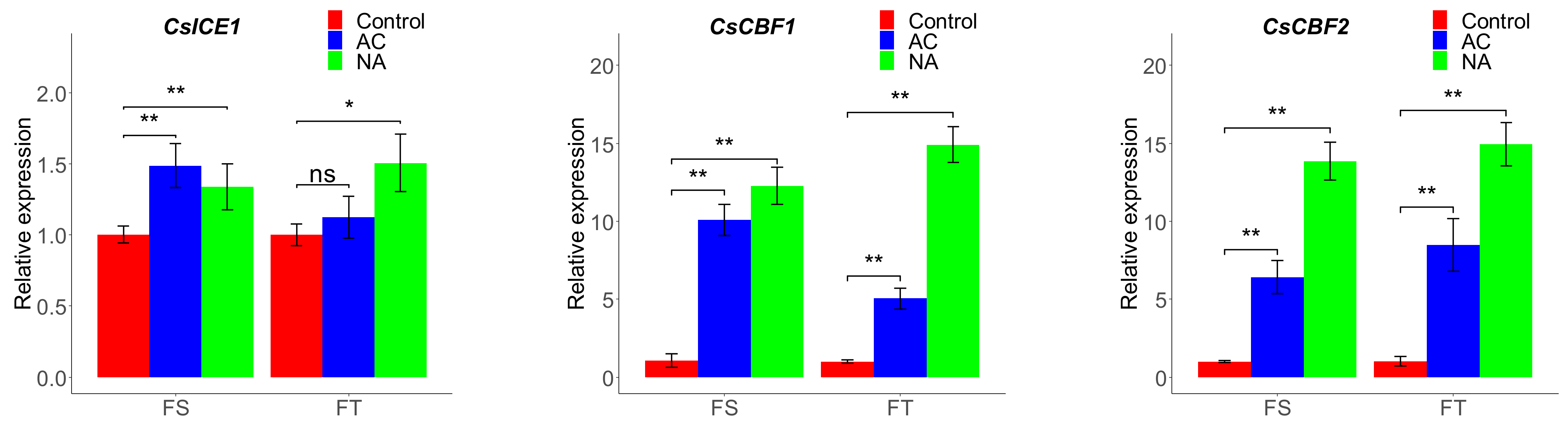
2.2. Expression Profiling of ICE, CBF, and COR Genes
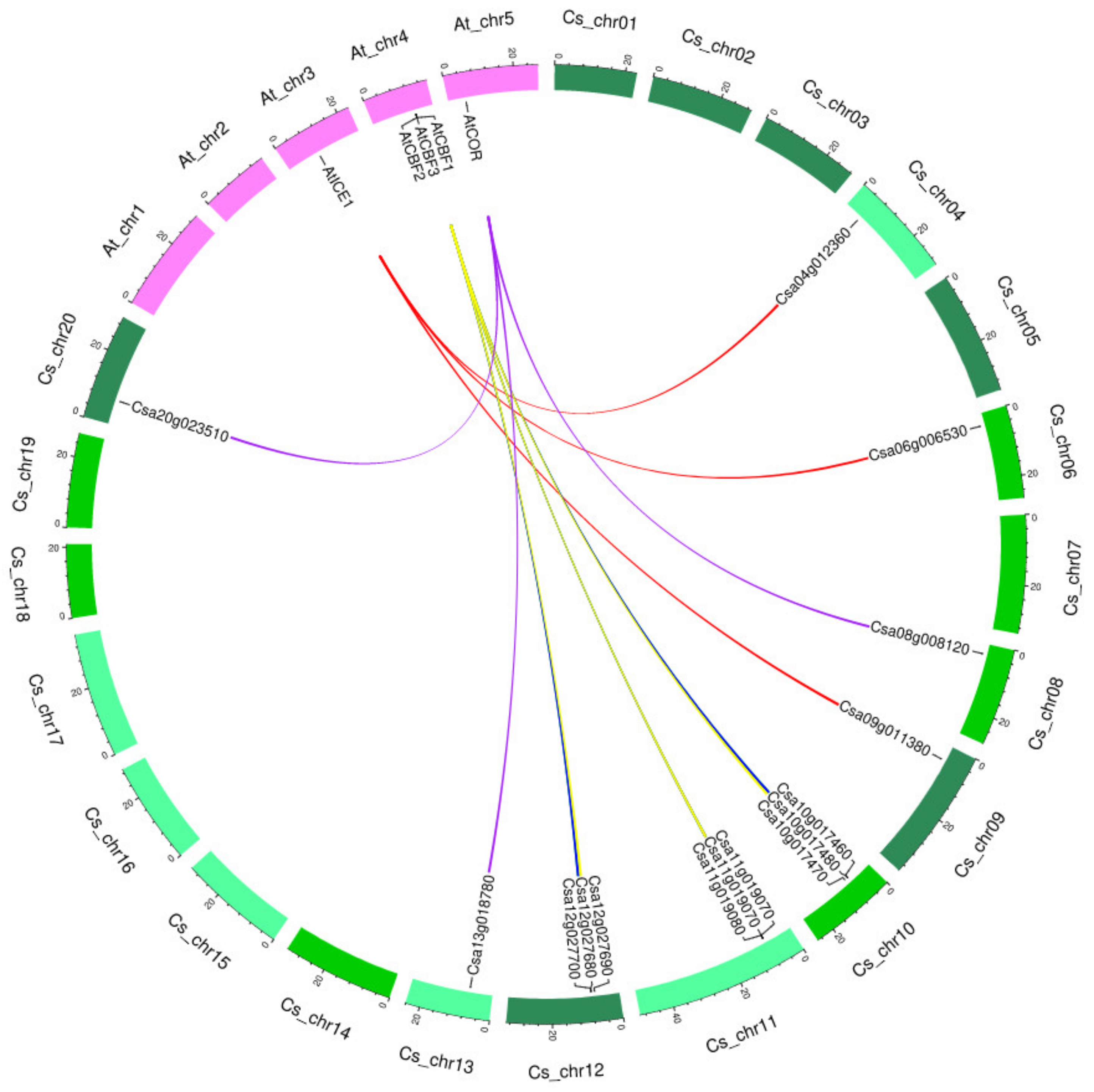
2.3. Identification of Syntelogs and Gene Duplication Analysis
3. Discussion
4. Materials and Methods
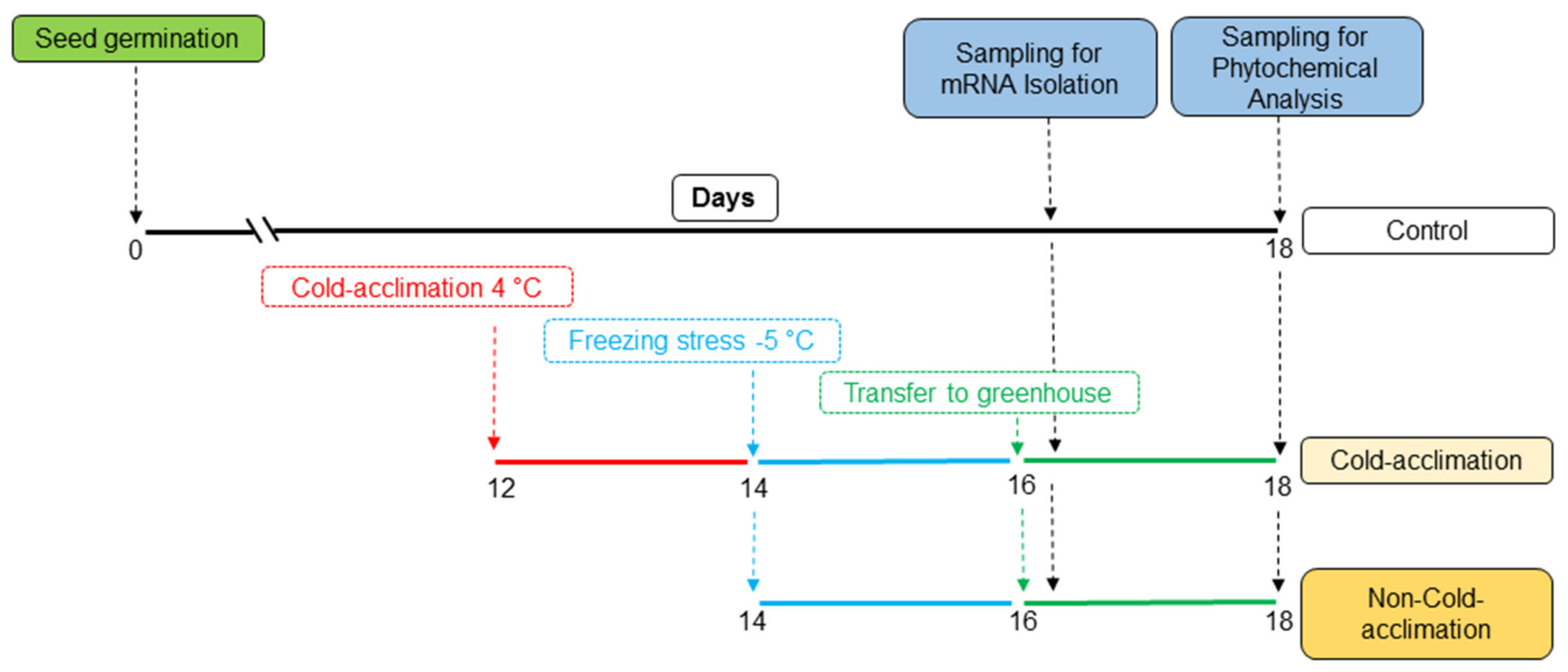
4.1. Plant Materials and Experimental Treatments
4.2. Electrolyte Leakage (EL)
4.3. Preparation of Enzyme Extracts and Antioxidant Enzymes Activity
4.4. Total Soluble Protein
4.5. Glycine Betaine (GB)
4.6. Proline Content
4.7. Hydrogen Peroxide (H2O2)
4.8. Estimation of Lipid Peroxidation (MDA)
4.9. Total RNA Extraction and cDNA Synthesis
4.10. Real-Time PCR Conditions (RT-qPCR) and Gene Expression Analysis
4.11. Synteny Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campbell, M. Camelina—An Alternative Oil Crop. In Biokerosene; Kaltschmitt, M., Neuling, U., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 259–275. [Google Scholar] [CrossRef]
- Soorni, J.; Shobbar, Z.S.; Kahrizi, D.; Zanetti, F.; Sadeghi, K.; Rostampour, S.; Kovács, P.G.; Kiss, A.; Mirmazloum, I. Correlational analysis of agronomic and seed quality traits in Camelina sativa doubled haploid lines under rain-fed condition. Agronomy 2022, 12, 359. [Google Scholar] [CrossRef]
- Choi, S.H.; Park, N.; Lee, K.Y.; Missaoui, A.M.; Lee, G.J. Novel genes in response to varying water deficit in oil crop Camelina sativa. Euphytica 2019, 215, 86. [Google Scholar] [CrossRef]
- Nishchenko, L.V.; Hasanuzzaman, M. Enhancement of Abiotic Stress Tolerance in Camelina sativa: Conventional Breeding and Biotechnology. In The Plant Family Brassicaceae; Hasanuzzaman, M., Ed.; Springer: Singapore, 2020; pp. 195–202. [Google Scholar] [CrossRef]
- Soorni, J.; Kazemitabar, S.K.; Kahrizi, D.; Dehestani, A.; Bagheri, N. Genetic analysis of freezing tolerance in camelina [Camelina sativa (L.) Crantz] by diallel cross of winter and spring biotypes. Planta 2021, 253, 9. [Google Scholar] [CrossRef]
- Kim, H.S.; Oh, J.M.; Luan, S.; Carlson, J.E.; Ahn, S.J. Cold stress causes rapid but differential changes in properties of plasma membrane H+-ATPase of camelina and rapeseed. J. Plant Physiol. 2013, 170, 828–837. [Google Scholar] [CrossRef]
- Hayat, F.; Sun, Z.; Ni, Z.; Iqbal, S.; Xu, W.; Gao, Z.; Qiao, Y.; Tufail, M.A.; Jahan, M.S.; Khan, U.; et al. Exogenous melatonin improves cold tolerance of strawberry (Fragaria × ananassa Duch.) through modulation of DREB/CBF-COR pathway and antioxidant defense system. Horticulturae 2022, 8, 194. [Google Scholar] [CrossRef]
- Barrero-Gil, J.; Salinas, J. Post-translational regulation of cold acclimation response. Plant Sci. 2013, 205–206, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Wu, Q.; Teng, L.; Tang, F.; Pi, Z.; Shen, S. Transcriptional regulation of the paper mulberry under cold stress as revealed by a comprehensive analysis of transcription factors. BMC Plant Biol. 2015, 15, 108. [Google Scholar] [CrossRef] [Green Version]
- Mehrotra, S.; Verma, S.; Kumar, S.; Kumari, S.; Mishra, B.N. Transcriptional regulation and signalling of cold stress response in plants: An overview of current understanding. Environ. Exp. Bot. 2020, 180, 104243. [Google Scholar] [CrossRef]
- Rapacz, M.; Jurczyk, B.; Krępski, T.; Płażek, A. C-repeat binding transcription factors from Miscanthus × giganteus and their expression at a low temperature. Ind. Crop Prod. 2018, 113, 283–287. [Google Scholar] [CrossRef]
- Shi, Y.; Huang, J.; Sun, T.; Wang, X.; Zhu, C.; Ai, Y.; Gu, H. The precise regulation of different COR genes by individual CBF transcription factors in Arabidopsis thaliana. J. Integr. Plant Biol. 2017, 59, 118–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Jiang, L.; Wang, F.; Yu, D. Jasmonate regulates the inducer of CBF expression–C-repeat binding factor/DRE binding factor1 cascade and freezing tolerance in Arabidopsis. Plant Cell 2013, 25, 2907–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaglo, K.R.; Kleff, S.; Amundsen, K.L.; Zhang, X.; Haake, V.; Zhang, J.Z.; Thomashow, M.F. Components of the Arabidopsis C-repeat/dehydration-responsive element binding factor cold-response pathway are conserved in Brassica napus and other plant species. Plant Physiol. 2001, 127, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, T.H.; Lee, J.T.; Yang, P.T.; Chiu, L.H.; Charng, Y.; Wang, Y.C.; Chan, M.T. Heterology expression of the Arabidopsis C-repeat/dehydration response element binding factor 1 gene confers elevated tolerance to chilling and oxidative stresses in transgenic tomato. Plant Physiol. 2002, 129, 1086–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomashow, M.F.; Gilmour, S.J.; Stockinger, E.J.; Jaglo-Ottosen, K.R.; Zarka, D.G. Role of the Arabidopsis CBF transcriptional activators in cold acclimation. Physiol. Plant 2001, 112, 171–175. [Google Scholar] [CrossRef]
- Savitch, L.V.; Allard, G.; Seki, M.; Robert, L.S.; Tinker, N.A.; Huner, N.P.; Shinozaki, K.; Singh, J. The effect of overexpression of two Brassica CBF/DREB1-like transcription factors on photosynthetic capacity and freezing tolerance in Brassica napus. Plant Cell Physiol. 2005, 46, 1525–1539. [Google Scholar] [CrossRef]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol. 2006, 47, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Qin, F.; Sakuma, Y.; Li, J.; Liu, Q.; Li, Y.Q.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Cloning and functional analysis of a novel DREB1/CBF transcription factor involved in cold-responsive gene expression in Zea mays L. Plant Cell Physiol. 2004, 45, 1042–1052. [Google Scholar] [CrossRef] [Green Version]
- Vágújfalvi, A.; Aprile, A.; Miller, A.; Dubcovsky, J.; Delugu, G.; Galiba, G.; Cattivelli, L. The expression of several Cbf genes at the Fr-A2 locus is linked to frost resistance in wheat. Mol. Gen. Genet. 2005, 274, 506–514. [Google Scholar] [CrossRef]
- Liu, L.; Li, S.; Guo, J.; Li, N.; Jiang, M.; Li, X. Low temperature tolerance is depressed in wild-type and abscisic acid-deficient mutant barley grown in Cd-contaminated soil. J. Hazard. Mater. 2022, 15, 128489. [Google Scholar] [CrossRef]
- Zhang, X.; Fowler, S.G.; Cheng, H.; Lou, Y.; Rhee, S.Y.; Stockinger, E.J.; Thomashow, M.F. Freezing-sensitive tomato has a functional CBF cold response pathway, but a CBF regulon that differs from that of freezing-tolerant Arabidopsis. Plant J. 2004, 39, 905–919. [Google Scholar] [CrossRef]
- Hui-Ming, G.U.O.; Zhao-Chun, L.I.; Zhang, H.; Yue-Zhi, X.I.N.; CHENG, H.M. Cloning of cotton CBF gene for cold tolerance and its expression in transgenic tobacco. Acta Agron. Sin. 2011, 37, 286–293. [Google Scholar] [CrossRef]
- Huang, B.O.; Jin, L.; Liu, J. Molecular cloning and functional characterization of a DREB1/CBF-like gene (GhDREB1L) from cotton. Sci. China Life Sci. 2007, 50, 7–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Liu, S.; Liu, X.; Chen, Z.; Liu, X.; Pang, Y.; Sun, X.; Tang, K. Molecular cloning and characterization of a CBF gene from Capsella bursa-pastoris. DNA Seq. 2004, 15, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Noman, A.; Kanwal, H.; Khalid, N.; Sanaullah, T.; Tufail, A.; Masood, A.; Sabir, S.U.R.; Aqeel, M.; He, S. Perspective research progress in cold responses of Capsella bursa-pastoris. Front. Plant Sci. 2017, 8, 1388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwarari, D.; Guan, Y.; Ahmad, B.; Movahedi, A.; Min, T.; Hao, Z.; Lu, Y.; Chen, J.; Yang, L. ICE-CBF-COR signaling cascade and its regulation in plants responding to cold stress. Int. J. Mol. Sci. 2022, 23, 1549. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cai, J.; Liu, F.; Dai, T.; Cao, W.; Jiang, D. Cold priming drives the sub-cellular antioxidant systems to protect photosynthetic electron transport against subsequent low temperature stress in winter wheat. Plant Physiol. Biochem. PPB 2014, 82, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Ritonga, F.N.; Chen, S. Physiological and Molecular Mechanism Involved in Cold Stress Tolerance in Plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Yerlikaya, B.A.; Ömezli, S.; Aydoğan, N. Environment, Climate, Plant and Vegetation Growth. In Climate Change Forecasting and Modeling for the Year of 2050; Springer: Cham, Switzerland, 2020; pp. 109–122. [Google Scholar] [CrossRef]
- Walia, M.K.; Zanetti, F.; Gesch, R.W.; Krzyżaniak, M.; Eynck, C.; Puttick, D.; Alexopoulou, E.; Royo-Esnal, A.; Stolarski, M.J.; Isbell, T.; et al. Winter camelina seed quality in different growing environments across Northern America and Europe. Ind. Crops Prod. 2021, 169, 113639. [Google Scholar] [CrossRef]
- Wang, H.; Doğramacı, M.; Anderson, J.V.; Horvath, D.P.; Chao, W.S. Transcript profiles differentiate cold acclimation-induced processes in a summer and winter biotype of Camelina. Plant Mol. Biol. Rep. 2022, 40, 359–375. [Google Scholar] [CrossRef]
- Wittenberg, A.; Anderson, J.V.; Berti, M.T. Winter and summer annual biotypes of camelina have different morphology and seed characteristics. Ind. Crops Prod. 2019, 135, 230–237. [Google Scholar] [CrossRef]
- Chao, W.S.; Wang, H.; Horvath, D.P.; Anderson, J.V. Selection of endogenous reference genes for qRT-PCR analysis in Camelina sativa and identification of FLOWERING LOCUS C allele-specific markers to differentiate summer-and winter-biotypes. Ind. Crops Prod. 2019, 129, 495–502. [Google Scholar] [CrossRef]
- Thalhammer, A.; Pagter, M.; Hincha, D.K.; Zuther, E. Measuring Freezing Tolerance of Leaves and Rosettes: Electrolyte Leakage and Chlorophyll Fluorescence Assays. In Plant Cold Acclimation; Hincha, D., Zuther, E., Eds.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2020; Volume 2156. [Google Scholar] [CrossRef]
- Soorni, J.; Kazemitabar, S.K.; Kahrizi, D.; Dehestani, A.; Bagheri, N. Screening of camelina (Camelina sativa L.) doubled haploid lines for freezing tolerance in the seedling stage. Genetika 2017, 49, 173–181. [Google Scholar] [CrossRef]
- Arslan, Ö.; Eyidoğan, F.; Ekmekçi, Y. Freezing tolerance of chickpea: Biochemical and molecular changes at vegetative stage. Biol. Plant 2018, 62, 140–148. [Google Scholar] [CrossRef]
- Dreyer, A.; Dietz, K.J. Reactive oxygen species and the redox-regulatory network in cold stress acclimation. Antioxidants 2018, 7, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, W.; Rajashekar, C.B. Glycine betaine involvement in freezing tolerance and water stress in Arabidopsis thaliana. Environ. Exp. Bot. 2001, 46, 21–28. [Google Scholar] [CrossRef]
- Kaur, G.; Kumar, S.; Thakur, P.; Malik, J.A.; Bhandhari, K.; Sharma, K.D.; Nayyar, H. Involvement of proline in response of chickpea (Cicer arietinum L.) to chilling stress at reproductive stage. Sci. Hortic. 2011, 128, 174–181. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold stress regulation of gene expression in plants. Trends Plant Sci. 2007, 12, 444–451. [Google Scholar] [CrossRef]
- Knight, M.R.; Knight, H. Low-temperature perception leading to gene expression and cold tolerance in higher plants. N. Phytol. 2012, 195, 737–751. [Google Scholar] [CrossRef]
- Novillo, F.; Alonso, J.M.; Ecker, J.R.; Salinas, J. CBF2/DREB1C is a negative regulator of CBF1/DREB1B and CBF3/DREB1A expression and plays a central role in stress tolerance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 3985–3990. [Google Scholar] [CrossRef] [Green Version]
- Novillo, F.; Medina, J.; Salinas, J. Arabidopsis CBF1 and CBF3 have a different function than CBF2 in cold acclimation and define different gene classes in the CBF regulon. Proc. Natl. Acad. Sci. USA 2007, 104, 21002–21007. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Liu, Y.; Mu, Y.; Anwar, A.; He, C.; Yan, Y.; Li, Y.; Yu, X. Heterotrimeric G-protein γ subunit CsGG3.2 positively regulates the expression of CBF genes and chilling tolerance in cucumber. Front. Plant Sci. 2018, 9, 488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Ding, Y.; Shi, Y.; Zhang, X.; Gong, Z.; Yang, S. The cbfs triple mutants reveal the essential functions of CBFs in cold acclimation and allow the definition of CBF regulons in Arabidopsis. N. Phytol. 2016, 212, 345–353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Zhang, Z.; Xie, S.; Si, T.; Li, Y.; Zhu, J.K. Mutational evidence for the critical role of CBF genes in cold acclimation in Arabidopsis. Plant Physiol. 2016, 171, 2744–2759. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, L.; Wang, Y.; Liu, H.; Hu, D.; Zhang, N.; Zhang, S.; Cao, H.; Cao, Q.; Zhang, Z.; et al. Arabidopsis PcaP2 plays an important role in chilling tolerance and ABA response by activating CBF-and SnRK2-mediated transcriptional regulatory network. Front. Plant Sci. 2018, 9, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.V.; Neubauer, M.; Horvath, D.P.; Chao, W.S.; Berti, M.T. Analysis of Camelina sativa transcriptomes identified specific transcription factors and processes associated with freezing tolerance in a winter biotype. Ind. Crops Prod. 2022, 177, 114414. [Google Scholar] [CrossRef]
- Horvath, D.; Anderson, J.V.; Chao, W.S.; Zheng, P.; Buchwaldt, M.; Parkin, I.A.; Dorn, K. Genes associated with chloroplasts and hormone-signaling, and transcription factors other than CBFs are associated with differential survival after low temperature treatments of Camelina sativa biotypes. PloS ONE 2019, 14, 0217692. [Google Scholar] [CrossRef]
- Tuteja, N.; Gill, S.S.; Tiburcio, A.F.; Tuteja, R. Improving Crop Resistance to Abiotic Stress; John Wiley & Sons: Hoboken, NJ, USA; Cambridge University Press: Cambridge, UK, 2012; 1534. [Google Scholar] [CrossRef]
- Cortés, A.J.; López-Hernández, F.; Blair, M.W. Genome–Environment Associations, an Innovative Tool for Studying Heritable Evolutionary Adaptation in Orphan Crops and Wild Relatives. Front. Genet. 2022, 13, 910386. [Google Scholar] [CrossRef]
- Cortés, A.J.; López-Hernández, F.; Osorio-Rodriguez, D. Predicting thermal adaptation by looking into populations’ genomic past. Front. Genet. 2020, 11, 564515. [Google Scholar] [CrossRef]
- Buitrago-Bitar, M.A.; Cortés, A.J.; López-Hernández, F.; Londoño-Caicedo, J.M.; Muñoz-Florez, J.E.; Muñoz, L.C.; Blair, M.W. Allelic Diversity at Abiotic Stress Responsive Genes in Relationship to Ecological Drought Indices for Cultivated Tepary Bean, Phaseolus acutifolius A. Gray, and Its Wild Relatives. Genes 2021, 12, 556. [Google Scholar] [CrossRef]
- López-Hernández, F.; Cortés, A.J. Last-Generation Genome–Environment Associations Reveal the Genetic Basis of Heat Tolerance in Common Bean (Phaseolus vulgaris L.). Front. Genet. 2019, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress in relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Chance, B.; Maehly, A.C. [136] Assay of catalases and peroxidases. Methods Enzymol. 1955, 2, 764–775. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: II. Purification and quantitative relationship with water-soluble protein in seedlings. Plant Physiol. 1977, 59, 315–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid assay for determination of water soluble quaternary ammonium compounds. Plant Soil. 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Kellős, T.; Tímár, I.; Szilágyi, V.; Szalai, G.; Galiba, G.; Kocsy, G. Stress hormones and abiotic stresses have different effects on antioxidants in maize lines with different sensitivity. Plant Biol. 2008, 10, 563–572. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Oszlányi, R.; Mirmazloum, I.; Pónya, Z.; Szegő, A.; Jamal, S.; Bat-Erdene, O.; Papp, I. Oxidative stress level and dehydrin gene expression pattern differentiate two contrasting cucumber F1 hybrids under high fertigation treatment. Int. J. Biol. Macromol. 2020, 161, 864–874. [Google Scholar] [CrossRef]
- Jaakola, L.; Pirttilä, A.M.; Halonen, M.; Hohtola, A. Isolation of high quality RNA from bilberry (Vaccinium myrtillus L.) fruit. Mol. Biotechnol. 2001, 19, 201–203. [Google Scholar] [CrossRef]
- Rstudio Team. Rstudio: Integrated Development Environment for R; Rstudio, Inc.: Boston, MA, USA, 2015; Available online: http://www.rstudio.com/ (accessed on 1 March 2022).
- Bolser, D.M.; Staines, D.M.; Perry, E.; Kersey, P.J. Ensembl plants: Integrating tools for visualizing, mining, and analyzing plant genomic data. In Plant Genomics Databases; Humana Press: New York, NY, USA, 2017; pp. 1–31. [Google Scholar] [CrossRef]
- Gomez-Cano, F.; Carey, L.; Lucas, K.; García Navarrete, T.; Mukundi, E.; Lundback, S.; Schnell, D.; Grotewold, E. CamRegBase: A gene regulation database for the biofuel crop Camelina sativa. Database 2020, 2020, baaa075. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ouyang, Y.; Yao, W. ShinyCircos: An R/Shiny application for interactive creation of Circos plot. Bioinformatics 2018, 34, 1229–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: http://www.R-project.org (accessed on 1 March 2022).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | At Gene Stable ID | Chr (n = 5) | Length bp (aa) | Cs Gene Stable ID | Chr (n = 20) | Length bp (aa) | E-Value | Identity (%) | |
|---|---|---|---|---|---|---|---|---|---|
| ICE1 | AT3G26744 | 3 | 2612 (494) | Csa09g011380 | 9 | 1497 (255) | 4.67 × 10−3 | 96.64 | |
| Csa06g006530 | 6 | 2171 (499) | 0 | 93.79 | |||||
| Csa04g012360 | 4 | 2581 (349) | 0 | 93.57 | |||||
| CBF1 (CsDREB1b) | AT4G25490 | 4 | 1216 (213) | Csa10g017460 | 10 | 1218 (212) | 3.33 × 10−127 | 85.85 | |
| Csa12g027680 | 12 | 929 (211) | 3.27 × 10−127 | 86.26 | |||||
| NA | - | - | - | - | |||||
| CBF2 (CsDREB1c) | AT4G25470 | 4 | 985 (216) | Csa11g019080 | 11 | 2540 (217) | 1.37 × 10−71 | 53.16 | |
| Csa10g017480 | 10 | 1008 (218) | 1.11 × 10−132 | 83.49 | |||||
| Csa12g027700 | 12 | 1008 (217) | 8.46 × 10−94 | 80.77 | |||||
| CBF3 (CsDREB1a) | AT4G25480 | 4 | 1390 (216) | Csa12g027690 | 12 | 995 (218) | 6.83 × 10−129 | 84.16 | |
| Csa11g019070 | 11 | 1093 (129) | 7.78 × 10−97 | 85.15 | |||||
| Csa10g017470 | 10 | 953 (218) | 6.33 × 10−130 | 83.27 | |||||
| COR6.6 | AT5G15970 | 5 | 1024 (66) | Csa08g008120 | 8 | 1428 (66) | 9.19 × 10−24 | 93.94 | |
| Csa13g018780 | 13 | 1619 (66) | 9.19 × 10−24 | 93.94 | |||||
| Csa20g023510 | 20 | 845 (66) | 3.14 × 10−23 | 92.42 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soorni, J.; Kazemitabar, S.K.; Kahrizi, D.; Dehestani, A.; Bagheri, N.; Kiss, A.; Kovács, P.G.; Papp, I.; Mirmazloum, I. Biochemical and Transcriptional Responses in Cold-Acclimated and Non-Acclimated Contrasting Camelina Biotypes under Freezing Stress. Plants 2022, 11, 3178. https://doi.org/10.3390/plants11223178
Soorni J, Kazemitabar SK, Kahrizi D, Dehestani A, Bagheri N, Kiss A, Kovács PG, Papp I, Mirmazloum I. Biochemical and Transcriptional Responses in Cold-Acclimated and Non-Acclimated Contrasting Camelina Biotypes under Freezing Stress. Plants. 2022; 11(22):3178. https://doi.org/10.3390/plants11223178
Chicago/Turabian StyleSoorni, Jahad, Seyed Kamal Kazemitabar, Danial Kahrizi, Ali Dehestani, Nadali Bagheri, Attila Kiss, Péter Gergő Kovács, István Papp, and Iman Mirmazloum. 2022. "Biochemical and Transcriptional Responses in Cold-Acclimated and Non-Acclimated Contrasting Camelina Biotypes under Freezing Stress" Plants 11, no. 22: 3178. https://doi.org/10.3390/plants11223178