
Nematicidal, Acaricidal and Plant Growth-Promoting Activity of Enterobacter Endophytic Strains and Identification of Genes Associated with These Biological Activities in the Genomes
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Genome Assembly and Annotation
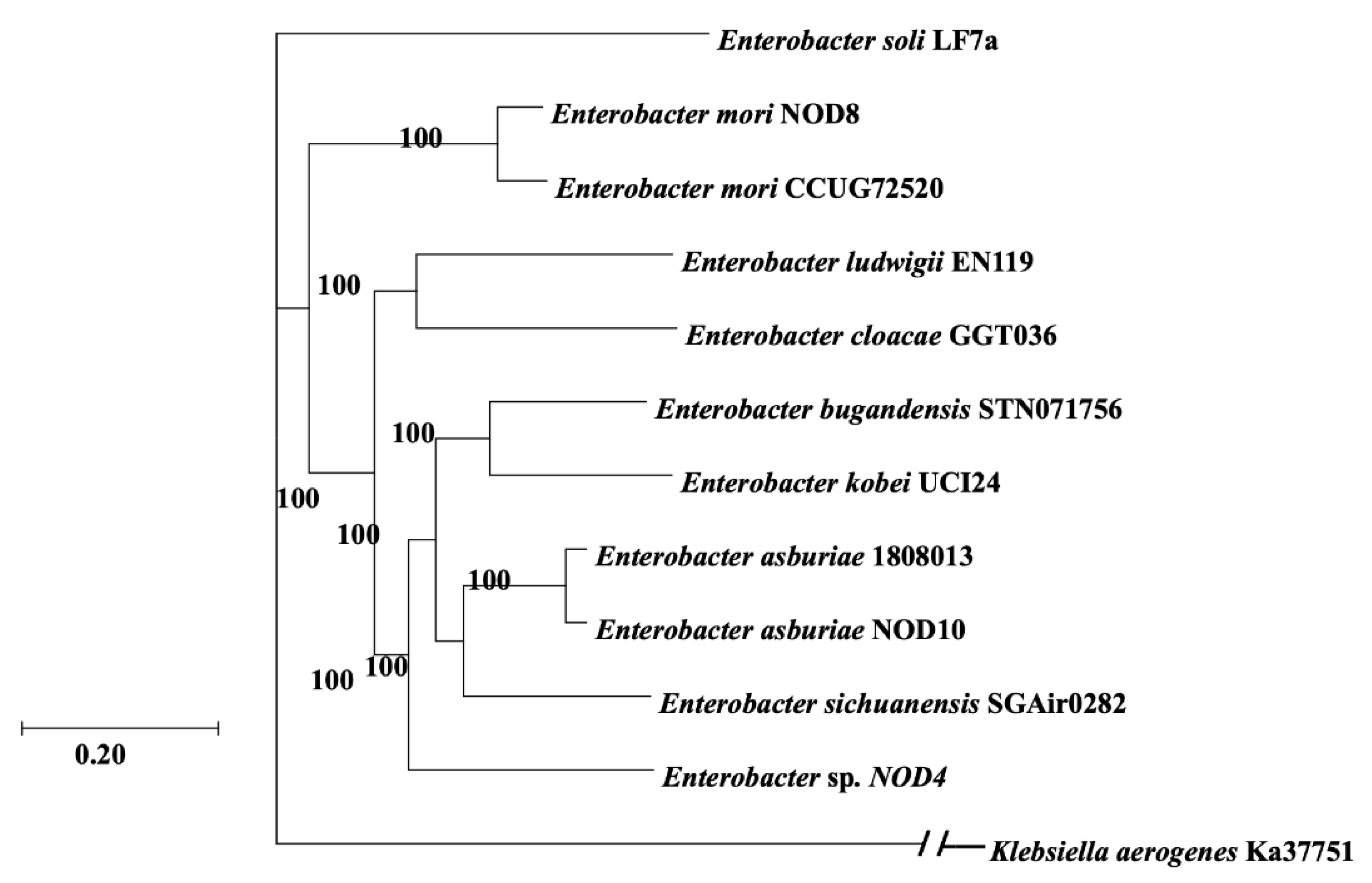
2.2. Phylogenomic Analysis
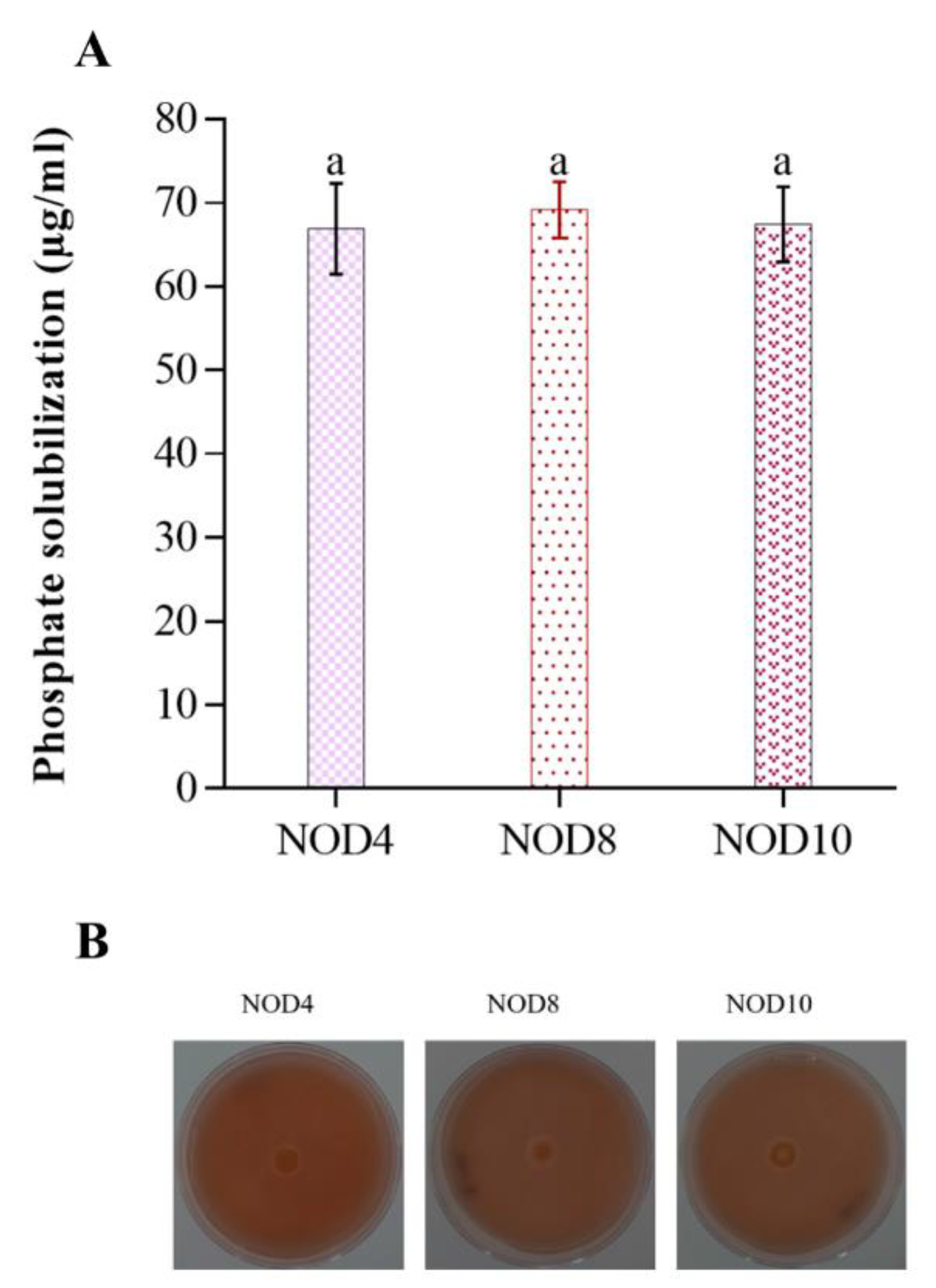
2.3. Phosphate Solubilization Activity
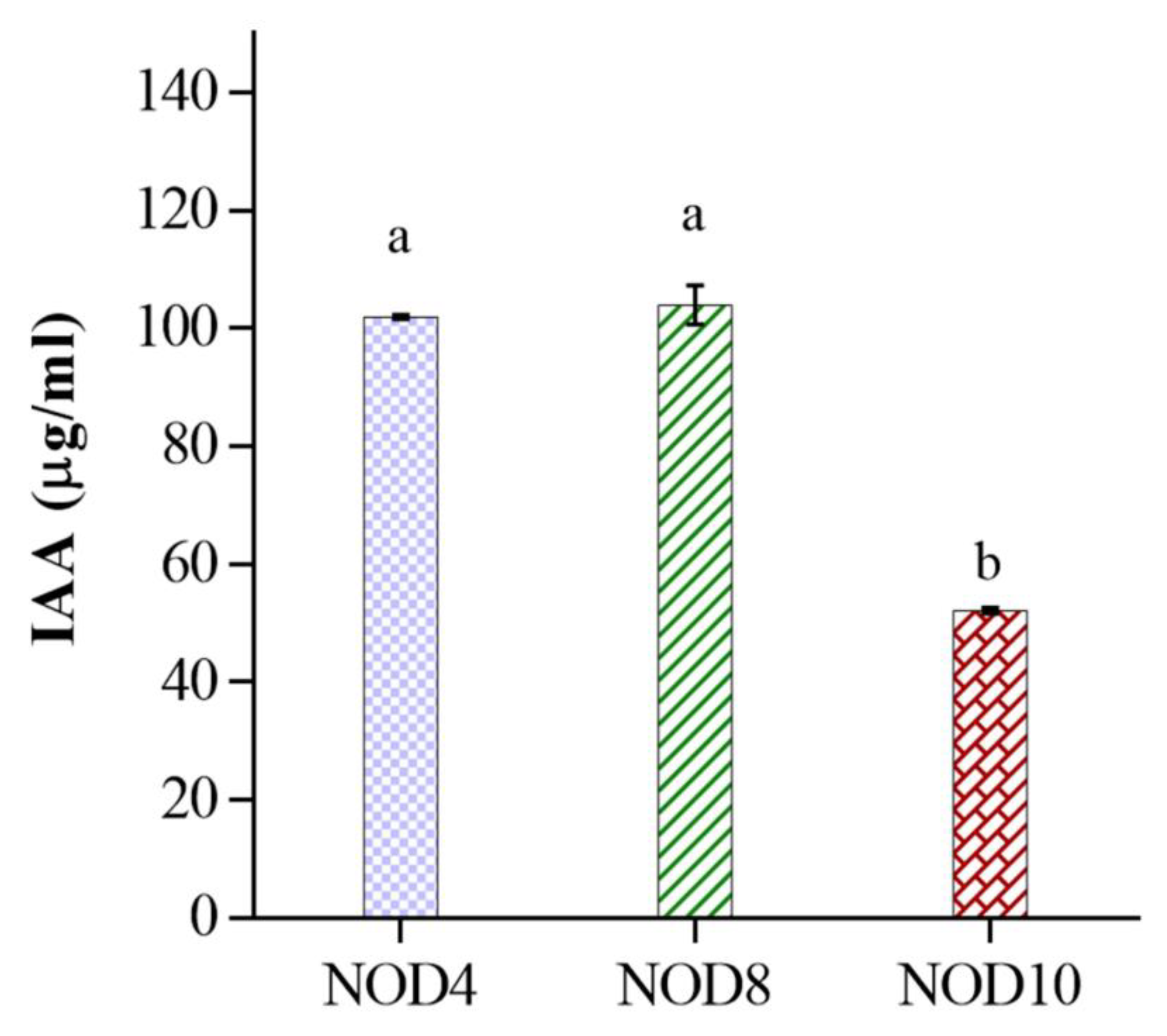
2.4. Determination of Auxin Production
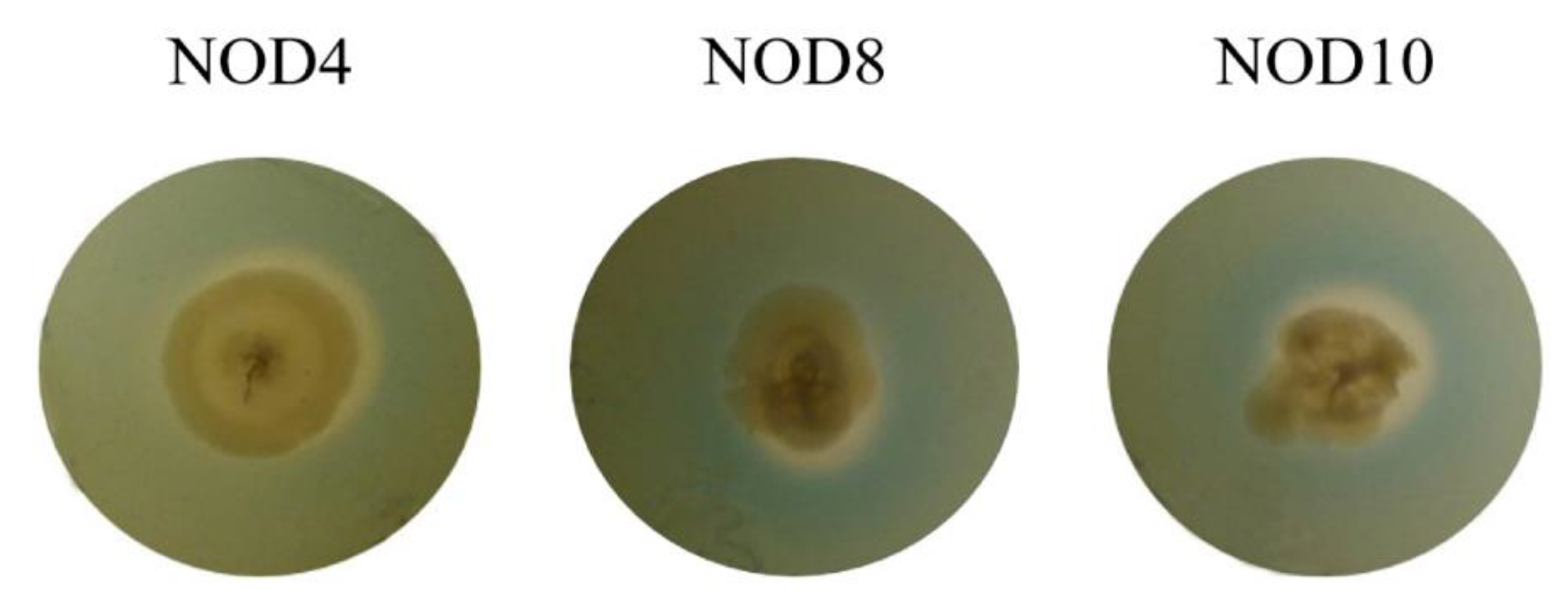
2.5. Siderophore Production
2.6. In Vitro Evaluation of Enterobacter Strains against P. redivivus and N. aberrans
2.7. In Vitro Evaluation of Enterobacter against the Mite T. putrescentiae
2.8. Bioinformatic Analysis of Genomes for the Detection of Genes Involved in Plant Growth-Promoting Activities and Biocontrol
3. Discussion
4. Materials and Methods
4.1. Isolation of Microorganisms from Mimosa Pudica Nodules
4.2. DNA Extraction, Library Preparation, and Sequencing
4.3. Phylogenetics and Phylogenomics
4.4. Phosphate Solubilization
4.5. Determination of Auxin Production
4.6. Siderophore Production Assays
4.7. Panagrellus Redivivus
4.8. Nacobbus Aberrans
4.9. Tyrophagus Putrescentiae
4.10. Experimental Design
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manzanilla-López, R.H.; Costilla, M.A.; Doucet, M.; Inserra, R.N.; Lehman, P.S.; Cid del Prado-Vera, L.; Souza, R.; Evans, K. The genus Nacobbus thorne & Allen, 1944 (Nematoda: Pratylenchidae): Systematics, distribution, biology and management. Nematropica 2002, 32, 149–227. [Google Scholar]
- Valadez, R.R.; Siller, M.C.; Chávez, E.C.; Fuentes, Y.M.O.; Pérez, A.H.; Vargas, L.M.T. Nematodos asociados al cultivo de berenjena en Cañada Honda, México. REMEXCA 2021, 12, 1304–1312. [Google Scholar]
- Erban, T.; Klimov, P.B.; Smrz, J.; Phillips, T.W.; Nesvorna, M.; Kopecky, J.; Hubert, J. Populations of Stored Product Mite Tyrophagus putrescentiae Differ in Their Bacterial Communities. Front. Microbiol. 2016, 7, 1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taghavi, S.; Van Der Lelie, D.; Hoffman, A.; Zhang, Y.B.; Walla, M.D.; Vangronsveld, J.; Monchy, S. Genome sequence of the plant growth promoting endophytic bacterium Enterobacter sp. 638. PLoS Genet. 2010, 6, e1000943. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Charles, T.C.; Glick, B.R. Delay of flower senescence by bacterial endophytes expressing 1-aminocyclopropane-1-carboxylate deaminase. J. Appl. Microbiol. 2012, 113, 1139–1144. [Google Scholar] [CrossRef]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, C.M.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Barrao, C.A.; Lafi, F.F.; Alam, I.; De Zélicourt, A.; Eida, A.A.; Bokhari, A.; Saad, M.M. Complete genome sequence analysis of Enterobacter sp. SA187, a plant multi-stress tolerance promoting endophytic bacterium. Front. Microbiol. 2017, 8, 2023. [Google Scholar] [CrossRef] [Green Version]
- Ludueña, L.M.; Anzuay, M.S.; Angelini, J.G.; McIntosh, M.; Becker, A.; Rupp, O.; Goesmann, A.; Blom, J.; Fabra, A.; Taurian, T. Genome sequence of the endophytic strain Enterobacter sp. J49, a potential biofertilizer for peanut and maize. Genomics 2019, 111, 913–920. [Google Scholar] [CrossRef]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased drought stress resilience of maize through endophytic colonization by Burkholderia phytofirmans PsJN and Enterobacter sp. FD17. Environ. Exp. Bot. 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Ali, S.; Duan, J.; Charles, T.C.; Glick, B.R. A bioinformatics approach to the determination of genes involved in endophytic behavior in Burkholderia spp. J. Theor. Biol. 2014, 343, 193–198. [Google Scholar] [CrossRef]
- Yaish, M.W. Draft genome sequence of endophytic bacterium Enterobacter asburiae PDA134, isolated from date palm (Phoenix dactylifera L.) roots. Genome Announc. 2016, 4, e00848-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinton, D.M.; Bacon, C.W. Enterobacter cloacae is an endophytic symbiont of corn. Mycopathologia 1995, 129, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Araújo, W.L.; Marcon, J.; Maccheroni, W.J.; van Elsas, D.; van Vuurde, J.W.L. Diversity of endophytic bacterial populations and their interaction with Xylella fastidiosa in citrus plants. Appl. Environ. Microbiol. 2002, 68, 4906–4914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asis, C.A.; Adachi, K. Isolation of endophytic diazotroph Pantoea agglomerans and nondiazotroph Enterobacter asburiae from sweet potato stem in Japan. Lett. Appl. Microbiol. 2003, 38, 19–23. [Google Scholar] [CrossRef]
- Anzuay, M.S.; Ruiz Ciancio, M.G.; Ludueña, L.M.; Angelini, J.G.; Barros, G.; Pastor, N.; Taurian, T. Growth promotion of peanut (Arachis hypogaea L.) and maize (Zea mays L.) plants by single and mixed cultures of efficient phosphate solubilizing bacteria that are tolerant to abiotic stress and pesticides. Microbiol. Res. 2017, 199, 98–109. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.K.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Maiti, T.K. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res Microbial. 2018, 169, 20–32. [Google Scholar] [CrossRef]
- Zhao, Y.; Yuan, Z.; Wang, S.; Wang, H.; Chao, Y.; Sederoff, R.R.; Sederoff, H.; Yan, H.; Pan, J.; Peng, M.; et al. Gene sdaB Is Involved in the Nematocidal Activity of Enterobacter ludwigii AA4 Against the Pine Wood Nematode Bursaphelenchus xylophilus. Front. Microbiol. 2022, l13, 870519. [Google Scholar] [CrossRef]
- Oh, M.; Han, J.W.; Lee, C.; Choi, G.J.; Kim, H. Nematicidal and Plant Growth-Promoting Activity of Enterobacter asburiae HK169: Genome Analysis Provides Insight into Its Biological Activities. J. Microbiol. Biotechnol. 2018, 28, 968–975. [Google Scholar] [CrossRef] [Green Version]
- Konstantinidis, K.T.; Tiedje, J.M. Genomic insights that advance the species definition for prokaryotes. Proc. Natl. Acad. Sci. USA 2005, 102, 2567–2572. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Ciufo, S.; Kannan, S.; Sharma, S.; Badretdin, A.; Clark, K.; Turner, S.; Brover, S.; Schoch, C.L.; Kimchi, A.; DiCuccio, M. Using average nucleotide identity to improve taxonomic assignments in prokaryotic genomes at the NCBI. Int. J. Syst. Evol. Microbiol. 2018, 68, 2386–2392. [Google Scholar] [CrossRef]
- Hall, S.A.; Mack, K.; Blackwell, A.; Evans, K.A. Identification and disruption of bacteria associated with sheep scab mites-novel means of control? Exp. Parasitol. 2015, 157, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Saccà, M.L.; Lodesani, M. Isolation of bacterial microbiota associated to honey bees and evaluation of potential biocontrol agents of Varroa destructor. Benef. Microbes. 2020, 15, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Cordova-Kreylos, A.L.; Fernandez, L.E.; Koivunen, M.; Yang, A.; Flor-Weiler, L.; Marrone, P.G. Isolation and characterization of Burkholderia rinojensis sp. nov., a non-Burkholderia cepacia complex soil bacterium with insecticidal and miticidal activities. Appl. Environ. Microbiol. 2013, 79, 7669–7678. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Y.; Wang, Y.H.; Yang, J.; Cui, W.Y.; He, P.J.; Munir, S.; He, P.F.; Wu, Y.X.; He, Y.Q. Acaricidal Activity of Cyclodipeptides from Bacillus amyloliquefaciens W1 against Tetranychus urticae. J. Agric. Food Chem. 2018, 66, 10163–10168. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, W.S.; Akhkha, A.; El-Naggar, M.Y.; Elbadry, M. In vitro antagonistic activity, plant growth promoting traits and phylogenetic affiliation of rhizobacteria associated with wild plants grown in arid soil. Front. Microbiol. 2014, 5, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, D.J.; Singh, R.K.; Singh, P.; Li, D.P.; Sharma, A.; Xing, Y.X.; Song, X.P.; Yang, L.T.; Li, Y.R. Complete Genome Sequence of Enterobacter roggenkampii ED5, a Nitrogen Fixing Plant Growth Promoting Endophytic Bacterium with Biocontrol and Stress Tolerance Properties, Isolated from Sugarcane Root. Front. Microbiol. 2020, 11, 580081. [Google Scholar] [CrossRef]
- Solanki, M.K.; Singh, R.K.; Srivastava, S.; Kumar, S.; Kashyap, P.L.; Srivastava, A.K.; Arora, D.K. Isolation and characterization of siderophore producing antagonistic rhizobacteria against Rhizoctonia solani. J. Basic. Microbiol. 2014, 54, 585–597. [Google Scholar] [CrossRef]
- Sánchez-Cruz, R.; Tapia Vázquez, I.; Batista-García, R.A.; Méndez-Santiago, E.W.; Sánchez-Carbente, M.D.R.; Leija, A.; Lira-Ruan, V.; Hernández, G.; Wong-Villarreal, A.; Folch-Mallol, J.L. Isolation and characterization of endophytes from nodules of Mimosa pudica with biotechnological potential. Microbiol. Res. 2019, 218, 76–86. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinuesa, P.; Ochoa-Sánchez, L.E.; Contreras-Moreira, B. GET_PHYLOMARKERS, a Software Package to Select Optimal Orthologous Clusters for Phylogenomics and Inferring Pan-Genome Phylogenies, Used for a Critical Geno-Taxonomic Revision of the Genus Stenotrophomonas. Front. Microbiol. 2018, 9, 771. [Google Scholar] [CrossRef] [PubMed]
- Caballero, M.J.; Onofre, L.J.; Estrada De Los, S.P.; Martínez, A.L. The tomato rhizosphere, an environment rich in nitrogen-fixing Burkholderia species with capabilities of interest for agriculture and bioremediation. Appl. Environ. Microbiol. 2007, 73, 5308–5319. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- De Lara, R.; Castro, T.; Castro, J.; Castro, G. Nematode culture of Panagrellus redivivus (Goodey, 1945) with Spirulina sp., enriched oatmeal. Rev. Biol. Mar Oceanogr. 2007, 42, 29–36. [Google Scholar] [CrossRef]
- Quintero-Elena, Z.J.; Aguilar-Marcelino, L.; Castañeeda-Ramírez, G.S.; Gómez-Rodríguez, O.; Villar-Luna, E.; López-Guillén, G.; Ramírez-Rojas, S.G. In vitro and micro-plot predatory activity of the mite Caloglyphus mycophagus against populations of nematode larvae of agricultural importance. Biol. Control 2022, 165, 104813. [Google Scholar] [CrossRef]
- Vrain, T.C. A technique for the collection of larvae of Meloidogyne spp., and a comparison of eggs and larvae as inocula. J. Nematol. 1977, 9, 249–251. [Google Scholar]
- Villar, L.E.; Reyes, T.B.; Rojas, M.R.; Gómez, R.O.; Hernández, A.A.; Zavaleta, M.E. Respuesta hipersensitiva en el follaje de chile CM.334 resistente a Phytophthora capsici infectado con Nacobbus aberrans. Nematropica 2009, 39, 143–155. [Google Scholar]
- García Ortiz, N.; Aguilar Marcelino, L.; Mendoza de Gives, P.; López Arellano, M.E.; Bautista Garfias, C.R.; González Garduño, R. In vitro predatory activity of Lasioseius penicilliger (Arachnida: Mesostigmata) against three nematode species: Teladorsagia circumcincta, Meloidogyne sp. and Caenorhabditis elegans. Vet. México OA 2015, 2, 01–09. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | Enterobacter sp. NOD4 | Enterobacter sp. NOD8 | Enterobacter sp. NOD10 |
|---|---|---|---|
| Contigs | 242 | 244 | 144 |
| Genome size | 4,649,192 bp | 4,835,007 bp | 4,517,298 bp |
| GC content (%) | 53.1 | 55.7 | 56.1 |
| Coding gene | 4378 | 4403 | 4198 |
| tRNA | 77 | 80 | 80 |
| rRNA | 17 | 19 | 12 |
| tmRNA | 1 | 1 | 1 |
| Hypothetical proteins | 1064 | 958 | 965 |
| Proteins with functional assignments | 3314 | 3445 | 3233 |
| Treatment | Concentration | %Mortality |
|---|---|---|
| Enterobacter sp. NOD 4 | 1 × 109 cell/mL | 81.2 b ± 11.08 |
| Enterobacter mori NOD8 | 1 × 109 cell/mL | 72.4 c ± 10.23 |
| Enterobacter asburiae NOD10 | 1 × 109 cell/mL | 64.8 b ± 27.33 |
| Control (Water) | - | 2.1 d ± 3.35 |
| Ivermectin | 2.5 mg/mL | 99.3 a ± 4.15 |
| Treatment | Concentration | % Mortality |
|---|---|---|
| Enterobacter sp. NOD4 | 1 × 109 cell/mL | 70.1 b ± 9.2 |
| Enterobacter mori NOD8 | 1 × 109 cell/mL | 62.5 b ± 10.2 |
| Enterobacter asburiae NOD10 | 1 × 109 cell/mL | 58.7 b ± 19.4 |
| Control (Water) | - | 2.1 d ± 3.35 |
| Nematrol | 6.0 mg/mL | 99.3 a ± 4.15 |
| Treatment | Concentration | % Mortality |
|---|---|---|
| Enterobacter sp. NOD 4 | 1 × 109 cell/mL | 68.2 b ± 7.31 |
| Enterobacter mori NOD 8 | 1 × 109 cell/mL | 64.3 b ± 10.1 |
| Enterobacter asburiae NOD 10 | 1 × 109 cell/mL | 77.8 b ± 15.2 |
| Control (Water) | - | 2.1 d ± 3.35 |
| Ivermectin | 2.5 mg/mL | 99.3 a ± 4.15 |
| Gene | Gene Locus NOD4 JAKKOK000000000.1 Enterobacter sp. | Gene Locus NOD8 JAKNRT000000000.1 Enterobacter mori | Gene Locus NOD10 JAKKOL000000000.1 Enterobacter asburiae | Gene Product |
|---|---|---|---|---|
| pstS | L2X67_10965 36964.38004 | L2X83_10375 11318.12358 | L2X78_07895 87736.88776 | Phosphate-binding protein |
| phoU | L2X67_10945 33473.34198 | L2X78_07875 84186.84911 | L2X83_10395 15184.15909 | Negative regulatory protein of pho regulon |
| aspC | L2X83_17980 1562.2752 | Catalyzes the conversion of indole-3-pyruvic acid to indole-3-acetaldehyde | ||
| nifJ | L2X67_22275 1..2275 | L2X78_22070 111.3635 | L2X83_09580 13515.17039 | Nitrogen fixation protein |
| hscA | L2X67_06355 33733.35583 | L2X78_13325 33980..35830 | L2X83_16170 11435.13285 | Fe-S protein assembly chaperone HscA |
| erpA | L2X67_09295 21597.21941 | L2X78_18585 13739..14086 | L2X83_05375 44667.45014 | Iron–sulfur cluster insertion protein ErpA |
| glnK | L2X67_18100 6179.6517 | L2X78_19110 3818.4156 | L2X83_18150 3818.4156 | P-II family nitrogen regulator |
| narX | L2X67_19200 1906.3702 | L2X78_20400 1883.3679 | L2X83_19625 11800.13596 | Nitrate/nitrite two-component system sensor histidine kinase NarX |
| amtB | L2X67_18095 4854.6143 | L2X78_19105 2496.3782 | L2X83_18145 7443.8732 | Ammonium transporter AmtB |
| nirD | L2X67_10630 32781.33107 | L2X78_00765 171713.172039 | L2X83_04385 140648.140974 | Nitrite reductase small subunit NirD |
| nasR | L2X67_19240 15543.16727 | L2X78_20440 15561.16745 | Nitrate regulatory protein NasR | |
| narI | L2X78_20090 2639.3319 | respiratory nitrate reductase subunit gamma | ||
| aldB | L2X67_04385 31416.32954 | L2X83_21280 5772.7310 | Aldehyde dehydrogenase AldB | |
| entH | L2X67_04135 81428.81841 | L2X78_06195 71878.72291 | L2X83_08015 81225.81638 | Proofreading thioesterase EntH |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sachman-Ruíz, B.; Wong-Villarreal, A.; Aguilar-Marcelino, L.; Lozano-Aguirre, L.F.; Espinosa-Zaragoza, S.; Reyes-Reyes, A.L.; Sanzón-Gómez, D.; Mireles-Arriaga, A.I.; Romero-Tirado, R.; Rocha-Martínez, M.K.; et al. Nematicidal, Acaricidal and Plant Growth-Promoting Activity of Enterobacter Endophytic Strains and Identification of Genes Associated with These Biological Activities in the Genomes. Plants 2022, 11, 3136. https://doi.org/10.3390/plants11223136
Sachman-Ruíz B, Wong-Villarreal A, Aguilar-Marcelino L, Lozano-Aguirre LF, Espinosa-Zaragoza S, Reyes-Reyes AL, Sanzón-Gómez D, Mireles-Arriaga AI, Romero-Tirado R, Rocha-Martínez MK, et al. Nematicidal, Acaricidal and Plant Growth-Promoting Activity of Enterobacter Endophytic Strains and Identification of Genes Associated with These Biological Activities in the Genomes. Plants. 2022; 11(22):3136. https://doi.org/10.3390/plants11223136
Chicago/Turabian StyleSachman-Ruíz, Bernardo, Arnoldo Wong-Villarreal, Liliana Aguilar-Marcelino, Luis Fernando Lozano-Aguirre, Saúl Espinosa-Zaragoza, Ana Laura Reyes-Reyes, Diana Sanzón-Gómez, Ana Isabel Mireles-Arriaga, Rodrigo Romero-Tirado, Marisol Karina Rocha-Martínez, and et al. 2022. "Nematicidal, Acaricidal and Plant Growth-Promoting Activity of Enterobacter Endophytic Strains and Identification of Genes Associated with These Biological Activities in the Genomes" Plants 11, no. 22: 3136. https://doi.org/10.3390/plants11223136