
Green Synthesized of Thymus vulgaris Chitosan Nanoparticles Induce Relative WRKY-Genes Expression in Solanum lycopersicum against Fusarium solani, the Causal Agent of Root Rot Disease

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. GC-MS Composition of T. vulgaris EO
2.2. Characterization of ThE-CsNPs
2.2.1. Transmission Electron Microscope
2.2.2. Fourier Transform Infrared Spectroscopy
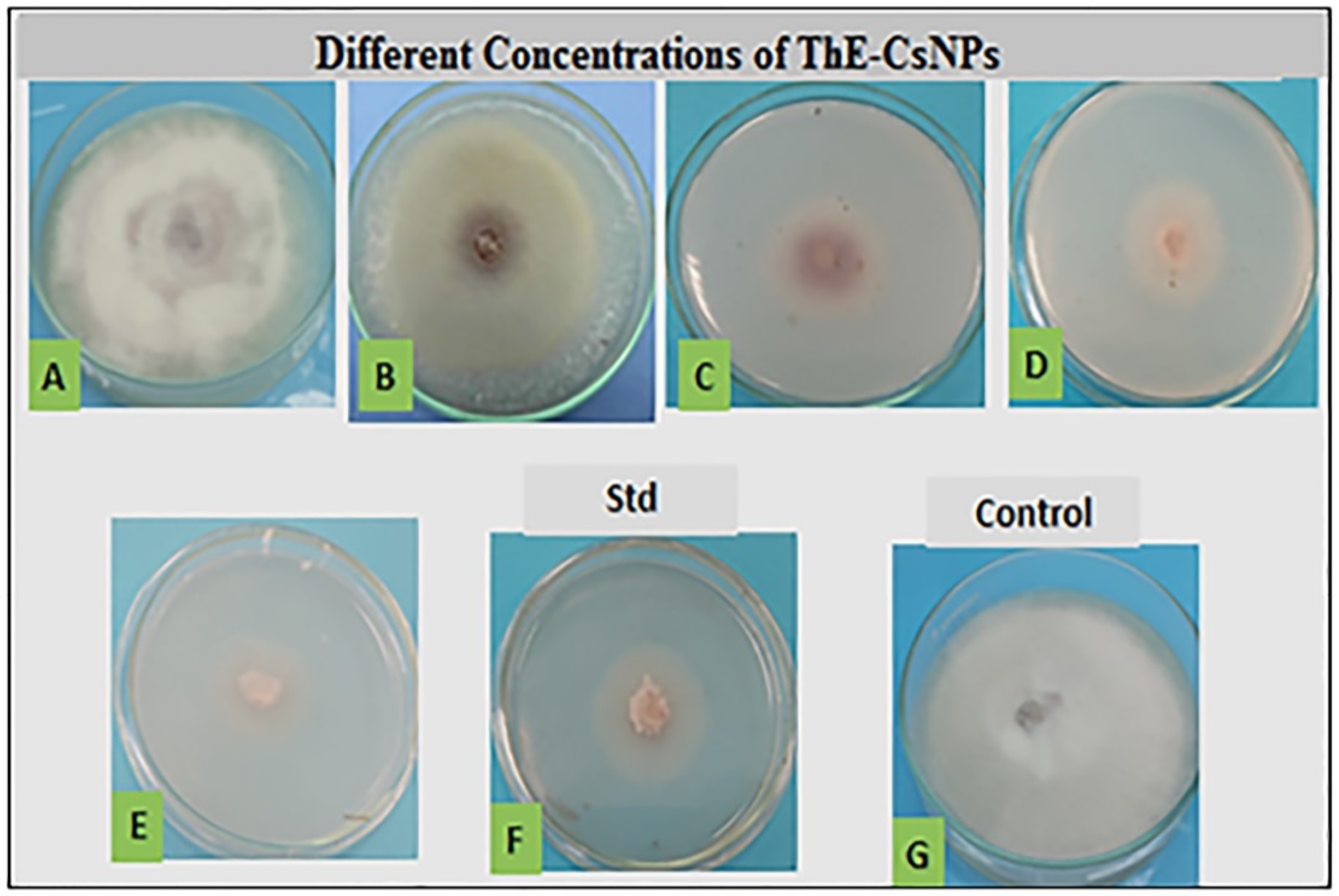
2.3. In Vitro Antifungal Activity
2.4. In Vivo Trial
2.4.1. Pathogenicity Assay
2.4.2. Eco-Physiological Parameters
2.4.3. Physiological and Biochemical Characteristics
2.4.4. Antioxidant Enzymes Activities
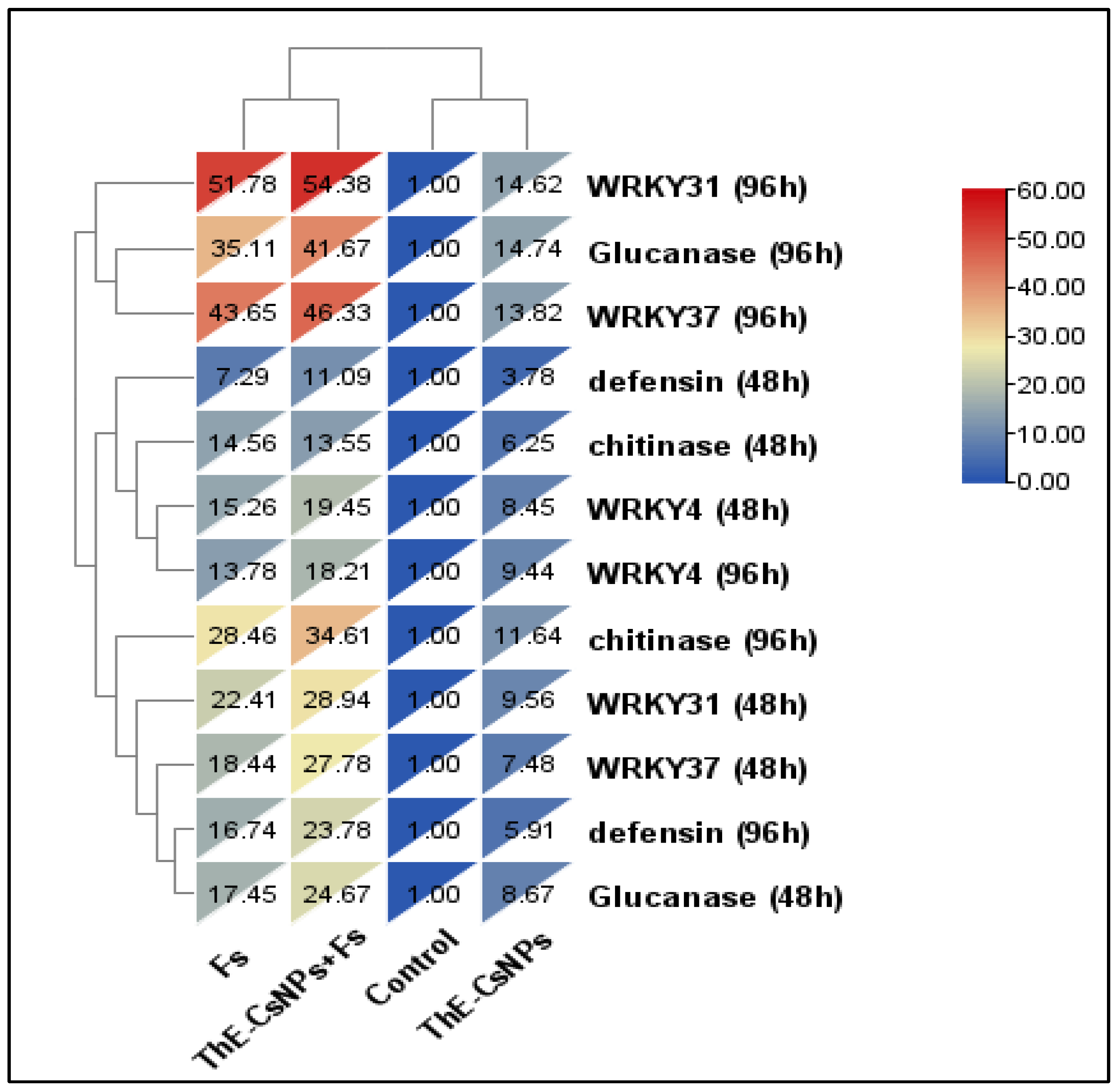
2.5. Molecular Analysis
3. Discussion
4. Materials and Methods
4.1. Sample Collection, Identification, and Preparation
4.2. Medicinal Plant Material
4.2.1. Extractions of Essential Oils
4.2.2. GC-MS of Essential Oils
4.2.3. Preparation of ThE-CsNPs
4.2.4. Fourier Transform Infrared Analysis
4.2.5. Transmission Electron Microscopy
4.3. In Vitro Antifungal Activity of ThE-CsNPs
4.4. In Vivo Greenhouse Trial
4.4.1. Seeds Treatments
4.4.2. Pathogenicity Assay
4.4.3. Pot Trials
4.4.4. Disease Assessments
4.4.5. Eco-Physiological Parameters
4.4.6. Biochemical Assessment
4.4.7. Assay of Antioxidant Enzymes
4.5. Molecular Analysis
4.5.1. RNA Extraction and cDNA Synthesis
4.5.2. Real-Time Quantitative PCR Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bihon, W.; Ognakossan, K.E.; Tignegre, J.-B.; Hanson, P.; Ndiaye, K.; Srinivasan, R. Evaluation of Different Tomato (Solanum lycopersicum L.) Entries and Varieties for Performance and Adaptation in Mali, West Africa. Horticulturae 2022, 8, 579. [Google Scholar] [CrossRef]
- EL-Mansy, A.B.; Abd El-Moneim, D.; ALshamrani, S.M.; Safhi, F.A.; Abdein, M.A.; Ibrahim, A.A. Genetic Diversity Analysis of Tomato (Solanum lycopersicum L.) with Morphological, Cytological, and Molecular Markers under Heat Stress. Horticulturae 2021, 7, 65. [Google Scholar] [CrossRef]
- Fufa, F.; Hanson, P.; Dagnoko, S.; Dhaliwal, M. AVRDC—The world vegetable center tomato breeding in Sub-Saharan Africa:Lessons from the past, present work, and future prospects. Acta Hortic. 2011, 911, 87–98. [Google Scholar] [CrossRef]
- Ayenan, M.A.T.; Danquah, A.; Hanson, P.; Asante, I.K.; Danquah, E.Y. Tomato (Solanum lycopersicum L.) Genotypes Respond Differently to Long-Term Dry and Humid Heat Stress. Horticulturae 2022, 8, 118. [Google Scholar] [CrossRef]
- Soliman, S.A.; Hafez, E.E.; Al-Kolaibe, A.M.G.; Abdel Razik, E.-S.S.; Abd-Ellatif, S.; Ibrahim, A.A.; Kabeil, S.S.A.; Elshafie, H.S. Biochemical Characterization, Antifungal Activity, and Relative Gene Expression of Two Mentha Essential Oils Controlling Fusarium oxysporum, the Causal Agent of Lycopersicon esculentum Root Rot. Plants 2022, 11, 189. [Google Scholar] [CrossRef]
- Bodah, E.T. Root rot diseases in plants: A review of common causal agents and management strategies. Agri. Res. Tech. 2017, 5, 555661. [Google Scholar]
- Srinivas, C.; Devi, D.N.; Murthy, K.N.; Mohan, C.D.; Lakshmeesha, T.R.; Singh, B.; Srivastava, R.K.; Niranjana, S.R.; Hashem, A. Fusarium oxysporum f. sp. lycopersici causal agent of vascular wilt disease of tomato: Biology to diversity—A review. Saudi J. Biol. Sci. 2019, 26, 1315–1324. [Google Scholar]
- Kabaş, A.; Fidan, H.; Batuhan, D.M. Identification of new sources of resistance to resistance-breaking isolates of tomato spotted wilt virus. Saudi J. Biol. Sci. 2021, 28, 3094–3099. [Google Scholar] [CrossRef]
- Arora, H.; Sharma, A.; Poczai, P.; Sharma, S.; Haron, F.F.; Gafur, A.; Sayyed, R.Z. Plant-Derived Protectants in Combating Soil-Borne Fungal Infections in Tomato and Chilli. J. Fungi. 2022, 8, 213. [Google Scholar] [CrossRef]
- Haq, I.U.; Sarwar, M.K.; Faraz, A.; Latif, M.Z. Synthetic chemicals: Major component of plant disease management. In Plant Disease Management Strategies for Sustainable Agriculture through Traditional and Modern Approaches; Haq, I.U., Ijaz, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 53–81. [Google Scholar]
- Aamir, M.; Singh, V.K.; Dubey, M.K.; Kashyap, S.P.; Zehra, A.; Upadhyay, R.S.; Singh, S. Structural and functional dissection of differentially expressed tomato WRKY transcripts in host defense response against the vascular wilt pathogen (Fusarium oxysporum f. sp. lycopersici. PLoS ONE 2018, 13, e0193922. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Shekhar, S.; Rustagi, A.; Sharma, V.; Kumar, D. Insights into the Role of WRKY Superfamily of Protein Transcription Factor in Defense Response. In Molecular Aspects of Plant-Pathogen Interaction; Singh, A., Singh, I., Eds.; Springer: Singapore, 2018; pp. 185–202. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Alkhatib, F.M.; Alzahrani, S.O.; Shafi, M.E.; Abdel-Hamid, S.E.; Taha, T.F.; Aboelenin, S.M.; Soliman, M.M.; Ahmed, N.H. Impact of mycogenic zinc nanoparticles on performance, behavior, immune response, and microbial load in Oreochromis niloticus. Saudi J. Biol. Sci. 2021, 28, 4592–4604. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Desoky, E.S.M.; Saad, A.M.; Eid, R.S.; Selem, E.; Elrys, A.S. Biological silicon nanoparticles improve Phaseolus vulgaris L. yield and minimize its contaminant contents on a heavy metals-contaminated saline soil. J. Environ. Sci. 2021, 106, 1–14. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Saad, A.M.; Najjar, A.A.; Alzahrani, S.O.; Alkhatib, F.M.; Shafi, M.E.; Selem, E.; Desoky, E.S.; Fouda, S.E.E.; El-Tahan, A.M.; et al. The use of biological selenium nanoparticles to suppress Triticum aestivum L. crown and root rot diseases induced by Fusarium species and improve yield under drought and heat stress. Saudi J. Biol. Sci. 2021, 28, 4461–4471. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Saad, A.M.; Taha, T.F.; Najjar, A.A.; Zabermawi, N.M.; Nader, M.M. Selenium nanoparticles from Lactobacillus paracasei HM1 capable of antagonizing animal pathogenic fungi as a new source from human breast milk. Saudi J. Biol. Sci. 2021, 28, 6782–6794. [Google Scholar] [CrossRef]
- Elizabath, A.; Babychan, M.; Mathew, A.M.; Syriac, G.M. Application of nanotechnology in agriculture. Int. J. Pure Appl. Biosci. 2019, 7, 131–139. [Google Scholar] [CrossRef]
- Elieh-Ali-Komi, D.; Hamblin, M.R. Chitin and chitosan: Production and application of versatile biomedical nanomaterials. Int. J. Adv. Res. 2016, 4, 411–427. [Google Scholar]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Shafi, M.E.; Zabermawi, N.M.; Arif, M.; Batiha, G.E.; Khafaga, A.F.; Abd El-Hakim, Y.M.; Al-Sagheer, A.A. Antimicrobial and antioxidant properties of chitosan and its derivatives and their applications: A review. Int. J. Biol. Macromol. 2020, 164, 2726–2744. [Google Scholar] [CrossRef]
- Pichyangkura, R.; Chadchawan, S. Biostimulant activity of chitosan in horticulture. Sci. Hortic. 2015, 196, 49–65. [Google Scholar] [CrossRef]
- Hassan, O.; Chang, T. Chitosan for eco-friendly control of plant disease. Asian J. Plant Pathol. 2017, 11, 53–70. [Google Scholar] [CrossRef] [Green Version]
- Ocaňa, A.; Reglero, G. Effects of Thyme Extract Oils (from Thymus vulgaris, Thymus zygis, and Thymus hyemalis) on Cytokine Production and Gene Expression of oxLDL-Stimulated THP-1-Macrophages. J. Obes. 2012, 2012, 104706. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, L.; Nair, C.K. Therapeutic potentials of silver nanoparticle complex of α-lipoic acid. Nanomater. Nanotechnol. 2011, 1, 17–24. [Google Scholar] [CrossRef]
- Tohidi, B.; Rahimmalek, M.; Arzani, A. Essential oil composition, total phenolic, flavonoid contents, and antioxidant activity of Thymus species collected from different regions of Iran. Food Chem. 2017, 220, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Loo, E.P.; Yasuda, S. Pattern recognition receptors and signaling in plant-microbe interactions. Plant J. 2018, 93, 592–613. [Google Scholar] [CrossRef] [PubMed]
- Vives-Peris, V.; Marmaneu, D.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Characterization of Citrus WRKY transcription factors and their responses to phytohormones and abiotic stresses. Biol. Plant 2018, 62, 33–44. [Google Scholar] [CrossRef]
- Shankar, A.; Pandey, A.; Pandey, G.K. WRKY transcription factor: Role in abiotic and biotic stress. Plant Stress 2013, 7, 26–34. [Google Scholar]
- Bai, Y.; Sunarti, S.; Kissoudis, C.; Visser, R.G.F.; van der Linden, C.G. The role of tomato WRKY genes in plant responses to combined abiotic and biotic stresses. Front. Plant Sci. 2018, 9, 801. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Gao, Y.; Liu, J.; Peng, X.; Niu, X.; Fei, Z. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum. Mol. Genet. Genomics 2012, 287, 495–513. [Google Scholar] [CrossRef]
- Karkute, S.G.; Gujjar, R.S.; Rai, A.; Akhtar, M.; Singh, M. Genome wide expression analysis of WRKY genes in tomato (Solanum lycopersicum) under drought stress. Plant Gene 2018, 13, 8–17. [Google Scholar] [CrossRef]
- Chi, Y.; Yang, Y.; Zhou, Y.; Zhou, J.; Fan, B.; Yu, J.Q. Protein—Protein interactions in the regulation of WRKY transcription factors. Mol. Plant 2013, 6, 287–300. [Google Scholar] [CrossRef] [Green Version]
- Shenashen, M.; Derbalah, A.; Hamza, A.; Mohamed, A.; El Safty, S. Recent trend in controlling root rot disease of tomato caused by Fusarium Solani using aluminasilica nanoparticles. Int. J. Adv. Res. Biol. Sci. 2017, 4, 105–119. [Google Scholar]
- Wani, A.H.; Shah, M.A. A unique and profound effect of MgO and ZnO nanoparticles on some plant pathogenic fungi. J. Appl. Pharm. Sci. 2012, 2, 40–44. [Google Scholar]
- Ozbay, N.; Newman, S.E. Fusarium crown and root rot of tomato and control methods. Plant Pathol. J. 2004, 3, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Cosentino, C.; Labella, C.; Elshafie, H.S.; Camele, I.; Musto, M.; Paolino, R.; Freschi, P. Effects of different heat treatments on lysozyme quantity and antimicrobial activity of jenny milk. J. Dairy Sci. 2016, 99, 5173–5179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshafie, H.S.; Viggiani, L.; Mostafa, M.S.; El-Hashash, M.A.; Bufo, S.A.; Camele, I. Biological activity and chemical identification of ornithine lipid produced by Burkholderia gladioli pv. agaricicola ICMP 11096 using LC-MS and NMR analyses. J. Biol. Res. 2017, 90, 96–103. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Devescovi, G.; Venturi, V.; Camele, I.; Bufo, S.A. Study of the regulatory role of N-acyl homoserine lactones mediated quorum sensing in the biological activity of Burkholderia gladioli pv. agaricicola causing soft rot of Agaricus spp. Front. Microbiol. 2019, 10, 2695. [Google Scholar] [CrossRef]
- Sofo, A.; Elshafie, H.S.; Scopa, A.; Mang, S.M.; Camele, I. Impact of airborne zinc pollution on the antimicrobial activity of olive oil and the microbial metabolic profiles of Zn-contaminated soils in an Italian olive orchard. J. Trace Elem. Med. Biol. 2018, 49, 276–284. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Racioppi, R.; Bufo, S.A.; Camele, I. In vitro study of biological activity of four strains of Burkholderia gladioli pv. agaricicola and identification of their bioactive metabolites using GC–MS. Saudi J. Biol. Sci. 2017, 24, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Al-Harbi, N.A.; Al Attar, N.M.; Hikal, D.M.; Mohamed, S.E.; Abdel Latef, A.A.H.; Ibrahim, A.A.; Abdein, M.A. Evaluation of Insecticidal Effects of Plants Essential Oils Extracted from Basil, Black Seeds and Lavender against Sitophilus oryzae. Plants 2021, 10, 829. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Caputo, L.; De Martino, L.; Gruľová, D.; Zheljazkov, V.D.; De Feo, V.; Camele, I. Biological investigations of essential oils extracted from three Juniperus species and evaluation of their antimicrobial, antioxidant and cytotoxic activities. J. Appl. Microbiol. 2020, 129, 1261–1271. [Google Scholar] [CrossRef]
- Camele, I.; Grul’ová, D.; Elshafie, H.S. Chemical composition and antimicrobial properties of Mentha piperita cv. ‘Kristinka’ essential oil. Plants 2021, 10, 1567. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.M.; Elgat, W.; El-Hefny, M.; Salem, M.Z.M.; Taha, A.S.; Al Farraj, D.A.; Elshikh, M.S.; Hatamleh, A.A.; Abdel-Salam, E.M. New Approach for Using of Mentha longifolia L. and Citrus reticulata L. Essential Oils as Wood-Biofungicides: GC-MS, SEM, and MNDO Quantum Chemical Studies. Materials 2021, 14, 1361. [Google Scholar] [CrossRef] [PubMed]
- Werrie, P.Y.; Durenne, B.; Delaplace, P.; Fauconnier, M.L. Phytotoxicity of Essential Oils: Opportunities and Constraints for the Development of Biopesticides. A Review. Foods 2020, 9, 1291. [Google Scholar] [CrossRef]
- Hjort, K.; Bergstrom, M.; Adesina, M.F.; Jansson, J.K.; Smalla, K.; Sjoling, S. Chitinase genes revealed and compared in bacterial isolates, DNA extracts and a metagenomic library from a phytopathogen-suppressive soil. FEMS Microbiol. Ecol. 2010, 71, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, A.A.; Hubbard, J.C.; Liu, L.; Davis, R.M.; Subbarao, K.V. Effects of chitin and chitosan on the incidence and severity of Fusarium yellows in celery. Plant Dis. 1998, 82, 322–328. [Google Scholar] [CrossRef]
- Hosseinnejad, M.; Jafari, S.M. Evaluation of different factors affecting antimicrobial properties of chitosan. Int. J. Biol. Macromol. 2016, 85, 467–475. [Google Scholar] [CrossRef]
- Cretoiu, M.S.; Korthals, G.W.; Visser, J.H.M.; van Elsas, J.D. Chitin amendment increases soil suppressiveness toward plant pathogens and modulates the actinobacterial and oxalobacteraceal communities in an experimental agricultural field. Appl. Environ. Microbiol. 2013, 79, 5291–5301. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Tripathi, A. Effects of Citrus sinensis (L.) Osbeck epicarp essential oil on growth and morphogenesis of Aspergillus niger Van Tieghem. Microbiol. Res. 2008, 163, 337–344. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Ghanney, N.; Mang, S.M.; Ferchichi, A.; Camele, I. An in vitro attempt for controlling severe phytopathogens and human pathogens using essential oils from Mediterranean plants of genus Schinus. J. Med. Food 2016, 19, 266–273. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Sakr, S.; Mang, S.M.; De Feo, V.; Camele, I. Antimicrobial activity and chemical composition of three essential oils extracted from Mediterranean aromatic plants. J. Med. Food 2016, 19, 1096–1103. [Google Scholar] [CrossRef]
- Ben-Jabeur, M.; Ghabri, E.; Myriam, M.; Hamada, W. Thyme essential oil as a defense inducer of tomato against gray mold and Fusarium wilt. Plant Physiol. Biochem. 2015, 94, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmari, A.K.; Athar, M.T.; Al-Faraidy, A.A.; Almuhaiza, M.S. Chemical Composition of Essential Oil of Thymus vulgaris Collected from Saudi Arabian Market. Asian Pac. J. Trop. Biomed. 2017, 7, 147–150. [Google Scholar] [CrossRef]
- Micucci, M.; Protti, M.; Aldini, R.; Frosini, M.; Corazza, I.; Marzetti, C.; Mattioli, L.B.; Tocci, G.; Chiarini, A.; Mercolini, L.; et al. Thymus vulgaris L. Essential Oil Solid Formulation: Chemical Profile and Spasmolytic and Antimicrobial Effects. Biomolecules 2020, 10, 860. [Google Scholar] [CrossRef] [PubMed]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igoe, R.S.; Hui, Y.H. Substances for use in foods: Listing under Title 21 of the Code of Federal Regulations. In Dictionary of Food Ingredients; Igoe, R.S., Hui, Y.H., Eds.; Springer: Boston, MA, USA, 1996; pp. 159–185. ISBN 978-1-4615-6838-4. [Google Scholar]
- Casarin, L.S.; Casarin, F.D.O.; Brandelli, A.; Novello, J.; Ferreira, S.O.; Tondo, E.C. Influence of Free Energy on the Attachment of Salmonella enteritidis and Listeria Monocytogenes on Stainless Steels AISI 304 and AISI 316. LWT-Food Sci. Technol. 2016, 69, 131–138. [Google Scholar] [CrossRef]
- Kostaki, M.; Chorianopoulos, N.; Braxou, E.; Nychas, G.-J.; Giaouris, E. Differential Biofilm Formation and Chemical Disinfection Resistance of Sessile Cells of Listeria Monocytogenes Strains under Monospecies and Dual-Species (with Salmonella Enterica) Conditions. Appl. Environ. Microbiol. 2012, 78, 2586–2595. [Google Scholar] [CrossRef] [Green Version]
- Nguefack, J.; Tamgue, O.; Dongmo, J.L.; Dakole, C.; Leth, V.; Vismer, H.; Zollo, P.A.; Nkengfack, A. Synergistic action between fractions of essential oils from Cymbopogon citratus, Ocimum gratissimum and Thymus vulgaris against Penicillium expansum. Food Control. 2012, 23, 377–383. [Google Scholar] [CrossRef]
- Arnal-Schnebelen, B.; Hadji-Minaglou, F.; Peroteau, J.F.; Ribeyre, F.; de Billerbeck, V.G. Essential oils in infectious gynaecological disease: A statistical study of 658 cases. Int. J. Aromatherapy. 2004, 14, 192–197. [Google Scholar] [CrossRef]
- Isham, C.R.; Bossou, A.R.; Negron, V.; Fisher, K.E.; Kumar, R.; Marlow, L.; Bible, K.C. Pazopanib enhances paclitaxel-induced mitotic catastrophe in anaplastic thyroid cancer. Sci. Transl. Med. 2013, 5, 166ra3. [Google Scholar] [CrossRef] [Green Version]
- Phukan, U.J.; Jeena, G.S.; Shukla, R.K. WRKY transcription factors: Molecular regulation and stress responses in plants. Front. Plant Sci. 2016, 7, 760. [Google Scholar] [CrossRef] [Green Version]
- Després, C.; Chubak, C.; Rochon, A.; Clark, R.; Bethune, T.; Desveaux, D.; Fobert, P. The Arabidopsis NPR1 disease resistance protein is a novel cofactor that confers redox regulation of DNA binding activity to the basic domain/leucine zipper transcription factor TGA1. Plant Cell 2003, 15, 2181–2191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.Y.; Lee, J.H.; Yoo, J.H.; Moon, B.C.; Choi, M.S.; Kang, Y.H.; Lee, S.M.; Kim, H.S.; Kang, K.Y.; Chung, W.S. WRKY group IId transcription factors interact with calmodulin. FEBS Lett. 2005, 579, 1545–1550. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, C.; Fan, B.; Chen, Z. Physical and Functional Interactions between Pathogen-Induced Arabidopsis WRKY18, WRKY40, and WRKY60 Transcription Factors. Plant Cell 2006, 18, 1310–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parnes, C.A. Efficacy of sodium hypochlorite bleach and “alternative” products in preventing transfer of bacteria to and from inanimate surfaces. Environ. Health 1997, 59, 14–20. [Google Scholar]
- Périno, S.; Chemat-Djenni, Z.; Petitcolas, E.; Giniès, C.; Chemat, F. Downscaling of industrial turbo-distillation to laboratory turbo-clevenger for extraction of essential oils. Application of concepts of green analytical chemistry. Molecules 2019, 24, 2734. [Google Scholar] [CrossRef] [Green Version]
- Elshafie, H.S.; Sakr, S.H.; Sadeek, S.A.; Camele, I. Biological investigations and spectroscopic studies of new Moxifloxacin/Glycine-Metal complexes. Chem. Biodiver. 2019, 16, e1800633. [Google Scholar] [CrossRef]
- Filion, M.; St-Arnaud, M.; Jabaji-Hare, S.H. Quantification of F. solani f. sp. phaseoli in mycorrhizal bean plants and surrounding mycorrhizosphere soil using realtime polymerase chain reaction and direct isolations on selective media. Phytopathology 2003, 93, 229–235. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total phenol analysis: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Chavan, J.J.; Gaikwad, N.B.; Kshirsagar, P. Total phenolics, flavonoids and antioxidant properties of three Ceropegia species from Western Ghats of India. S. Afr. J. Bot. 2013, 88, 273–277. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Yang, Y.C.; Sun, D.W.; Pu, H.; Wang, N.N.; Zhu, Z. Rapid detection of anthocyanin content in lychee pericarp during storage using hyperspectral imaging coupled with model fusion. Postharvest. Biol. Technol. 2015, 103, 55–65. [Google Scholar] [CrossRef]
- Luck, H. Catalase in Methods of Enzymatic Analysis; Bergmeyer, J., Grabi, M., Eds.; Academic Press: New York, NY, USA, 1974; Volume II. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quantitative ID | Component Identified | Retention Time (min) | Retention Index (RI) | Area (%) | Identification |
|---|---|---|---|---|---|
| 1 | Cyclohexane | 4.11 | 1215 | 10.12 | RI, MS * |
| 2 | Myrcene | 5.13 | 106 | 7.55 | RI, MS |
| 3 | Caryophyllene | 9.14 | 974 | 3.25 | RI, MS |
| 4 | α-Pinene | 12.39 | 993 | 6.49 | RI, MS |
| 5 | Camphene | 16.81 | 1429 | 35.97 | RI, MS |
| 6 | β-Myrcene | 17.74 | 974 | 2.77 | RI, MS |
| 7 | Carene | 18.56 | 1042 | 2.86 | RI, MS |
| 8 | p-Cymene | 20.35 | 938 | 2.19 | RI, MS |
| 9 | γ-Terpinene | 21.16 | 1062 | 2.68 | RI, MS |
| 10 | α-Terpineol | 21.53 | 1138 | 2.92 | RI, MS |
| 11 | Linalool | 26.13 | 1126 | 2.84 | RI, MS |
| 12 | Thymol | 27.85 | 1074 | 2.31 | RI, MS |
| 13 | α-Thymol | 29.14 | 1261 | 5.17 | RI, MS |
| 14 | Carvacrol | 29.38 | 1062 | 2.21 | RI, MS |
| 15 | Caryophyllen | 32.75 | 1211 | 4.91 | RI, MS |
| 16 | Total | - | - | 94.24 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-Ellatif, S.; Ibrahim, A.A.; Safhi, F.A.; Abdel Razik, E.S.; Kabeil, S.S.A.; Aloufi, S.; Alyamani, A.A.; Basuoni, M.M.; ALshamrani, S.M.; Elshafie, H.S. Green Synthesized of Thymus vulgaris Chitosan Nanoparticles Induce Relative WRKY-Genes Expression in Solanum lycopersicum against Fusarium solani, the Causal Agent of Root Rot Disease. Plants 2022, 11, 3129. https://doi.org/10.3390/plants11223129
Abd-Ellatif S, Ibrahim AA, Safhi FA, Abdel Razik ES, Kabeil SSA, Aloufi S, Alyamani AA, Basuoni MM, ALshamrani SM, Elshafie HS. Green Synthesized of Thymus vulgaris Chitosan Nanoparticles Induce Relative WRKY-Genes Expression in Solanum lycopersicum against Fusarium solani, the Causal Agent of Root Rot Disease. Plants. 2022; 11(22):3129. https://doi.org/10.3390/plants11223129
Chicago/Turabian StyleAbd-Ellatif, Sawsan, Amira A. Ibrahim, Fatmah A. Safhi, Elsayed S. Abdel Razik, Sanaa S. A. Kabeil, Salman Aloufi, Amal A. Alyamani, Mostafa M. Basuoni, Salha Mesfer ALshamrani, and Hazem S. Elshafie. 2022. "Green Synthesized of Thymus vulgaris Chitosan Nanoparticles Induce Relative WRKY-Genes Expression in Solanum lycopersicum against Fusarium solani, the Causal Agent of Root Rot Disease" Plants 11, no. 22: 3129. https://doi.org/10.3390/plants11223129