
Assessing the Cooling and Air Pollution Tolerance among Urban Tree Species in a Tropical Climate
 , , ,
, , ,
Abstract
:1. Introduction
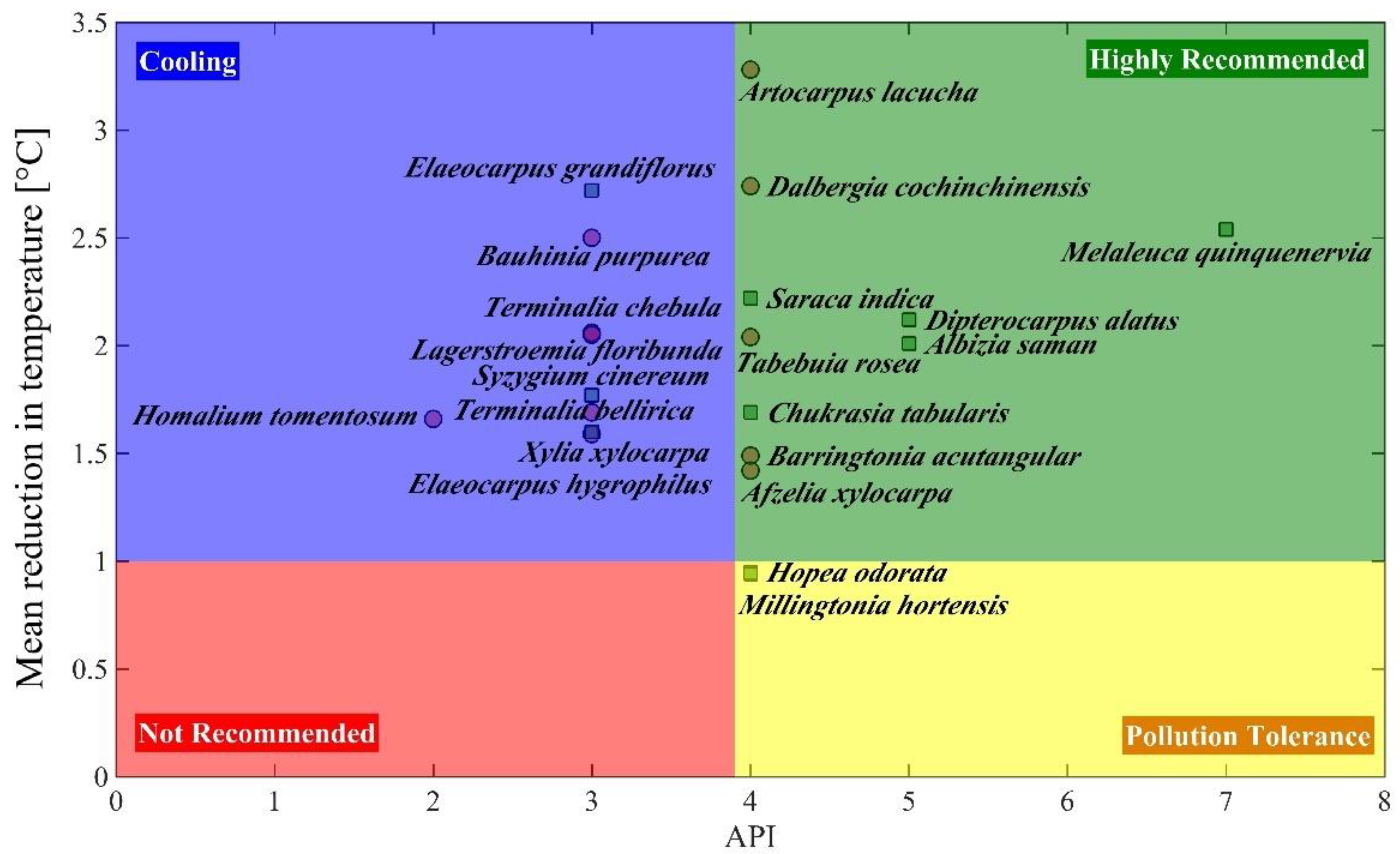
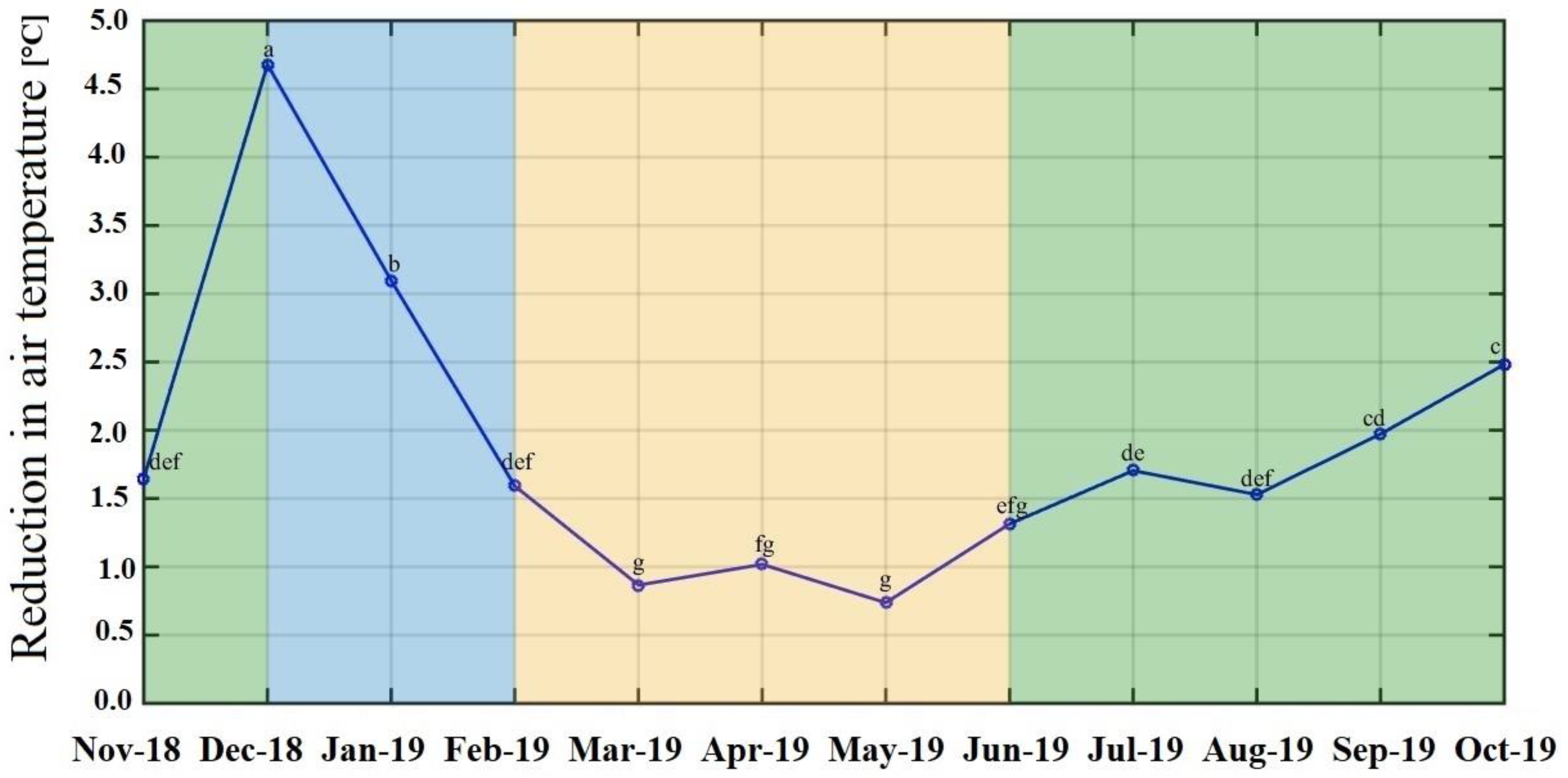
2. Results
3. Discussion
4. Materials and Methods
4.1. Tree Selection
4.2. Calculation of Air Pollution Tolerant Index (APTI) and Anticipated Performance Index (API)
4.3. Calculation of Reduction in Soil Temperature (ΔTs)
0.025 L* − 3.267 vapor pressure (kPa) + 0.682 solar radiation (mW. M−2) −
0.063 wind velocity (Km.h−1)
4.4. Measurement of Reduction in Air Temperature (ΔTa)
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. World Urbanization Prospects 2018. Available online: https://population.un.org/wup (accessed on 28 October 2021).
- Zheng, Z.; Ren, G.; Wang, H.; Dou, J.; Gao, Z.; Guan, C.; Li, Y.; Ngarukiyimana, J.P.; Zhao, C.; Cao, C.; et al. Relationship between fine-particle pollution and the urban heat island in Beijing, Chigna: Observational evidence. Bound.-Layer Meteorol. 2018, 169, 93–113. [Google Scholar] [CrossRef]
- Arifwidodo, S.D.; Tanaka, T. The characteristics of urban heat island in Bangkok, Thailand. ScienceDirect 2015, 195, 423–428. [Google Scholar] [CrossRef]
- Keeratikasikorn, C.; Bonafoni, S. Urban heat island analysis over the land use xoning plan of Bangkok by means of Landst 8 imagery. Remote Sens. 2018, 10, 440. [Google Scholar] [CrossRef]
- Popek, R.; Fornal-Pieniak, B.; Chyliński, F.; Pawełkowicz, M.; Bobrowicz, J.; Chrzanowska, D.; Piechota, N.; Przybysz, A. Not only trees matter-Traffic-Related PM accumulation by vegetation of urban forests. Sustainability 2022, 14, 2973. [Google Scholar] [CrossRef]
- Yarnvudhi, A.; Leksungnoen, N.; Tor-Ngern, P.; Premashthira, A.; Thinkampheang, S.; Hermhuk, S. Evaluation of regulating and provisioning services provided by a park designed to be resilient to climate change in Bangkok, Thailand. Sustainability 2021, 13, 13624. [Google Scholar] [CrossRef]
- Lin, B.-S.; Lin, Y.-J. Cooling effect of shade trees with different characteristics in a subtropical urban park. HortScience 2010, 45, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Rao, D.N.; Agrawal, M.; Pandey, J.; Narayan, D. Air pollution tolerance index of plants. J. Environ. Manag. 1991, 32, 45–55. [Google Scholar] [CrossRef]
- Shannigrahi, A.S.; Fukushima, T.; Sharmaa, R.C. Anticipated air pollutin tolerance of some plant species considered for green belt development in and around an industrial/urban area in India: An overview. Int. J. Environ. Stud. 2004, 2, 125–137. [Google Scholar] [CrossRef]
- Woodward, F.I. Forest air conditioning. Nature 2008, 454, 422–423. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Rudcing Urban Heat Islands: Compendium of Strategies. Available online: https://www.epa.gov/heat-islands/heat-island-compendium. (accessed on 15 December 2021).
- Shiflett, S.A.; Liang, L.L.; Crum, S.M.; Feyisa, G.L.; Wang, J.; Jenerette, G.D. Variation in the urban vegetation, surface temperature, air temperature nexus. Sci. Total Environ. 2017, 579, 495–505. [Google Scholar] [CrossRef]
- Baltzer, J.L.; Grégoire, D.M.; Bunyavejchewin, S.; Noor, N.S.M.; Davies, S.J. Coordination of foliar and wood anatomical traits contributes to tropical tree distributions and productivity along the Malay-Thai Peninsula. Am. J. Bot. 2009, 96, 2214–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaluarachichi, T.U.N.; Tjoelker, M.G.; Pfautsch, S. Temperature reduction in urban surface materials through tree shading depends on surface type not tree species. Forests 2020, 11, 1141. [Google Scholar] [CrossRef]
- Armson, D.; Stringer, P.; Ennos, A.R. The effect of street trees and amenity grass on urban surface water runoff in Manchester, UK. Urban For. Urban Green. 2013, 12, 282–286. [Google Scholar] [CrossRef]
- Oksanen, E.; Kontunen-Soppela, S. Plants have different strategies to defend against air pollutants. Environ. Sci. Health 2021, 19, 100222. [Google Scholar] [CrossRef]
- Przybysz, A.; Popek, R.; Stankiewicz-Kosyl, M.; Zhu, C.Y.; Małecka-Przybysz, M.; Maulidyawati, T.; Mikowska, K.; Deluga, D.; Griżuk, K.; Sokalski-Wieczorek, J.; et al. Where trees cannot grow - Particulate matter accumulation by urban meadows. Sci. Total Environ. 2021, 785, 147310. [Google Scholar] [CrossRef]
- Ter, S.; Chettri, M.K.; Shakya, K. Air pollution tolerance index of some tree species of Pashupati and Budhanilkantha area, Kathmandu. Amrit Res. J. 2020, 1, 20–28. [Google Scholar] [CrossRef]
- Popek, R.; Łukowski, A.; Grabowski, M. Influence of particulate matter accumulation on photosynthetic apparatus of Physocarpus opulifolius and Sorbaria sorbifolia. Pol. J. Environ. Stud. 2018, 27, 2391–2396. [Google Scholar] [CrossRef]
- Smith, W.H. Air Pollution and Forests: Interaction between Air Contaminants and Forest Ecosystems; Springer Inc.: New York, NY, USA, 1990. [Google Scholar]
- Nowak, D.J.; Hirabayashi, S.; Bodine, A.; Hoehn, R. Modeled PM2.5 removal by trees in ten U.S. cities and associated health effects. Environ. Pollut. 2013, 178, 395–402. [Google Scholar] [CrossRef]
- Leghari, S.K.; Zaid, M.A. Effect of air pollutio on the leaf morphology of common plant species of Quetta city. Pak. J. Bot. 2013, 45, 447–454. [Google Scholar]
- Aribal, L.G.; Llamos, E.J.N.; Bruno, A.G.T.; Medina, M.A.P. Comparative leaf morphometrics of two urban tree species: An assessment to air pollution impacts. J. Biodivers. Environ. Sci. 2016, No.1, 106–115. [Google Scholar]
- Pandey, A.K. Air pollution tolerance index and anticipated performance index of some plant species for development of urban forest. Urban For. Urban Green. 2015, 14, 866–871. [Google Scholar] [CrossRef]
- Sumangala, H.P.; Aswath, C.; Laxman, R.H.; Namratha, M.R. Evaluation of air pollution tolerance index (APTI) of selected ornamental tree species of Bengaluru, India. Int. J. Pure Appl. Biosci. 2018, 6, 366–373. [Google Scholar] [CrossRef]
- Khanoranga; Khalid, S. Phytomonitoring of air pollution around brick kilns in Balochistan province Pakistan through air pollution index and metal accumulation index. J. Clean. Prod. 2019, 229. [Google Scholar] [CrossRef]
- Karmakar, D.; Deb, K.; Kumar, P.P. Ecophysiological responses of tree species due to air pollution for biomonitoring of environmental health in urban area. Urban Clim. 2021, 35, 100741. [Google Scholar] [CrossRef]
- Kumar, M.; Nandini, N. Identification and evaluation of air pollution tolerance index of selected avenue tree species of urban Bangalore, India. Int. J. Emerg. Technol. Comput. Appl. Sci. 2013, 5, 1–3. [Google Scholar]
- Kaur, M.; Nagpal, A.K. Evaluation of air pollution tolerance index and anticipated performance index of plants and their application in development of green space along the urban areas. Environ. Sci. Pollut. Res. 2017, 24, 18881–18895. [Google Scholar] [CrossRef]
- Yousafzai, A.; Wongpakapattanawong, P.; Seaengnil, K.; Saipunkaew, W. An assessment of air pollution tolerance capability of common trees in Chiang Mai city, Thailand. Int. J. Adv. Sci. Eng. Technol. 2017, 5, 16–21. [Google Scholar]
- Utarasakul, T. Efficiency of ground cover types under Samanea saman on reducing outdoor thermal environment in urban parks of Bangkok, Thailand. Sci. Direct 2017, 37, 78–82. [Google Scholar] [CrossRef]
- Oliveira, S.; Andrade, H.; Vaz, T. The cooling effect of green spaces as a contribution to the mitigation of urban heat: A case study in Lisbon. Build. Environ. 2011, 46, 2186–2194. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Y.; Ma, K.; Bongers, F.; Sterck, F.J. Fully exposed canopy tree and liana branches in a tropical forest differ in mechanical traits but are similar in hydraulic traits. Tree Physiol. 2019, 39, 1713–1724. [Google Scholar] [CrossRef]
- Prajapati, S.K.; Tripathi, B.D. Anticipated Performance Index of some tree species considered for green belt development in and around an urban area: A caase study of Varanasi city, India. J. Environ. Manag. 2008, 88, 1343–1349. [Google Scholar] [CrossRef] [PubMed]
- Czaja, M.; Kolton, A.; Muras, P. The complex issue of urban trees-stress factor accumulation and ecological service possibilities. Forests 2020, 11, 932. [Google Scholar] [CrossRef]
- Molnár, V.E.; Tóthmérész, B.; Szabó, S.; Simon, E. Pollution assessment in urban areas using air pollution tolerance index of tree species. WIT Trans. Ecol. Environ. 2019. [Google Scholar] [CrossRef] [Green Version]
- Leksungnoen, N.; Uthairatsamee, S.; Andriyas, T. Adaptability of Siamese Rosewood and Teak Seedlings to Varying Light Conditions. Environ. Nat. Resour. J. 2021, 19, 449–458. [Google Scholar] [CrossRef]
- Mahecha, G.S.; Bamniya, B.R.; Nair, N.; Saini, D. Air pollution tolerance index of certain plant species-A study of Madri industrial area, Udaipur (Raj.), India. Int. J. Innov. Res. Sci. Eng. Technol. 2013, 2, 7927–7929. [Google Scholar]
- Rai, P.K.; Panda, L.L.S. Dust capturing potential and air pollution tolerance index (APTI) of some road side tree vegetation in Aizawl, Mizoram, India: An Indo-Burma hot spot region. Air Qual. Atmos. Health 2014, 7, 93–101. [Google Scholar] [CrossRef]
- Das, S.; Prasad, P. Seasonal variation in air pollution tolerance indices and selection of plant species for industrial areas of Rourkela. Indian J. Environ. Prot. 2010, 3. [Google Scholar]
- Kwak, M.J.; Lee, J.K.; Park, S.; Lim, Y.J.; Kim, H.; Kim, K.N.; Je, S.M.; Park, C.R.; Woo, S.Y. Evaluation of the importance of some East Asian tree species for refinement of air quality by estimating air pollution tolerance index, anticipated performance index, and air pollutant uptake. Sustainability 2020, 12, 3067. [Google Scholar] [CrossRef] [Green Version]
- Govindaraju, M.; Ganeshkumar, R.S.; Muthukumaran, V.R.; Visvanatham, P. Identification and evaluation of air-pollution-tolerant plants around lignite-based thermal power station for greenbelt development. Environ. Sci. Pollut. Res. 2011, 19, 1210–1223. [Google Scholar] [CrossRef]
- Michael, B.L.M.; Burke, J.J. Temperature Effects on Root Growth Plant; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Rahman, M.A.; Moser, A.; Gold, A.; Rötzer, T.; Pauleit, S. Vertical air temperature gradients under the shade of two contrasting urban tree species during difference types of summer days. Sci. Total Environ. 2018, 633, 100–111. [Google Scholar] [CrossRef]
- Centenaroa, G.; Hudekb, C.; Zanellaa, A.; Crivellaro, A. Root-soil physical and biotic interactions with a focus on tree root systems: A review. Appl. Soil Ecol. 2018, 123, 318–327. [Google Scholar] [CrossRef]
- Mori, H.; Kamijo, T.; Masaki, T. Liana distribution and community structure in an oldgrowth temperate forest: The relative importance of past disturbances, host trees, and microsite characteristics. Plant Ecol. 2016, 217, 1171–1182. [Google Scholar] [CrossRef]
- Peters, E.B.; McFadden, J.P. Influence of seasonality and vegetation type on suburban microclimates. Urban Ecosyst. 2010, 13, 443–460. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.D.; Dong, L.; Yan, H.; Yang, N.; Xiong, Y. The influence of the spatial characteristics of urban green space on the urban heat island effect in Suzhou industrial park. Sustain. Cities Soc. 2018, 40, 428–439. [Google Scholar] [CrossRef]
- Vidrih, B.; Medved, S. Multiparametric model of urban park cooling island. Urban For. Urban Green. 2013, 12, 220–229. [Google Scholar] [CrossRef]
- McPhearson, T.; Tidball, K.G. Disturbances in Urban Social-Ecological Systems: Niche Opportunities for Environmental Education; Peter Lang: New York, NY, USA, 2001; pp. 1230–1993. [Google Scholar]
- Peguero-Pina, J.J.; Vilagrosa, A.; Alonso-Forn, D.; Ferrio, J.P.; Sancho-Knapik, D.; Gil-Pelegrín, E. Living in drylands: Functional adaptations of trees and shrubs to cope with high temperatures and water scarcity. Forests 2020, 11, 1028. [Google Scholar] [CrossRef]
- Speak, A.; Montagnani, L.; Wellstein, C.; Zerbe, S. The influence of tree traits on urban ground surface shade cooling. Landsc. Urban Plan. 2020, 197, 103748. [Google Scholar] [CrossRef]
- Huber, M.; Nieuwendijk, N.M.; Pantazopoulou, C.K.; Pierik, R. Light signalling shapes plant–plant interactions in dense canopies. Plant Cell Environ. 2021, 44, 1014–1029. [Google Scholar] [CrossRef]
- Chulalongkorn University Centenary Park. Chulalongkorn University Centenary Park a Gift of the Century to Society. Available online: http://www.cu100.chula.ac.th/cu-centenary-park/ (accessed on 23 October 2021).
- Thai Meteorological Department. Weather forecast. Available online: https://www.tmd.go.th/en/ (accessed on 10 March 2020).
- Dat, J.; Vandenabeele, S.; Vranová, E.; Montagu, M.V.; Inzé, D.; Breusegem, F.V. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 2000, 57, 779–795. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Breusegem, F.V. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Miller, G.; SuZuki, N.; ciftci-yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Rathore, D. Oxidative stress defence responses of wheat (Triticum aestivum L.) and chilli (Capsicum annum L.) cultivars grown under textile effluent fertilitzation. Plant Physiol. Biochem. 2017, 123, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, I.J.; Rathore, D. Suspended particulate matter deposition and its impact on urban trees. Atmos. Pollut. Res. 2018, 9, 1072–1082. [Google Scholar] [CrossRef]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2. [Google Scholar] [CrossRef]
- Shahid, M.; Pourrut, B.; Dumat, C.; Nadeem, M.; Aslam, M.; Pinelli, E. Heavy-Metal-Induced Reactive Oxygen Species: Phytotoxicity and Physicochemical Changes in Plants; Springer International Publishing: Cham, Switzerland, 2014; Volume 232. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No. | Species | Forest Type | Habitat | Count (Number of Trees) | LAI | Crown Cover (m2) | DBH (cm) | Total Height (m) | Voucher Specimen Name |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Dalbergia cochinchinensis Pierre | D | Native | 136 | 2.51 ± 5.29 | 4.57 ± 2.21 | 13.82 ± 2.11 | 8.33 ± 1.43 | CU01-2018 |
| 2 | Tabebuia rosea (Bertol.) Bertero ex A. DC. | D | Exotic | 81 | 1.05 ± 0.42 | 2.78 ± 1.40 | 12.71 ± 2.42 | 6.73 ± 0.87 | CU02-2018 |
| 3 | Albizia saman (Jacq.) Merr. | E | Exotic | 74 | 1.32 ± 0.44 | 15.75 ± 7.42 | 19.70 ± 4.89 | 7.71 ± 1.42 | CU03-2018 |
| 4 | Millingtonia hortensis L. f. | E | Native | 70 | 1.36 ± 0.58 | 1.99 ± 0.88 | 14.50 ± 1.78 | 7.97 ± 1.21 | CU04-2018 |
| 5 | Dipterocarpus alatus Roxb. ex G. Don | E | Native | 47 | 0.93 ± 0.49 | 1.62 ± 0.94 | 12.57 ± 2.25 | 8.19 ± 1.38 | CU05-2018 |
| 6 | Afzelia xylocarpa (Kurz) Craib | D | Native | 27 | 5.76 ± 25.17 | 2.37 ± 1.19 | 13.54 ± 2.03 | 7.26 ± 0.83 | CU06-2018 |
| 7 | Hopea odorata Roxb. | E | Native | 26 | 2.15 ± 0.88 | 1.85 ± 1.66 | 13.68 ± 2.14 | 8.70 ± 1.37 | CU07-2018 |
| 8 | Homalium tomentosum (Vent.) Benth. | D | Native | 23 | 1.32 ± 0.44 | 3.03 ± 1.47 | 12.41 ± 1.74 | 8.86 ± 1.56 | CU08-2018 |
| 9 | Lagerstroemia floribunda Jack var. floribunda | D | Native | 22 | 1.72 ± 0.52 | 2.74 ± 1.03 | 12.9 ± 2.35 | 6.45 ± 0.95 | CU09-2018 |
| 10 | Bauhinia purpurea L. | D | Native | 20 | 2.47 ± 1.03 | 5.18 ± 2.05 | 16.28 ± 23.41 | 6.01 ± 1.10 | CU10-2018 |
| 11 | Xylia xylocarpa (Roxb.) W. Theob. var. kerrii (Craib & Hutch.) I. C. Nielsen | D | Native | 17 | 1.71 ± 0.54 | 3.22 ± 1.39 | 13.74 ± 2.33 | 8.47 ± 1.22 | CU11-2018 |
| 12 | Syzygium cinereum (L.) Skeels | E | Native | 12 | 1.55 ± 0.94 | 2.72 ± 1.29 | 16.98 ± 1.99 | 6.45 ± 0.26 | CU12-2018 |
| 13 | Melaleuca quinquenervia (Cav.) S.T. Blake | E | Native | 11 | 1.36 ± 0.39 | 2.23 ± 0.92 | 17.20 ± 2.88 | 8.73 ± 1.60 | CU13-2018 |
| 14 | Chukrasia tabularis A. Juss. | E | Native | 9 | 1.13 ± 0.61 | 2.42 ± 1.06 | 13.49 ± 3.12 | 7.69 ± 0.90 | CU14-2018 |
| 15 | Terminalia chebula Retz. var. chebula | D | Native | 8 | 1.21 ± 0.46 | 1.98 ± 0.92 | 13.44 ± 3.47 | 6.89 ± 1.24 | CU15-2018 |
| 16 | Baringtonia acutangula (L.) Gaertn. | D | Native | 7 | 0.95 ± 0.69 | 2.70 ± 1.74 | 17.01 ± 3.11 | 5.96 ± 0.86 | CU16-2018 |
| 17 | Saraca indica L. | E | Native | 7 | 0.87 0.21 | 2.50 0.53 | 11.77 ± 1.21 | 4.56 ± 0.61 | CU17-2018 |
| 18 | Elaeocarpus hygrophilus Kurz | E | Native | 7 | 1.04 ± 0.51 | 1.84 ± 0.73 | 10.50 ± 2.98 | 5.20 ± 1.05 | CU18-2018 |
| 19 | Terminalia bellirica (Gaertn.) Roxb. | D | Native | 6 | 0.69 ± 0.19 | 3.10 ± 1.21 | 14.48 ± 1.60 | 9.70 ± 0.70 | CU19-2018 |
| 20 | Artocarpus lacucha Roxb. ex Buch.-Ham. | D | Native | 5 | 2.07 ± 0.52 | 2.61 ± 0.65 | 14.08 ± 0.87 | 8.79 ± 1.25 | CU20-2018 |
| 21 | Elaeocarpus grandiflorus Sm. | E | Native | 5 | 1.78 ± 0.92 | 1.99 ± 0.87 | 7.86 ± 2.38 | 3.28 ± 0.62 | CU21-2018 |
| Grading Characteristic | Pattern of Assessment | Grade Allotted | ||
|---|---|---|---|---|
| (a) Tolerance | Air pollution tolerance index (APTI) | 7.0–8.0 | + | |
| 8.1–10.0 | ++ | |||
| 10.1–11.0 | +++ | |||
| 11.1–12.0 | ++++ | |||
| 12.1–13.0 | +++++ | |||
| (b) Biological and socio-economic | (i) Stem size | Small | - | |
| Medium | + | |||
| Large | ++ | |||
| (ii) Canopy structure | Spare/Irregular/Globular | - | ||
| Spreading crown/open/semi-dense | + | |||
| Spreading dense | ++ | |||
| (iii) Leaf phenological type | Deciduous | - | ||
| Evergreen | + | |||
| (iv) Leaf structure | Size (length × width) | Small (2–5 × 1.5–3 cm) | - | |
| Medium (5–15 × 3–8 cm) | + | |||
| Large (8–25 × 5–15 cm) | ++ | |||
| Texture | Smooth (no hair) | - | ||
| Coriaceous (hairy) | + | |||
| Hardiness | Delineate (easy to tear) | - | ||
| Hardy (leathery) | + | |||
| (iv) Economic value | Less than three uses | - | ||
| Three or four uses | + | |||
| Five or more uses | ++ | |||
| After the calculation of APTI, an assessment of the anticipated performance index (API) of plant species (Maximum grade that can be scored by an individual = 16). | ||||
| Grade | Score (%) | Assessment of plant species | ||
| 0 | Up to 30 | Not recommended for planting | ||
| 1 | 31–40 | Very poor | ||
| 2 | 41–50 | Poor | ||
| 3 | 51–60 | Moderate | ||
| 4 | 61–70 | Good | ||
| 5 | 71–80 | Very good | ||
| 6 | 81–90 | Excellent | ||
| 7 | 91–100 | Best | ||
| Pollutants | Study Area | WHO (2021) | Thailand’s Standard | Pollution Level |
|---|---|---|---|---|
| PM2.5 (μg/m3) | 38.01 ± 25.27 | 15 | 25 | High |
| PM10 (μg/m3) | 49.24 ± 32.36 | 45 | 50 | Moderate |
| NO2 (ppb) | 22.47 ± 12.99 | 13 | 60 | Moderate |
| SO2 (ppb) | 2.72 ± 1.36 | 15 | 100 | Low |
| CO (ppm) | 0.90 ± 0.34 | 4 | 4.4 | Low |
| O3 (ppb) | 19.38 ± 14.03 | 51 | 35 | Low |
| No | Plant Species | Total APTI | Assessment Parameters | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| APTI | Tree Habit | Canopy Structure | Type of Tree | Laminar Structure | Economic Importance | Grade Allotted | API Grade | ||||||
| Size | Texture | Hardiness | Total Plus (+) | % Scoring | |||||||||
| 1 | Melaleuca quinquenervia (E) | 12.261 ± 0.06 bc | +++++ | ++ | ++ | + | + | + | + | ++ | 15 | 94 | 7 |
| 2 | Dipterocarpus alatus (E) | 12.130 ± 0.06 bcdce | +++++ | ++ | - | + | ++ | + | + | ++ | 14 | 88 | 6 |
| 3 | Albizia saman (E) | 13.059 ± 0.16 a | +++++ | ++ | ++ | + | - | - | - | ++ | 12 | 75 | 5 |
| 4 | Chukrasia tabularis (E) | 12.191 ± 0.10 bcd | +++++ | ++ | ++ | + | + | - | - | + | 12 | 75 | 5 |
| 5 | Artocarpus lacucha (D) | 11.455 ± 0.06 fghi | ++++ | ++ | ++ | - | + | + | - | ++ | 12 | 75 | 5 |
| 6 | Saraca indica (E) | 11.441 ± 0.12 fghi | ++++ | ++ | ++ | + | + | - | + | + | 12 | 75 | 5 |
| 7 | Baringtonia acutangular (D) | 11.805 ± 0.06 cdef | ++++ | + | ++ | - | + | - | + | ++ | 11 | 69 | 4 |
| 8 | Afzelia xylocarpa (D) | 11.667 ± 0.09 efg | ++++ | ++ | ++ | - | + | - | - | ++ | 11 | 69 | 4 |
| 9 | Tabebuia rosea (D) | 11.662 ± 0.16 efgh | ++++ | ++ | ++ | - | + | + | + | - | 11 | 69 | 4 |
| 10 | Lagerstroemia floribunda (D) | 11.820 ± 0.13 cdef | ++++ | + | ++ | - | ++ | + | + | - | 11 | 69 | 4 |
| 11 | Hopea odorata (E) | 11.479 ± 0.09 fghi | ++++ | ++ | ++ | - | + | - | - | ++ | 11 | 69 | 4 |
| 12 | Millingtonia hortensis (E) | 12.380 ± 0.11 b | +++++ | ++ | ++ | + | - | - | - | - | 10 | 63 | 4 |
| 13 | Dalbergia cochinchinensis (D) | 10.348 ± 0.23 k | +++ | ++ | ++ | - | - | + | - | ++ | 10 | 63 | 4 |
| 14 | Bauhinia purpurea (D) | 11.809 ± 0.07 cdef | ++++ | + | ++ | - | + | - | - | ++ | 10 | 63 | 4 |
| 15 | Syzygium cinereum (E) | 11.707 ± 0.06 def | ++++ | + | ++ | + | + | - | - | + | 10 | 63 | 4 |
| 16 | Terminalia bellirica (D) | 11.420 ± 0.10 fghij | ++++ | ++ | ++ | - | + | - | - | + | 10 | 63 | 4 |
| 17 | Elaeocarpus hygrophilus (E) | 11.205 ± 0.11 ghij | ++++ | + | ++ | + | + | - | - | + | 10 | 63 | 4 |
| 18 | Terminalia chebula (D) | 11.179 ± 0.07 hij | ++++ | ++ | ++ | - | + | - | - | + | 10 | 63 | 4 |
| 19 | Elaeocarpus grandiflorus (E) | 11.161 ± 0.10 ij | ++++ | - | ++ | + | + | + | - | + | 10 | 63 | 4 |
| 20 | Xylia xylocarpa (D) | 10.946 ± 0.11 j | +++ | ++ | ++ | - | + | - | - | ++ | 10 | 63 | 4 |
| 21 | Homalium tomentosum (D) | 11.677 ± 0.08 efg | ++++ | ++ | ++ | - | + | - | - | - | 9 | 56 | 3 |
| Species | Soi Temperature Reduction (°C) | Leaf Thickness | L* | LAI | Roughness |
|---|---|---|---|---|---|
| Deciduous | |||||
| Artocarpus lacucha | 5.02 ± 0.49 c | 0.31 ± 0.04 | 33.71 ± 1.32 | 1.36 ± 0.36 | hairy |
| Dalbergia cochinchinensis | 4.76 ± 0.61 d | 0.15 ± 0.01 | 34.54 ± 3.56 | 1.83 ± 0.71 | hairy |
| Bauhinia purpurea | 4.64 ± 0.50 de | 0.19 ± 0.02 | 38.47 ± 2.20 | 2.24 ± 0.35 | smooth |
| Lagerstroemia floribunda | 4.62 ± 0.47 de | 0.23 ± 0.05 | 39.46 ± 1.66 | 0.96 ± 0.40 | hairy |
| Terminalia chebula | 4.50 ± 0.55 ef | 0.27 ± 0.04 | 44.33 ± 2.76 | 1.36 ± 0.45 | smooth |
| Tabebuia rosea | 4.38 ± 0.46 fg | 0.25 ± 0.02 | 36.81 ± 4.22 | 0.60 ± 0.12 | hairy |
| Terminalia bellirica | 4.30 ± 0.77 g | 0.26 ± 0.03 | 39.74 ± 6.05 | 1.30 ± 0.81 | smooth |
| Homalium tomentosum | 3.94 ± 0.51 h | 0.25 ± 0.07 | 43.24 ± 4.82 | 0.65 ± 0.32 | smooth |
| Xylia xylocarpa | 3.94 ± 0.56 h | 0.17 ± 0.03 | 38.02 ± 1.92 | 1.30 ± 0.55 | smooth |
| Barringtonia acutangular | 3.88 ± 0.54 hi | 0.22 ± 0.02 | 39.92 ± 4.99 | 0.88 ± 0.47 | smooth |
| Afzelia xylocarpa | 3.85 ± 0.59 hi | 0.16 ± 0.02 | 40.61 ± 2.09 | 1.16 ± 0.59 | smooth |
| Average | 4.35 ± 0.40 | 0.22 ± 0.05 | 38.99 ± 3.23 | 1.24 ± 0.49 | |
| Evergreen | |||||
| Elaeocarpus grandiflorus | 5.90 ± 0.55 a | 0.27 ± 0.02 | 41.28 ± 5.84 | 2.65 ± 0.58 | hairy |
| Melaleuca quinquenervia | 5.32 ± 0.57 b | 0.33 ± 0.05 | 32.29 ± 6.95 | 1.74 ± 0.35 | hairy |
| Saraca indica | 4.36 ± 0.45 fg | 0.23 ± 0.03 | 47.26 ± 3.13 | 1.26 ± 0.13 | smooth |
| Dipterocarpus alatus | 4.34 ± 0.46 fg | 0.20 ± 0.02 | 45.06 ± 2.23 | 0.54 ± 0.15 | hairy |
| Albizia saman | 4.30 ± 0.58 g | 0.26 ± 0.02 | 33.04 ± 2.06 | 1.55 ± 0.62 | smooth |
| Syzygium cinereum | 4.29 ± 0.53 g | 0.28 ± 0.02 | 38.41 ± 3.04 | 1.20 ± 0.37 | smooth |
| Chukrasia tabularis | 4.28 ± 0.57 g | 0.14 ± 0.02 | 36.66 ± 5.27 | 2.09 ± 0.51 | smooth |
| Elaeocarpus hygrophilus | 3.74 ± 0.48 ij | 0.25 ± 0.02 | 39.59 ± 5.02 | 0.50 ± 0.12 | smooth |
| Hopea odorata | 3.68 ± 0.45 jk | 0.19 ± 0.02 | 44.16 ± 2.51 | 0.61 ± 0.08 | smooth |
| Millingtonia hortensis | 3.56 ± 0.53 k | 0.14 ± 0.03 | 34.45 ± 2.98 | 1.03 ± 0.63 | smooth |
| Average | 4.38 ± 0.69 | 0.23 ± 0.06 | 39.22 ± 4.94 | 1.31 ± 0.67 | |
| t-test (p-value) | 0.91 (NS) | 0.83 (NS) | 0.37 (NS) | 0.77 (NS) | |
| Species | Air Temperature Reduction (°C) | LAI | Crown Cover (m) | DBH (cm) | Total Height (m) |
|---|---|---|---|---|---|
| Deciduous | |||||
| Bauhinia purpurea | 3.28 ± 1.77 a | 2.76 | 4.4 | 9.6 | 6.7 |
| Homalium tomentosum | 2.74 ± 1.94 ab | 1.19 | 4.1 | 10.5 | 9.2 |
| Lagerstroemia floribunda | 2.50 ± 1.71 abc | 1.17 | 2.9 | 13.5 | 4.1 |
| Artocarpus lacucha | 2.06 ± 1.41 bcd | 1.33 | 3.4 | 15.6 | 8.4 |
| Baringtonia acutangula | 2.05 ± 1.04 bcd | 1.67 | 3 | 16.4 | 5.5 |
| Dalbergia cochinchinensis | 2.04 ± 1.39 bcd | 0.6 | 4.9 | 14 | 7.9 |
| Terminalia bellirica | 1.69 ± 0.99 bcd | 1.71 | 2.1 | 13.1 | 10.1 |
| Afzelia xylocarpa | 1.66 ± 1.34 bcd | 0.61 | 4 | 13.6 | 9.6 |
| Tabebuia rosea | 1.59 ± 1.44 cd | 0.61 | 3.5 | 13.9 | 8.8 |
| Xylia xylocarpa | 1.49 ± 1.46 cd | 1.22 | 2.7 | 13.6 | 8.9 |
| Terminalia chebula | 1.42 ± 1.37 cd | 1.05 | 1.2 | 10.2 | 6 |
| Average | 2.05 ± 0.58 | 1.27 ± 0.63 | 3.29 ± 1.07 | 13.09 ± 2.16 | 7.75 ± 1.91 |
| Species | Air temperature reduction (°C) | LAI | Crown cover (m) | DBH (cm) | Total height (m) |
| Evergreen | |||||
| Elaeocarpus grandiflorus | 2.72 ± 1.07 ab | 3.2 | 2.5 | 11.9 | 3.3 |
| Melaleuca quinquenervia | 2.54 ± 1.16 abc | 1.48 | 1.8 | 15.3 | 11 |
| Albizia saman | 2.22 ± 1.30 abc | 0.69 | 21 | 18 | 6.4 |
| Chukrasia tabularis | 2.12 ± 1.29 abcd | 1.51 | 2.8 | 13.1 | 7.9 |
| Syzygium cinereum | 2.01 ± 1.33 bcd | 0.77 | 1.1 | 17.4 | 8.5 |
| Hopea odorata | 1.77 ± 1.35 bcd | 0.54 | 1.3 | 12.3 | 8.8 |
| Millingtonia hortensis | 1.69 ± 1.10 bcd | 1.29 | 1.7 | 14.7 | 9.2 |
| Saraca indica | 1.60 ± 1.39 bcd | 1.04 | 3.2 | 12.8 | 2.8 |
| Dipterocarpus alatus | 0.95 ± 1.42 d | 0.68 | 3.2 | 14.9 | 9.2 |
| Elaeocarpus hygrophilus | 0.94 ± 1.33 d | 0.59 | 2.8 | 12.1 | 6.4 |
| Average | 1.86 ± 0.60 | 1.18 ± 0.80 | 4.14 ± 5.97 | 14.25 ± 2.19 | 7.35 ± 2.64 |
| t-test (p-value) | 0.47 (NS) | 0.78 (NS) | 0.65 (NS) | 0.24 (NS) | 0.70 (NS) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yarnvudhi, A.; Leksungnoen, N.; Andriyas, T.; Tor-Ngern, P.; Premashthira, A.; Wachrinrat, C.; Marod, D.; Hermhuk, S.; Pattanakiat, S.; Nakashizuka, T.; et al. Assessing the Cooling and Air Pollution Tolerance among Urban Tree Species in a Tropical Climate. Plants 2022, 11, 3074. https://doi.org/10.3390/plants11223074
Yarnvudhi A, Leksungnoen N, Andriyas T, Tor-Ngern P, Premashthira A, Wachrinrat C, Marod D, Hermhuk S, Pattanakiat S, Nakashizuka T, et al. Assessing the Cooling and Air Pollution Tolerance among Urban Tree Species in a Tropical Climate. Plants. 2022; 11(22):3074. https://doi.org/10.3390/plants11223074
Chicago/Turabian StyleYarnvudhi, Arerut, Nisa Leksungnoen, Tushar Andriyas, Pantana Tor-Ngern, Aerwadee Premashthira, Chongrak Wachrinrat, Dokrak Marod, Sutheera Hermhuk, Sura Pattanakiat, Tohru Nakashizuka, and et al. 2022. "Assessing the Cooling and Air Pollution Tolerance among Urban Tree Species in a Tropical Climate" Plants 11, no. 22: 3074. https://doi.org/10.3390/plants11223074