


Receptor-like Kinases (LRR-RLKs) in Response of Plants to Biotic and Abiotic Stresses

 , and
, and
Abstract
:1. Introduction
2. Abundance of LRR-RLKs Genes in the Plant Genome
3. Stress-Related LRR-RLKs in Plants
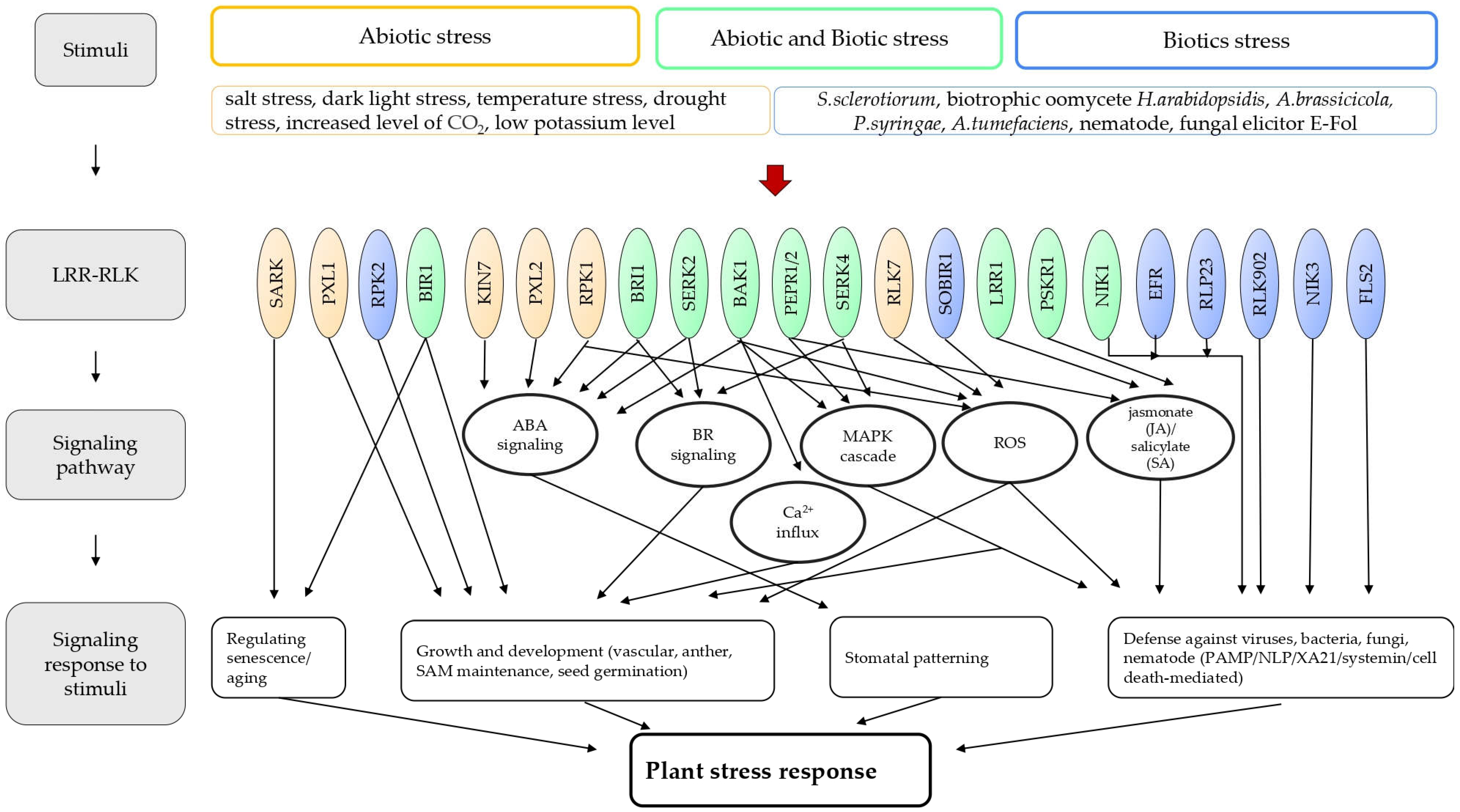
4. Interactions among the Stress-Related LRR-LRKs

{kind=link}
| LRR-RLK | SERK2 | SERK1 | SERK4/BKK1 | SERK3/BAK1 | SARK/Cik3 | NIK1 | NIK3/Cik1 | References |
|---|---|---|---|---|---|---|---|---|
| co-receptors (LRR-RLKs from LRR II family accordingly classification of [22]) | ||||||||
| BIR1 | heterodimer (BiFC) | heterodimer (BiFC) | heterodimer (BiFC) | heterodimer (genetically, in vivo, pull-down) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | NA | [14,152] |
| FLS2 | NA | heterodimer (in vivo) | heterodimer (in vivo) | heterodimer (in vivo, CoIP, gel filtration) | NA | heterodimer (in vitro, pull down) | NA | [16,153,154] |
| ERL1 | heterodimer (CoIP) | heterodimer (CoIP) | heterodimer (CoIP) | heterodimer (in vivo, CoIP) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | NA | [147,152,153] |
| BRI1 | NA | heterodimer (genetically, in vivo) | heterodimer (in vivo) | heterodimer (genetically, in vivo, in vitro) | NA | NA | NA | [146,149,155] |
| EFR | NA | heterodimer (in vivo) | heterodimer | heterodimer (genetically, CoIP, in vivo) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | NA | [16,152,156] |
| ER | heterodimer (genetically, CoIP) | heterodimer (genetically, CoIP) | heterodimer (solid-phase assay) | heterodimer (genetically, pull-down, in vivo) | heterodimer (solid phase assay) | heterodimer (solid-phase assay) | NA | [29,147,152,153] |
| PXY | heterodimer (genetically, in vivo, in vitro) | heterodimer (genetically, in vivo, in vitro) | NA | heterodimer (genetically, in vivo, in vitro) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | [23,153] |
| co-receptors (LRR-RLKs from LRR II family accordingly classification of [22]) | ||||||||
| HSL2 | heterodimer (CoIP) | heterodimer (CoIP) | NA | heterodimer (CoIP) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | NA | [152,157] |
| BAK1 | heterodimer (genetically, CoIP) | heterodimer (BiFC, CoIP) | heterodimer (genetically) | NA | heterodimer (solid-phase assay) | heterodimer (genetically, in vivo, in vitro) | heterodimer (solid-phase assay) | [127,130,131,132] |
| SERK1 | heterodimer (genetically, in vivo) | homodimer (genetically, in vivo) | heterodimer (solid-phase assay) | heterodimer (BiFC, CoIP) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | NA | [152,153,155,158] |
| SERK2 | homodimer (genetically, in vivo) | heterodimer (genetically, in vivo) | NA | NA | NA | NA | heterodimer (solid-phase assay) | [152,158] |
| EMS1 | heterodimer (genetically, kinase assay, BiFC) | heterodimer (genetically, kinase assay, BiFC) | NA | NA | NA | NA | NA | [135] |
| SERK4 | heterodimer (genetically, CoIP) | heterodimer (solid-phase assay) | NA | heterodimer (genetically) | Heterodimer (solid-phase assay) | Heterodimer (solid-phase assay) | NA | [16,152] |
| NIK1 | NA | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | Heterodimer (genetically, in vivo, in vitro) | Heterodimer (solid-phase assay) | NA | NA | [152,153,154] |
| PSKR1 | heterodimer (solid-phase assay, co-sedimentation in solution) | heterodimer(molecular sieving) | NA | heterodimer (genetically, CoIP, gel filtration) | heterodimer (solid-phase assay) | NA | NA | [143,153] |
| co-receptors (LRR-RLKs from LRR II family accordingly classification of [22]) | ||||||||
| SOBIR1 | heterodimer (solid-phase assay) | NA | NA | heterodimer (genetically, CoIP) | NA | NA | NA | [30,153] |
| RPK1 | NA | NA | NA | heterodimer (genetically, pull-down, kinase assay) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | NA | [109,152] |
| PEPR 1 | NA | NA | NA | heterodimer (pull-down, gel filtration) | NA | NA | heterodimer (solid-phase assay) | [153,159] |
| ERL2 | NA | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | heterodimer (genetically) | heterodimer (solid-phase assay) | heterodimer (solid-phase assay) | NA | [29,152] |
| RPK2 | NA | NA | NA | NA | heterodimer (in vivo, pull-down) | heterodimer (solid-phase assay) | heterodimer (genetically) | [129,153] |
| CLV1 | NA | NA | NA | NA | heterodimer (in vivo, pull-down) | NA | heterodimer (genetically) | [129] |
| BAM1 | heterodimer (solid-phase assay) | NA | NA | NA | heterodimer (pull-down) | heterodimer (solid-phase assay) | heterodimer (genetically, BiFC, CoIP) | [33,153] |
| BAM2 | NA | NA | NA | NA | heterodimer (pull-down) | heterodimer (solid-phase assay) | heterodimer (genetically, BiFC, CoIP) | [33,153] |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gish, L.A.; Clark, S.E. The RLK/Pelle Family of Kinases. Plant J. 2011, 66, 117–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osakabe, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S.P. Sensing the Environment: Key Roles of Membrane-Localized Kinases in Plant Perception and Response to Abiotic Stress. J. Exp. Bot. 2013, 64, 445–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dievart, A.; Gottin, C.; Peacuterin, C.; Ranwez, V.; Chantret, N. Origin and Diversity of Plant Receptor-like Kinases. Annu. Rev. Plant Biol. 2020, 71, 131–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dievart, A.; Clark, S.E. LRR-Containing Receptors Regulating Plant Development and Defense. Development 2004, 131, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Kucukoglu, M.; Zhang, L.; Chen, P.; Decker, D.; Nilsson, O.; Jones, B.; Sandberg, G.; Zheng, B. The Arabidopsis LRR-RLK, PXC1, Is a Regulator of Secondary Wall Formation Correlated with the TDIF-PXY/TDR-WOX4 Signaling Pathway. BMC Plant Biol. 2013, 13, 94. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Ji, Y.; Wang, J.; Yang, S.; Song, Y. Vascular Expression of Populus LRR-RLK Genes and the Effects of Their Overexpression on Wood Formation. Mol. Breed. 2015, 35, 220. [Google Scholar] [CrossRef]
- Li, X.; Salman, A.; Guo, C.; Yu, J.; Cao, S.; Gao, X.; Li, W.; Li, H.; Guo, Y. Identification and Characterization of LRR-RLK Family Genes in Potato Reveal Their Involvement in Peptide Signaling of Cell Fate Decisions and Biotic/Abiotic Stress Responses. Cells 2018, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wen, J.; Lease, K.A.; Doke, J.T.; Tax, F.E.; Walker, J.C. BAK1, an Arabidopsis LRR Receptor-like Protein Kinase, Interacts with BRI1 and Modulates Brassinosteroid Signaling. Cell 2002, 110, 213–222. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wang, P.; Claus, L.A.N.; Savatin, D.V.; Xu, G.; Wu, S.; Meng, X.; Russinova, E.; He, P.; Shan, L. Proteolytic Processing of Serk3/Bak1 Regulates Plant Immunity, Development, and Cell Death. Plant Physiol. 2019, 180, 543–558. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Moon, J.C.; Park, Y.C.; Kim, J.H.; Kim, D.S.; Jang, C.S. Molecular Dissection of the Response of a Rice Leucine-Rich Repeat Receptor-like Kinase (LRR-RLK) Gene to Abiotic Stresses. J. Plant Physiol. 2014, 171, 1465–1653. [Google Scholar] [CrossRef]
- Liu, X.S.; Liang, C.C.; Hou, S.G.; Wang, X.; Chen, D.H.; Shen, J.L.; Zhang, W.; Wang, M. The LRR-RLK Protein HSL3 Regulates Stomatal Closure and the Drought Stress Response by Modulating Hydrogen Peroxide Homeostasis. Front. Plant Sci. 2020, 11, 548034. [Google Scholar] [CrossRef]
- Lin, F.; Li, S.; Wang, K.; Tian, H.; Gao, J.; Zhao, Q.; Du, C. A Leucine-Rich Repeat Receptor-like Kinase, OsSTLK, Modulates Salt Tolerance in Rice. Plant Sci. 2020, 296, 110465. [Google Scholar] [CrossRef]
- Albert, I.; Böhm, H.; Albert, M.; Feiler, C.E.; Imkampe, J.; Wallmeroth, N.; Brancato, C.; Raaymakers, T.M.; Oome, S.; Zhang, H.; et al. An RLP23-SOBIR1-BAK1 Complex Mediates NLP-Triggered Immunity. Nat. Plants 2015, 1, 15140. [Google Scholar] [CrossRef]
- Gao, M.; Wang, X.; Wang, D.; Xu, F.; Ding, X.; Zhang, Z.; Bi, D.; Cheng, Y.T.; Chen, S.; Li, X.; et al. Regulation of Cell Death and Innate Immunity by Two Receptor-like Kinases in Arabidopsis. Cell Host Microbe 2009, 6, 34–44. [Google Scholar] [CrossRef] [Green Version]
- Russinova, E.; Borst, J.W.; Kwaaitaal, M.; Caño-Delgado, A.; Yin, Y.; Chory, J.; De Vries, S.C. Heterodimerization and Endocytosis of Arabidopsis Brassinosteroid Receptors BRI1 and AtSERK3 (BAK1). Plant Cell 2004, 16, 3216–3229. [Google Scholar] [CrossRef] [Green Version]
- Roux, M.; Schwessinger, B.; Albrecht, C.; Chinchilla, D.; Jones, A.; Holton, N.; Malinovsky, F.G.; Tör, M.; de Vries, S.; Zipfel, C. The Arabidopsis Leucine-Rich Repeat Receptor-like Kinases BAK1/SERK3 and BKK1/SERK4 Are Required for Innate Immunity to Hemibiotrophic and Biotrophic Pathogens. Plant Cell 2011, 23, 2440–2455. [Google Scholar] [CrossRef] [Green Version]
- Nam, K.H.; Li, J. BRI1/BAK1, a Receptor Kinase Pair Mediating Brassinosteroid Signaling. Cell 2002, 110, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Liebrand, T.W.H.; Van Den Berg, G.C.M.; Zhang, Z.; Smit, P.; Cordewener, J.H.G.; America, A.H.P.; Sklenar, J.; Jones, A.M.E.; Tameling, W.I.L.; Robatzek, S.; et al. Receptor-like Kinase SOBIR1/EVR Interacts with Receptor-like Proteins in Plant Immunity against Fungal Infection. Proc. Natl. Acad. Sci. USA 2013, 110, 10010–10015. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Ding, Y.; Jiang, Q.; Wang, F.; Sun, J.; Zhu, C. The Role of Receptor-like Protein Kinases (RLKs) in Abiotic Stress Response in Plants. Plant Cell Rep. 2017, 36, 235–242. [Google Scholar] [CrossRef]
- Ma, X.; Xu, G.; He, P.; Shan, L. SERKing Coreceptors for Receptors. Trends Plant Sci. 2016, 21, 1017–1033. [Google Scholar] [CrossRef]
- Diévart, A.; Clark, S.E. Using Mutant Alleles to Determine the Structure and Function of Leucine-Rich Repeat Receptor-like Kinases. Curr. Opin. Plant Biol. 2003, 6, 507–516. [Google Scholar] [CrossRef]
- Xi, L.; Wu, X.N.; Gilbert, M.; Schulze, W.X. Classification and Interactions of LRR Receptors and Co-Receptors within the Arabidopsis Plasma Membrane—An Overview. Front. Plant Sci. 2019, 10, 472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Lin, X.; Han, Z.; Wang, J.; Qu, L.J.; Chai, J. SERK Family Receptor-like Kinases Function as Co-Receptors with PXY for Plant Vascular Development. Mol. Plant 2016, 9, 1406–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohmann, U.; Nicolet, J.; Moretti, A.; Hothorn, L.A.; Hothorn, M. The SERK3 Elongated Allele Defines a Role for BIR Ectodomains in Brassinosteroid Signalling. Nat. Plants 2018, 4, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Hohmann, U.; Santiago, J.; Nicolet, J.; Olsson, V.; Spiga, F.M.; Hothorn, L.A.; Butenko, M.A.; Hothorn, M. Mechanistic Basis for the Activation of Plant Membrane Receptor Kinases by SERK-Family Coreceptors. Proc. Natl. Acad. Sci. USA 2018, 115, 3488–3493. [Google Scholar] [CrossRef] [Green Version]
- Chinchilla, D.; Shan, L.; He, P.; de Vries, S.; Kemmerling, B. One for All: The Receptor-Associated Kinase BAK1. Trends Plant Sci. 2009, 14, 535–541. [Google Scholar] [CrossRef] [Green Version]
- Postel, S.; Küfner, I.; Beuter, C.; Mazzotta, S.; Schwedt, A.; Borlotti, A.; Halter, T.; Kemmerling, B.; Nürnberger, T. The Multifunctional Leucine-Rich Repeat Receptor Kinase BAK1 Is Implicated in Arabidopsis Development and Immunity. Eur. J. Cell Biol. 2010, 89, 169–174. [Google Scholar] [CrossRef]
- Lin, W.; Li, B.; Lu, D.; Chen, S.; Zhu, N.; He, P.; Shan, L. Tyrosine Phosphorylation of Protein Kinase Complex BAK1/BIK1 Mediates Arabidopsis Innate Immunity. Proc. Natl. Acad. Sci. USA 2014, 111, 3632–3637. [Google Scholar] [CrossRef] [Green Version]
- Jordá, L.; Sopeña-Torres, S.; Escudero, V.; Nuñez-Corcuera, B.; Delgado-Cerezo, M.; Torii, K.U.; Molina, A. ERECTA and BAK1 Receptor like Kinases Interact to Regulate Immune Responses in Arabidopsis. Front. Plant Sci. 2016, 7, 897. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Huang, X.; Li, M.; He, P.; Zhang, Y. Loss-of-Function of Arabidopsis Receptor-like Kinase BIR1 Activates Cell Death and Defense Responses Mediated by BAK1 and SOBIR1. New Phytol. 2016, 212, 637–645. [Google Scholar] [CrossRef]
- Wang, Z.; Meng, P.; Zhang, X.; Ren, D.; Yang, S. BON1 Interacts with the Protein Kinases BIR1 and BAK1 in Modulation of Temperature-Dependent Plant Growth and Cell Death in Arabidopsis. Plant J. 2011, 67, 1081–1093. [Google Scholar] [CrossRef]
- Bleckmann, A.; Weidtkamp-Peters, S.; Seidel, C.A.M.; Simon, R. Stem Cell Signaling in Arabidopsis Requires CRN to Localize CLV2 to the Plasma Membrane. Plant Physiol. 2010, 152, 166–176. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Hu, C.; Zhu, Y.; Cheng, K.; Li, X.; Wei, Z.; Xue, L.; Lin, F.; Shi, H.; Yi, J.; et al. CIK Receptor Kinases Determine Cell Fate Specificatioduring Early Anther Development in Arabidopsis. Plant Cell 2018, 30, 2383–2401. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.L.; Du, L.; Huang, Y.; Gao, S.M.; Yu, M. Origin and Diversification of Leucine-Rich Repeat Receptor-like Protein Kinase (LRR-RLK) Genes in Plants. BMC Evol. Biol. 2017, 17, 47. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Feng, T.; Zhang, D.; Zhuo, R.; Liu, M. Identification and Comprehensive Analysis of the Characteristics and Roles of Leucine-Rich Repeat Receptor-like Protein Kinase (LRR-RLK) Genes in Sedum alfredii Hance Responding to Cadmium Stress. Ecotoxicol. Environ. Saf. 2019, 167, 95–106. [Google Scholar] [CrossRef]
- Liu, P.L.; Xie, L.L.; Li, P.W.; Mao, J.F.; Liu, H.; Gao, S.M.; Shi, P.H.; Gong, J.Q. Duplication and Divergence of Leucine-Rich Repeat Receptor-like Protein Kinase (LRR-RLK) Genes in Basal Angiosperm Amborella trichopoda. Front. Plant Sci. 2016, 7, 1952. [Google Scholar] [CrossRef]
- Dufayard, J.F.; Bettembourg, M.; Fischer, I.; Droc, G.; Guiderdoni, E.; Périn, C.; Chantret, N.; Diévart, A. New Insights on Leucine-Rich Repeats Receptor-like Kinase Orthologous Relationships in Angiosperms. Front. Plant Sci. 2017, 8, 381. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Xia, W.; Yang, Y.; Mason, A.S.; Lei, X.; Ma, Z. Characterization and Evolution of Conserved MicroRNA through Duplication Events in Date Palm (Phoenix dactylifera). PLoS ONE 2013, 8, e71435. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The Genome of the Cucumber, Cucumis sativus, L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Li, L.; Wang, P.; Zhang, S.; Wu, J. Genome-Wide Characterization, Evolution, and Expression Analysis of the Leucine-Rich Repeat Receptor-like Protein Kinase (LRR-RLK) Gene Family in Rosaceae Genomes. BMC Genom. 2017, 18, 763. [Google Scholar] [CrossRef]
- Meng, J.; Yang, J.; Peng, M.; Liu, X.; He, H. Genome-Wide Characterization, Evolution, and Expression Analysis of the Leucine-Rich Repeat Receptor-like Protein Kinase (Lrr-Rlk) Gene Family in Medicago truncatula. Life 2020, 10, 176. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.; Peng, R.H.; Cheng, Z.M.; Zhang, J.; Cai, B.; Zhang, Z.; Gao, F.; Zhu, B.; Fu, X.Y.; Jin, X.F.; et al. Genome-Wide Analysis of the Putative AP2/ERF Family Genes in Vitis vinifera. Sci. Hortic. 2009, 123, 73–81. [Google Scholar] [CrossRef]
- Wu, Y.; Xun, Q.; Guo, Y.; Zhang, J.; Cheng, K.; Shi, T.; He, K.; Hou, S.; Gou, X.; Li, J. Genome-Wide Expression Pattern Analyses of the Arabidopsis Leucine-Rich Repeat Receptor-like Kinases. Mol. Plant 2016, 9, 289–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakawa, H.; Tsuchimoto, S.; Sakai, H.; Nakayama, S.; Fujishiro, T.; Kishida, Y.; Kohara, M.; Watanabe, A.; Yamada, M.; Aizu, T.; et al. Upgraded Genomic Information of Jatropha curcas L. Plant Biotechnol. 2012, 29, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Wang, Y.; Yin, H.; Gao, M.; Zhang, Q.; Chen, Y. Genome-Wide Identification and Characterization of the LRR-RLK Gene Family in Two Vernicia Species. Int. J. Genom. 2015, 2015, 823427. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Mo, W.; Li, Y.; Li, W.; Dong, X.; Liu, M.; Jiang, L.; Zhang, L. Deciphering the Roles of Leucine-Rich Repeat Receptor-like Protein Kinases (LRR-RLKs) in Response to Fusarium Wilt in the Vernicia fordii (Tung Tree). Phytochemistry 2021, 185, 112686. [Google Scholar] [CrossRef]
- Sakamoto, T.; Deguchi, M.; Brustolini, O.J.B.; Santos, A.A.; Silva, F.F.; Fontes, E.P.B. The Tomato RLK Superfamily: Phylogeny and Functional Predictions about the Role of the LRRII-RLK Subfamily in Antiviral Defense. BMC Plant Biol. 2012, 12, 229. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Zhang, Q.; Shi, Y.; Sun, Z.; Zhang, Q.; Hou, S.; Wu, R.; Jiang, L.; Zhao, X.; Guo, Y. Genome-Wide Analysis of the NAC Domain Transcription Factor Gene Family in Theobroma cacao. Genes 2020, 11, 35. [Google Scholar] [CrossRef] [Green Version]
- Song, W.; Wang, B.; Li, X.; Wei, J.; Chen, L.; Zhang, D.; Zhang, W.; Li, R. Identification of Immune Related LRR-Containing Genes in Maize (Zea mays L.) by Genome-Wide Sequence Analysis. Int. J. Genom. 2015, 2015, 231358. [Google Scholar] [CrossRef] [Green Version]
- Dezhsetan, S. Genome Scanning for Identification and Mapping of Receptor-like Kinase (RLK) Gene Superfamily in Solanum tuberosum. Physiol. Mol. Biol. Plants 2017, 23, 755–765. [Google Scholar] [CrossRef]
- Wang, J.; Hu, T.; Wang, W.; Hu, H.; Wei, Q.; Bao, C. Investigation of Evolutionary and Expressional Relationships in the Function of the Leucine-Rich Repeat Receptor-like Protein Kinase Gene Family (LRR-RLK) in the Radish (Raphanus sativus L.). Sci. Rep. 2019, 9, 6937. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Wang, S.; Ma, D.; Liu, C. Genome-Wide Analysis of LRR-RLK Gene Family in Four Gossypium Species and Expression Analysis during Cotton Development and Stress Responses. Genes 2018, 9, 592. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, D.M.; Scholte, L.L.S.; Silva, N.V.; Oliveira, G.C.; Zipfel, C.; Takita, M.A.; De Souza, A.A. LRR-RLK Family from Two Citrus Species: Genome-Wide Identification and Evolutionary Aspects. BMC Genom. 2016, 17, 623. [Google Scholar] [CrossRef] [Green Version]
- Rameneni, J.J.; Lee, Y.; Dhandapani, V.; Yu, X.; Choi, S.R.; Oh, M.H.; Lim, Y.P. Genomic and Post-Translational Modification Analysis of Leucine-Rich-Repeat Receptor-like Kinases in Brassica rapa. PLoS ONE 2015, 10, e0142255. [Google Scholar] [CrossRef]
- Li, W.; Chen, M.; Wang, E.; Hu, L.; Hawkesford, M.J.; Zhong, L.; Chen, Z.; Xu, Z.; Li, L.; Zhou, Y.; et al. Genome-Wide Analysis of Autophagy-Associated Genes in Foxtail Millet (Setaria italica L.) and Characterization of the Function of SiATG8a in Conferring Tolerance to Nitrogen Starvation in Rice. BMC Genom. 2016, 17, 797. [Google Scholar] [CrossRef] [Green Version]
- Truco, M.J.; Quiros, C.F. Structure and Organization of the B Genome Based on a Linkage Map in Brassica nigra. Theor. Appl. Genet. 1994, 89, 590–598. [Google Scholar] [CrossRef]
- Sun, X.; Wang, G.L. Genome-Wide Identification, Characterization and Phylogenetic Analysis of the Rice LRR-Kinases. PLoS ONE 2011, 6, e16079. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Chang, S.; Hartman, G.L.; Domier, L.L. Assembly and Annotation of a Draft Genome Sequence for Glycine latifolia, a Perennial Wild Relative of Soybean. Plant J. 2018, 95, 71–85. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Yu, M.; Zou, C.; Lu, C.; Yu, D.; Cheng, H.; Jiang, P.; Feng, X.; Zhang, Y.; Wang, Q.; et al. Genome-Wide Comparative Analysis of RNA-Binding Glycine-Rich Protein Family Genes between Gossypium arboreum and Gossypium raimondii. PLoS ONE 2018, 14, e0218938. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Han, Y.; Meng, D.; Li, D.; Jin, Q.; Lin, Y.; Cai, Y. Structural, Evolutionary, and Functional Analysis of the Class III Peroxidase Gene Family in Chinese Pear (Pyrus bretschneideri). Front. Plant Sci. 2016, 7, 1874. [Google Scholar] [CrossRef]
- Zan, Y.; Ji, Y.; Zhang, Y.; Yang, S.; Song, Y.; Wang, J. Genome-Wide Identification, Characterization and Expression Analysis of Populus Leucine-Rich Repeat Receptor-like Protein Kinase Genes. BMC Genom. 2013, 14, 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Bayer, P.E.; Tirnaz, S.; Edwards, D.; Batley, J. Genome-Wide Identification and Evolution of Receptor-like Kinases (RLKs) and Receptor like Proteins (RLPs) in Brassica juncea. Biology 2021, 10, 17. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Guo, Y.; Qiu, L.J. Genome-Wide Identification and Evolutionary Analysis of Leucine-Rich Repeat Receptor-like Protein Kinase Genes in Soybean. BMC Plant Biol. 2016, 16, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shumayla; Sharma, S.; Kumar, R.; Mendu, V.; Singh, K.; Upadhyay, S.K. Genomic Dissection and Expression Profiling Revealed Functional Divergence in Triticum aestivum Leucine Rich Repeat Receptor like Kinases (TaLRRKs). Front. Plant Sci. 2016, 7, 1374. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wu, M.H.; Xiao, D.; Huang, R.L.; Zhan, J.; Wang, A.Q.; He, L.F. Genome-Wide Identification and Evolutionary Analysis of RLKs Involved in the Response to Aluminium Stress in Peanut. BMC Plant Biol. 2021, 21, 281. [Google Scholar] [CrossRef]
- Mishra, D.; Suri, G.S.; Kaur, G.; Tiwari, M. Comprehensive Analysis of Structural, Functional, and Evolutionary Dynamics of Leucine Rich Repeats-RLKs in Thinopyrum elongatum. Int. J. Biol. Macromol. 2021, 183, 513–527. [Google Scholar] [CrossRef]
- Zoulias, N.; Harrison, E.L.; Casson, S.A.; Gray, J.E. Molecular Control of Stomatal Development. Biochem. J. 2018, 475, 441–454. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Hu, C.; Cui, Y.; Zeng, L.; Li, S.; Zhu, M.; Meng, F.; Huang, S.; Long, L.; Yi, J.; et al. Conserved and Differentiated Functions of CIK Receptor Kinases in Modulating Stem Cell Signaling in Arabidopsis. Mol. Plant 2021, 14, 1119–1134. [Google Scholar] [CrossRef]
- Ou, Y.; Kui, H.; Li, J. Receptor-like Kinases in Root Development: Current Progress and Future Directions. Mol. Plant 2021, 14, 166–185. [Google Scholar] [CrossRef]
- Nadeau, J.A. Stomatal Development: New Signals and Fate Determinants. Curr. Opin. Plant Biol. 2009, 12, 29–35. [Google Scholar] [CrossRef]
- Cammarata, J.; Scanlon, M.J. A Functionally Informed Evolutionary Framework for the Study of LRR-RLKs during Stem Cell Maintenance. J. Plant Res. 2020, 133, 331–342. [Google Scholar] [CrossRef]
- Kim, B.H.; Kim, S.Y.; Nam, K.H. Assessing the Diverse Functions of BAK1 and Its Homologs in Arabidopsis, beyond BR Signaling and PTI Responses. Mol. Cells 2013, 35, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Dai, C.; Lee, M.M.; Kwak, J.M.; Nam, K.H. BRI1-Associated Receptor Kinase 1 Regulates Guard Cell ABA Signaling Mediated by Open Stomata 1 in Arabidopsis. Mol. Plant 2016, 9, 447–460. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, A.; Pastorczyk, M.; Piślewska-Bednarek, M.; Nishiuchi, T.; Ono, E.; Suemoto, H.; Ishikawa, A.; Frerigmann, H.; Kaido, M.; Mise, K.; et al. Tryptophan-Derived Metabolites and BAK1 Separately Contribute to Arabidopsis Postinvasive Immunity against Alternaria brassicicola. Sci. Rep. 2021, 11, 1488. [Google Scholar] [CrossRef]
- Peng, H.C.; Kaloshian, I. The Tomato Leucine-Rich Repeat Receptor-like Kinases SlSERK3A and SlSERK3B Have Overlapping Functions in Bacterial and Nematode Innate Immunity. PLoS ONE 2014, 9, e93302. [Google Scholar] [CrossRef] [Green Version]
- Chaparro-Garcia, A.; Wilkinson, R.C.; Gimenez-Ibanez, S.; Findlay, K.; Coffey, M.D.; Zipfel, C.; Rathjen, J.P.; Kamoun, S.; Schornack, S. The Receptor-like Kinase Serk3/Bak1 Is Required for Basal Resistance against the Late Blight Pathogen Phytophthora Infestans in Nicotiana benthamiana. PLoS ONE 2011, 6, e16608. [Google Scholar] [CrossRef]
- Chen, X.; Zuo, S.; Schwessinger, B.; Chern, M.; Canlas, P.E.; Ruan, D.; Zhou, X.; Wang, J.; Daudi, A.; Petzold, C.J.; et al. An XA21-Associated Kinase (OsSERK2) Regulates Immunity Mediated by the XA21 and XA3 Immune Receptors. Mol. Plant 2014, 7, 874–892. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Xiong, L.; Yang, Y. Rice SERK1 Gene Positively Regulates Somatic Embryogenesis of Cultured Cell and Host Defense Response against Fungal Infection. Planta 2005, 222, 107–117. [Google Scholar] [CrossRef]
- Derbyshire, M.; Mbengue, M.; Barascud, M.; Navaud, O.; Raffaele, S. Small RNAs from the Plant Pathogenic Fungus Sclerotinia sclerotiorum Highlight Host Candidate Genes Associated with Quantitative Disease Resistance. Mol. Plant Pathol. 2019, 20, 1279–1297. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Yin, W.; Liu, D.; Zhang, X.; Yu, Z.; Huang, W.; Liu, J.; Yang, Y.; Meng, W.; Niu, M.; et al. Regulation of Brassinosteroid Signaling and Salt Resistance by SERK2 and Potential Utilization for Crop Improvement in Rice. Front. Plant Sci. 2020, 11, 621859. [Google Scholar] [CrossRef]
- Li, X.; Ahmad, S.; Ali, A.; Guo, C.; Li, H.; Yu, J.; Zhang, Y.; Gao, X.; Guo, Y. Characterization of Somatic Embryogenesis Receptor-like Kinase 4 as a Negative Regulator of Leaf Senescence in Arabidopsis. Cells 2019, 8, 50. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Li, X.; Meisenhelder, J.; Hunter, T.; Yoshida, S.; Asami, T.; Chory, J. Autoregulation and Homodimerization Are Involved in the Activation of the Plant Steroid Receptor BRI1. Dev. Cell 2005, 8, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Goddard, R.; Peraldi, A.; Ridout, C.; Nicholson, P. Enhanced Disease Resistance Caused by BRI1 Mutation Is Conserved between Brachypodium distachyon and Barley (Hordeum vulgare). Mol. Plant-Microbe Interact. 2014, 27, 1095–1106. [Google Scholar] [CrossRef] [Green Version]
- Clouse, S.D.; Langford, M.; McMorris, T.C. A Brassinosteroid-Insensitive Mutant in Arabidopsis thaliana Exhibits Multiple Defects in Growth and Development. Plant Physiol. 1996, 111, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Lease, K.A.; Tax, F.E.; Walker, J.C. BRS1, a Serine Carboxypeptidase, Regulates BRI1 Signaling in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2001, 98, 5916–5921. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Yin, Y.; Fei, S. Down-Regulation of BdBRI1, a Putative Brassinosteroid Receptor Gene Produces a Dwarf Phenotype with Enhanced Drought Tolerance in Brachypodium distachyon. Plant Sci. 2015, 234, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Holton, N.; Harrison, K.; Yokota, T.; Bishop, G.J. Tomato BRI1 and Systemin Wound Signalling. Plant Signal. Behav. 2008, 3, 54–55. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Wang, H.; Gan, S.; Hussein, M.A.M.; Wang, Q.; Wang, X.; Zhang, Y.; Wang, X. Molecular Identification and Functional Analysis of Brbri1 as Brassinosteroid Receptor Gene in Brassica rapa. Pak. J. Bot. 2018, 50, 85–95. [Google Scholar]
- Sharma, A.; Matsuoka, M.; Tanaka, H.; Komatsu, S. Antisense Inhibition of a BRI1 Receptor Reveals Additional Protein Kinase Signaling Components Downstream to the Perception of Brassinosteroids in Rice. FEBS Lett. 2001, 507, 346–350. [Google Scholar] [CrossRef] [Green Version]
- Scheer, J.M.; Pearce, G.; Ryan, C.A. Generation of Systemin Signaling in Tobacco by Transformation with the Tomato Systemin Receptor Kinase Gene. Proc. Natl. Acad. Sci. USA 2003, 100, 10114–10117. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.G.; Zhu, T.; Zou, L.J.; Han, X.Y.; Zhou, X.; Xi, D.H.; Zhang, D.W.; Lin, H.H. Orchestration of Hydrogen Peroxide and Nitric Oxide in Brassinosteroid-Mediated Systemic Virus Resistance in Nicotiana benthamiana. Plant J. 2016, 85, 478–493. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Breja, P.; Khurana, J.P.; Khurana, P. Wheat Brassinosteroid-Insensitive1 (TaBRI1) Interacts with Members of TaSERK Gene Family and Cause Early Flowering and Seed Yield Enhancement in Arabidopsis. PLoS ONE 2016, 11, e0153273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, S.; Huang, S.; Wang, S.; Cheng, D.; Liu, J.; Lv, S.; Li, Q.; Wang, X. Enhancing Brassinosteroid Signaling via Overexpression of Tomato (Solanum lycopersicum) SlBRI1 Improves Major Agronomic Traits. Front. Plant Sci. 2017, 8, 1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Zheng, C.; Zhao, Y.; Li, Q.; Liu, J.; Deng, R.; Lei, T.; Wang, S.; Wang, X. RNA Interference Knockdown of the Brassinosteroid Receptor BRI1 in Potato (Solanum tuberosum L.) Reveals Novel Functions for Brassinosteroid Signaling in Controlling Tuberization. Sci. Hortic. 2021, 290, 110516. [Google Scholar] [CrossRef]
- Costa, A.T.; Bravo, J.P.; Krause-Sakate, R.; Maia, I.G. The Receptor-like Kinase SlSOBIR1 Is Differentially Modulated by Virus Infection but Its Overexpression in Tobacco Has No Significant Impact on Virus Accumulation. Plant Cell Rep. 2016, 35, 35–75. [Google Scholar] [CrossRef]
- Huang, W.R.H.; Schol, C.; Villanueva, S.L.; Heidstra, R.; Joosten, M.H.A.J. Knocking out SOBIR1 in Nicotiana benthamiana Abolishes Functionality of Transgenic Receptor-like Protein Cf-4. Plant Physiol. 2021, 185, 290–294. [Google Scholar] [CrossRef]
- Zhou, Y.; Sun, L.; Wassan, G.M.; He, X.; Shaban, M.; Zhang, L.; Zhu, L.; Zhang, X. GbSOBIR1 Confers Verticillium Wilt Resistance by Phosphorylating the Transcriptional Factor GbbHLH171 in Gossypium barbadense. Plant Biotechnol. J. 2019, 17, 152–163. [Google Scholar] [CrossRef] [Green Version]
- Zipfel, C.; Kunze, G.; Chinchilla, D.; Caniard, A.; Jones, J.D.G.; Boller, T.; Felix, G. Perception of the Bacterial PAMP EF-Tu by the Receptor EFR Restricts Agrobacterium-Mediated Transformation. Cell 2006, 125, 749–760. [Google Scholar] [CrossRef]
- Lacombe, S.; Rougon-Cardoso, A.; Sherwood, E.; Peeters, N.; Dahlbeck, D.; Van Esse, H.P.; Smoker, M.; Rallapalli, G.; Thomma, B.P.H.J.; Staskawicz, B.; et al. Interfamily Transfer of a Plant Pattern-Recognition Receptor Confers Broad-Spectrum Bacterial Resistance. Nat. Biotechnol. 2010, 28, 365–369. [Google Scholar] [CrossRef]
- Holton, N.; Nekrasov, V.; Ronald, P.C.; Zipfel, C. The Phylogenetically-Related Pattern Recognition Receptors EFR and XA21 Recruit Similar Immune Signaling Components in Monocots and Dicots. PLoS Pathog. 2015, 11, e1004602. [Google Scholar] [CrossRef] [Green Version]
- Piazza, S.; Campa, M.; Pompili, V.; Costa, L.D.; Salvagnin, U.; Nekrasov, V.; Zipfel, C.; Malnoy, M. The Arabidopsis Pattern Recognition Receptor EFR Enhances Fire Blight Resistance in Apple. Hortic. Res. 2021, 8, 204. [Google Scholar] [CrossRef]
- Pfeilmeier, S.; George, J.; Morel, A.; Roy, S.; Smoker, M.; Stransfeld, L.; Downie, J.A.; Peeters, N.; Malone, J.G.; Zipfel, C. Expression of the Arabidopsis thaliana Immune Receptor EFR in Medicago truncatula Reduces Infection by a Root Pathogenic Bacterium, but Not Nitrogen-Fixing Rhizobial Symbiosis. Plant Biotechnol. J. 2019, 17, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Boschi, F.; Schvartzman, C.; Murchio, S.; Ferreira, V.; Siri, M.I.; Galván, G.A.; Smoker, M.; Stransfeld, L.; Zipfel, C.; Vilaró, F.L.; et al. Enhanced Bacterial Wilt Resistance in Potato through Expression of Arabidopsis Efr and Introgression of Quantitative Resistance from Solanum commersonii. Front. Plant Sci. 2017, 8, 1642. [Google Scholar] [CrossRef] [Green Version]
- Mosher, S.; Seybold, H.; Rodriguez, P.; Stahl, M.; Davies, K.A.; Dayaratne, S.; Morillo, S.A.; Wierzba, M.; Favery, B.; Keller, H.; et al. The Tyrosine-Sulfated Peptide Receptors PSKR1 and PSY1R Modify the Immunity of Arabidopsis to Biotrophic and Necrotrophic Pathogens in an Antagonistic Manner. Plant J. 2013, 73, 469–482. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, B.; Qi, G.; Shang, L.; Liu, H.; Ding, X.; Chu, Z. Identification of the Phytosulfokine Receptor 1 (OsPSKR1) Confers Resistance to Bacterial Leaf Streak in Rice. Planta 2019, 250, 1603–1612. [Google Scholar] [CrossRef]
- Nakaminami, K.; Okamoto, M.; Higuchi-Takeuchi, M.; Yoshizumi, T.; Yamaguchi, Y.; Fukao, Y.; Shimizu, M.; Ohashi, C.; Tanaka, M.; Matsui, M.; et al. AtPep3 Is a Hormone-like Peptide That Plays a Role in the Salinity Stress Tolerance of Plants. Proc. Natl. Acad. Sci. USA 2018, 115, 5810–5815. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Kang, S.; Jing, Y.; Ren, Z.; Li, L.; Zhou, J.M.; Berkowitz, G.; Shi, J.; Fu, A.; Lan, W.; et al. Danger-Associated Peptides Close Stomata by OST1-Independent Activation of Anion Channels in Guard Cells. Plant Cell 2018, 30, 1132–1146. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Liao, C.J.; Jaiswal, N.; Lee, S.; Yun, D.J.; Lee, S.Y.; Garvey, M.; Kaplan, I.; Mengiste, T. Tomato PEPR1 ORTHOLOG RECEPTOR-LIKE KINASE1 Regulates Responses to Systemin, Necrotrophic Fungi, and Insect Herbivory. Plant Cell 2018, 30, 2214–2229. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Yang, D.; Ha, Y.; Nam, K.H. BAK1-Induced RPK1 Phosphorylation Is Essential for RPK1-Mediated Cell Death in Arabidopsis. Biochem. Biophys. Res. Commun. 2021, 573, 125–131. [Google Scholar] [CrossRef]
- Choi, D.S.; Hwang, I.S.; Hwang, B.K. Requirement of the Cytosolic Interaction between PATHOGENESIS-RELATED PROTEIN10 and LEUCINE-RICH REPEAT PROTEIN1 for Cell Death and Defense Signaling in Pepper. Plant Cell 2012, 24, 1675–1690. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, T.; Rehman, A.U.; Wang, Y.; Qi, J.; Li, Z.; Song, C.; Wang, B.; Yang, S.; Gong, Z. Arabidopsis U-Box E3 Ubiquitin Ligase PUB11 Negatively Regulates Drought Tolerance by Degrading the Receptor-like Protein Kinases LRR1 and KIN7. J. Integr. Plant Biol. 2021, 63, 494–509. [Google Scholar] [CrossRef]
- Isner, J.C.; Begum, A.; Nuehse, T.; Hetherington, A.M.; Maathuis, F.J.M. KIN7 Kinase Regulates the Vacuolar TPK1 K+ Channel during Stomatal Closure. Curr. Biol. 2018, 28, 466–472.e4. [Google Scholar] [CrossRef] [Green Version]
- Thapa, G.; Gunupuru, L.R.; Hehir, J.G.; Kahla, A.; Mullins, E.; Doohan, F.M. A Pathogen-Responsive Leucine Rich Receptor like Kinase Contributes to Fusarium Resistance in Cereals. Front. Plant Sci. 2018, 9, 867. [Google Scholar] [CrossRef] [Green Version]
- Caddell, D.F.; Park, C.J.; Thomas, N.C.; Canlas, P.E.; Ronald, P.C. Silencing of the Rice Gene LRR1 Compromises Rice Xa21 Transcript Accumulation and XA21-Mediated Immunity. Rice 2017, 10, 23. [Google Scholar] [CrossRef] [Green Version]
- ten Hove, C.A.; de Jong, M.; Lapin, D.; Andel, A.; Sanchez-Perez, G.F.; Tarutani, Y.; Suzuki, Y.; Heidstra, R.; van den Ackerveken, G. Trans-Repression of Gene Activity Upstream of T-DNA Tagged Rlk902 Links Arabidopsis Root Growth Inhibition and Downy Mildew Resistance. PLoS ONE 2011, 6, e19028. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wu, G.; Shi, H.; Tang, D. RECEPTOR-LIKE KINASE 902 Associates with and Phosphorylates BRASSINOSTEROID-SIGNALING KINASE1 to Regulate Plant Immunity. Mol. Plant 2019, 12, 59–70. [Google Scholar] [CrossRef] [Green Version]
- Fontes, E.P.B.; Santos, A.A.; Luz, D.F.; Waclawovsky, A.J.; Chory, J. The Geminivirus Nuclear Shuttle Protein Is a Virulence Factor That Suppresses Transmembrane Receptor Kinase Activity. Genes Dev. 2004, 18, 2545–2556. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Weiberg, A.; Dellota, E.; Yamane, D.; Jin, H. Botrytis Small RNA Bc-SiR37 Suppresses Plant Defense Genes by Cross-Kingdom RNAi. RNA Biol. 2017, 14, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Jiang, G.Q.; Yao, X.F.; Liu, C.M. The Leucine-Rich Repeat Receptor-like Kinase OsERL Plays a Critical Role in Anther Lobe Formation in Rice. Biochem. Biophys. Res. Commun. 2021, 563, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Nanda, A.K.; El Habti, A.; Hocart, C.H.; Masle, J. ERECTA Receptor-Kinases Play a Key Role in the Appropriate Timing of Seed Germination under Changing Salinity. J. Exp. Bot. 2019, 70, 6417–6435. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, D.; Xu, Y.; Jin, S.; Zhang, L.; Zhang, S.; Yang, G.; Huang, J.; Yan, K.; Wu, C.; et al. CEPR2 Phosphorylates and Accelerates the Degradation of PYR/PYLs in Arabidopsis. J. Exp. Bot. 2019, 70, 5457–5469. [Google Scholar] [CrossRef]
- Jung, C.G.; Hwang, S.G.; Park, Y.C.; Park, H.M.; Kim, D.S.; Park, D.H.; Jang, C.S. Molecular Characterization of the Cold- and Heat-Induced Arabidopsis PXL1 Gene and Its Potential Role in Transduction Pathways under Temperature Fluctuations. J. Plant Physiol. 2015, 176, 138–146. [Google Scholar] [CrossRef]
- Pfister, A.; Barberon, M.; Alassimone, J.; Kalmbach, L.; Lee, Y.; Vermeer, J.E.M.; Yamazaki, M.; Li, G.; Maurel, C.; Takano, J.; et al. A Receptor-like Kinase Mutant with Absent Endodermal Diffusion Barrier Displays Selective Nutrient Homeostasis Defects. elife 2014, 3, e03115. [Google Scholar] [CrossRef]
- Tunc-Ozdemir, M.; Jones, A.M. BRL3 and AtRGS1 Cooperate to Fine Tune Growth Inhibition and ROS Activation. PLoS ONE 2017, 12, e0177400. [Google Scholar] [CrossRef] [Green Version]
- Fàbregas, N.; Lozano-Elena, F.; Blasco-Escámez, D.; Tohge, T.; Martínez-Andújar, C.; Albacete, A.; Osorio, S.; Bustamante, M.; Riechmann, J.L.; Nomura, T.; et al. Overexpression of the Vascular Brassinosteroid Receptor BRL3 Confers Drought Resistance without Penalizing Plant Growth. Nat. Commun. 2018, 9, 4680. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, A.; Fujioka, S.; Sunohara, H.; Kamiya, N.; Hong, Z.; Inukai, Y.; Miura, K.; Takatsuto, S.; Yoshida, S.; Ueguchi-Tanaka, M.; et al. The Role of OsBRI1 and Its Homologous Genes, OsBRL1 and OsBRL3, in Rice. Plant Physiol. 2006, 140, 580–590. [Google Scholar] [CrossRef] [Green Version]
- Pitorre, D.; Llauro, C.; Jobet, E.; Guilleminot, J.; Brizard, J.P.; Delseny, M.; Lasserre, E. RLK7, a Leucine-Rich Repeat Receptor-like Kinase, Is Required for Proper Germination Speed and Tolerance to Oxidative Stress in Arabidopsis thaliana. Planta 2010, 232, 1339–1353. [Google Scholar] [CrossRef]
- Replogle, A.; Wang, J.; Paolillo, V.; Smeda, J.; Kinoshita, A.; Durbak, A.; Tax, F.E.; Wang, X.; Sawa, S.; Mitchum, M.G. Synergistic Interaction of Clavata1, Clavata2, and Receptor-like Protein Kinase 2 in Cyst Nematode Parasitism of Arabidopsis. Mol. Plant-Microbe Interact. 2013, 26, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.; Zhu, Y.; Cui, Y.; Cheng, K.; Liang, W.; Wei, Z.; Zhu, M.; Yin, H.; Zeng, L.; Xiao, Y.; et al. A Group of Receptor Kinases Are Essential for CLAVATA Signalling to Maintain Stem Cell Homeostasis. Nat. Plants 2018, 4, 205–211. [Google Scholar] [CrossRef]
- Xu, F.; Meng, T.; Li, P.; Yu, Y.; Cui, Y.; Wang, Y.; Gong, Q.; Wang, N.N. A Soybean Dual-Specificity Kinase, GmSARK, and Its Arabidopsis Homolog, AtSARK, Regulate Leaf Senescence through Synergistic Actions of Auxin and Ethylene. Plant Physiol. 2011, 157, 2131–2153. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Yang, H.; Liu, G.; Ma, W.; Li, C.; Huo, H.; He, J.; Liu, L. PpSARK Regulates Moss Senescence and Salt Tolerance through ABA Related Pathway. Int. J. Mol. Sci. 2018, 19, 2609. [Google Scholar] [CrossRef]
- da Silva, H.A.P.; Caetano, V.S.; Pessoa, D.D.V.; Pacheco, R.S.; Simoes-Araujo, J.L. Molecular and Biochemical Changes of Aging-Induced Nodules Senescence in Common Bean. Symbiosis 2019, 79, 33–48. [Google Scholar] [CrossRef]
- Soltabayeva, A.; Ongaltay, A.; Omondi, J.O.; Srivastava, S. Morphological, Physiological and Molecular Markers for Salt-Stressed Plants. Plants 2021, 10, 243. [Google Scholar] [CrossRef]
- Meng, X.; Zhou, J.; Tang, J.; Li, B.; de Oliveira, M.V.V.; Chai, J.; He, P.; Shan, L. Ligand-Induced Receptor-like Kinase Complex Regulates Floral Organ Abscission in Arabidopsis. Cell Rep. 2016, 14, 1330–1338. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, Y.; Huang, J.; Ahsan, N.; Biener, G.; Paprocki, J.; Thelen, J.J.; Raicu, V.; Zhao, D. Two SERK Receptor-like Kinases Interact with EMS1 to Control Anther Cell Fate Determination. Plant Physiol. 2017, 173, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Wu, S.; Gao, X.; Zhang, Y.; Shan, L.; He, P. A Receptor-like Cytoplasmic Kinase, BIK1, Associates with a Flagellin Receptor Complex to Initiate Plant Innate Immunity. Proc. Natl. Acad. Sci. USA 2010, 107, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Li, M.; Yu, L.; Zhou, Z.; Liang, X.; Liu, Z.; Cai, G.; Gao, L.; Zhang, X.; Wang, Y.; et al. The FLS2-Associated Kinase BIK1 Directly Phosphorylates the NADPH Oxidase RbohD to Control Plant Immunity. Cell Host Microbe 2014, 15, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Wang, Y.; Li, R.; Song, X.; Wang, Q.; Huang, S.; Jin, J.B.; Liu, C.M.; Lin, J. Analysis of Interactions among the CLAVATA3 Receptors Reveals a Direct Interaction between CLAVATA2 and CORYNE in Arabidopsis. Plant J. 2010, 61, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Schulze, B.; Mentzel, T.; Jehle, A.K.; Mueller, K.; Beeler, S.; Boller, T.; Felix, G.; Chinchilla, D. Rapid Heteromerization and Phosphorylation of Ligand-Activated Plant Transmembrane Receptors and Their Associated Kinase BAK1. J. Biol. Chem. 2010, 285, 9444–9451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amorim-Silva, V.; García-Moreno, Á.; Castillo, A.G.; Lakhssassi, N.; Del Valle, A.E.; Pérez-Sancho, J.; Li, Y.; Posé, D.; Pérez-Rodriguez, J.; Lin, J.; et al. TTL Proteins Scaffold Brassinosteroid Signaling Components at the Plasma Membrane to Optimize Signal Transduction in Arabidopsis. Plant Cell 2019, 31, 1807–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago, J.; Henzler, C.; Hothorn, M. Molecular Mechanism for Plant Steroid Receptor Activation by Somatic Embryogenesis Co-Receptor Kinases. Science 2013, 341, 889–892. [Google Scholar] [CrossRef]
- Nicaise, V.; Joe, A.; Jeong, B.R.; Korneli, C.; Boutrot, F.; Westedt, I.; Staiger, D.; Alfano, J.R.; Zipfel, C. Pseudomonas HopU1 Modulates Plant Immune Receptor Levels by Blocking the Interaction of Their MRNAs with GRP7. EMBO J. 2013, 32, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, H.; Han, Z.; Zhang, H.; Wang, T.; Lin, G.; Chang, J.; Yang, W.; Chai, J. Allosteric Receptor Activation by the Plant Peptide Hormone Phytosulfokine. Nature 2015, 525, 265–268. [Google Scholar] [CrossRef]
- Kinoshita, A.; Betsuyaku, S.; Osakabe, Y.; Mizuno, S.; Nagawa, S.; Stahl, Y.; Simon, R.; Yamaguchi-Shinozaki, K.; Fukuda, H.; Sawa, S. RPK2 Is an Essential Receptor-like Kinase That Transmits the CLV3 Signal in Arabidopsis. Development 2010, 137, 3911–3920. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Han, L.; Hymes, M.; Denver, R.; Clark, S.E. CLAVATA2 Forms a Distinct CLE-Binding Receptor Complex Regulating Arabidopsis Stem Cell Specification. Plant J. 2010, 573, 125–131. [Google Scholar] [CrossRef] [Green Version]
- He, K.; Gou, X.; Yuan, T.; Lin, H.; Asami, T.; Yoshida, S.; Russell, S.D.; Li, J. BAK1 and BKK1 Regulate Brassinosteroid-Dependent Growth and Brassinosteroid-Independent Cell-Death Pathways. Curr. Biol. 2007, 17, 1109–1115. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Chen, X.; Mang, H.; Liu, C.; Yu, X.; Gao, X.; Torii, K.U.; He, P.; Shan, L. Differential Function of Arabidopsis SERK Family Receptor-like Kinases in Stomatal Patterning. Curr. Biol. 2015, 25, 2361–2372. [Google Scholar] [CrossRef] [Green Version]
- Mou, S.; Zhang, X.; Han, Z.; Wang, J.; Gong, X.; Chai, J. CLE42 Binding Induces PXL2 Interaction with SERK2. Protein Cell 2017, 8, 612–617. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Kota, U.; He, K.; Blackburn, K.; Li, J.; Goshe, M.B.; Huber, S.C.; Clouse, S.D. Sequential Transphosphorylation of the BRI1/BAK1 Receptor Kinase Complex Impacts Early Events in Brassinosteroid Signaling. Dev. Cell 2008, 15, 220–235. [Google Scholar] [CrossRef] [Green Version]
- Franco-Orozco, B.; Berepiki, A.; Ruiz, O.; Gamble, L.; Griffe, L.L.; Wang, S.; Birch, P.R.J.; Kanyuka, K.; Avrova, A. A New Proteinaceous Pathogen-Associated Molecular Pattern (PAMP) Identified in Ascomycete Fungi Induces Cell Death in Solanaceae. New Phytol. 2017, 214, 1657–1672. [Google Scholar] [CrossRef] [Green Version]
- Crook, A.D.; Schnabel, E.L.; Frugoli, J.A. The Systemic Nodule Number Regulation Kinase SUNN in Medicago truncatula Interacts with MtCLV2 and MtCRN. Plant J. 2016, 88, 108–119. [Google Scholar] [CrossRef]
- Smakowska-Luzan, E.; Mott, G.A.; Parys, K.; Stegmann, M.; Howton, T.C.; Layeghifard, M.; Neuhold, J.; Lehner, A.; Kong, J.; Grünwald, K.; et al. An Extracellular Network of Arabidopsis Leucine-Rich Repeat Receptor Kinases. Nature 2018, 553, 342–346. [Google Scholar] [CrossRef]
- Mott, G.A.; Smakowska-Luzan, E.; Pasha, A.; Parys, K.; Howton, T.C.; Neuhold, J.; Lehner, A.; Grünwald, K.; Stolt-Bergner, P.; Provart, N.J.; et al. Data Descriptor: Map of Physical Interactions between Extracellular Domains of Arabidopsis Leucine-Rich Repeat Receptor Kinases. Sci. Data 2019, 6, 190025. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Ferreira, M.A.; Huang, M.; Camargos, L.F.; Yu, X.; Teixeira, R.M.; Carpinetti, P.A.; Mendes, G.C.; Gouveia-Mageste, B.C.; Liu, C.; et al. The Receptor-like Kinase NIK1 Targets FLS2/BAK1 Immune Complex and Inversely Modulates Antiviral and Antibacterial Immunity. Nat. Commun. 2019, 10, 4996. [Google Scholar] [CrossRef] [Green Version]
- Karlova, R.; Boeren, S.; Russinova, E.; Aker, J.; Vervoort, J.; De Vries, S. The Arabidopsis SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASE1 Protein Complex Includes BRASSINOSTEROID-INSENSITIVE1. Plant Cell 2006, 18, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Li, L.; Macho, A.P.; Han, Z.; Hu, Z.; Zipfel, C.; Zhou, J.-M.; Chai, J. Structural Basis for Flg22-Induced Activation of the Arabidopsis FLS2-BAK1 Immune Complex. Science 2013, 342, 624–628. [Google Scholar] [CrossRef]
- He, Y.; Zhou, J.; Shan, L.; Meng, X. Plant Cell Surface Receptor-Mediated Signaling-A Common Theme amid Diversity. J. Cell Sci. 2018, 131, jcs209353. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, C.; Russinova, E.; Hecht, V.; Baaijens, E.; de Vries, S. The Arabidopsis thaliana SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASES1 and 2 Control Male Sporogenesis. Plant Cell 2005, 17, 3337–3349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, J.; Han, Z.; Sun, Y.; Zhang, H.; Gong, X.; Chai, J. Structural Basis for Recognition of an Endogenous Peptide by the Plant Receptor Kinase PEPR1. Cell Res. 2015, 25, 110–120. [Google Scholar] [CrossRef]
- Xiang, T.; Zong, N.; Zou, Y.; Wu, Y.; Zhang, J.; Xing, W.; Li, Y.; Tang, X.; Zhu, L.; Chai, J.; et al. Pseudomonas Syringae Effector AvrPto Blocks Innate Immunity by Targeting Receptor Kinases. Curr. Biol. 2008, 18, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Ranf, S.; Eschen-Lippold, L.; Pecher, P.; Lee, J.; Scheel, D. Interplay between Calcium Signalling and Early Signalling Elements during Defence Responses to Microbe- or Damage-Associated Molecular Patterns. Plant J. 2011, 68, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Warpeha, K.M.; Huber, S.C. The Brassinosteroid Receptor Kinase, BRI1, Plays a Role in Seed Germination and the Release of Dormancy by Cold Stratification. J. Plant Physiol. 2019, 241, 153031. [Google Scholar] [CrossRef] [PubMed]
- Kemmerling, B.; Halter, T.; Mazzotta, S.; Mosher, S.; Nürnberger, T. A Genome-Wide Survey for Arabidopsis Leucine-Rich Repeat Receptor Kinases Implicated in Plant Immunity. Front. Plant Sci. 2011, 2, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Jiang, N.; Liu, J.; Liu, W.; Wang, G.L. The Role of Effectors and Host Immunity in Plant–Necrotrophic Fungal Interactions. Virulence 2014, 5, 722–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loivamäki, M.; Stührwohldt, N.; Deeken, R.; Steffens, B.; Roitsch, T.; Hedrich, R.; Sauter, M. A Role for PSK Signaling in Wounding and Microbial Interactions in Arabidopsis. Physiol. Plant. 2010, 139, 348–357. [Google Scholar] [CrossRef]
- Furukawa, K.; Hoshi, Y.; Maeda, T.; Nakajima, T.; Abe, K. Aspergillus Nidulans HOG Pathway Is Activated Only by Two-Component Signalling Pathway in Response to Osmotic Stress. Mol. Microbiol. 2005, 56, 1246–1261. [Google Scholar] [CrossRef]
- Carvalho, C.M.; Santos, A.A.; Pires, S.R.; Rocha, C.S.; Saraiva, D.I.; Machado, J.P.B.; Mattos, E.C.; Fietto, L.G.; Fontes, E.P.B. Regulated Nuclear Trafficking of RpL10A Mediated by NIK1 Represents a Defense Strategy of Plant Cells against Virus. PLoS Pathog. 2008, 4, e1000247. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Yang, D.; Ha, Y.; Shin, H.Y.; Nam, K.H. Receptor-like Protein Kinases RPK1 and BAK1 Sequentially Form Complexes with the Cytoplasmic Kinase OST1 to Regulate ABA-Induced Stomatal Closure. J. Exp. Bot. 2020, 71, 1491–1502. [Google Scholar] [CrossRef]
- Osakabe, Y.; Mizuno, S.; Tanaka, H.; Maruyama, K.; Osakabe, K.; Todaka, D.; Fujita, Y.; Kobayashi, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Overproduction of the Membrane-Bound Receptor-like Protein Kinase 1, RPK1, Enhances Abiotic Stress Tolerance in Arabidopsis. J. Biol. Chem. 2010, 285, 9190–9201. [Google Scholar] [CrossRef] [Green Version]
- Schoonbeek, H.J.; Wang, H.H.; Stefanato, F.L.; Craze, M.; Bowden, S.; Wallington, E.; Zipfel, C.; Ridout, C.J. Arabidopsis EF-Tu Receptor Enhances Bacterial Disease Resistance in Transgenic Wheat. New Phytol. 2015, 206, 606–613. [Google Scholar] [CrossRef]
- Shen, H.; Zhong, X.; Zhao, F.; Wang, Y.; Yan, B.; Li, Q.; Chen, G.; Mao, B.; Wang, J.; Li, Y.; et al. Overexpression of Receptor-like Kinase ERECTA Improves Thermotolerance in Rice and Tomato. Nat. Biotechnol. 2015, 33, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Citerne, S.; Antelme, S.; Le Bris, P.; Daniel, S.; Bouder, A.; D’Orlando, A.; Cartwright, A.; Tellier, F.; Pateyron, S.; et al. BdERECTA Controls Vasculature Patterning and Phloem-Xylem Organization in Brachypodium Distachyon. BMC Plant Biol. 2021, 21, 196. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, S.; Xue, S.; Yang, S.; Huang, J.; Wang, L. Phylogenetic and CRISPR/Cas9 Studies in Deciphering the Evolutionary Trajectory and Phenotypic Impacts of Rice ERECTA Genes. Front. Plant Sci. 2018, 9, 473. [Google Scholar] [CrossRef] [PubMed]
- De Gruijl, F.R.; Rebel, H. Early Events in UV Carcinogenesis-DNA Damage, Target Cells and Mutant P53 Foci. Photochem. Photobiol. 2008, 84, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Liao, C.J.; Mengesha, B.; Han, H.; Lee, S.; Sharon, A.; Zhou, Y.; Mengiste, T. Regulation of Plant Immunity and Growth by Tomato Receptor-like Cytoplasmic Kinase TRK1. New Phytol. 2022, 233, 458–478. [Google Scholar] [CrossRef]
- Araya, T.; Miyamoto, M.; Wibowo, J.; Suzuki, A.; Kojima, S.; Tsuchiya, Y.N.; Sawa, S.; Fukuda, H.; Von Wirén, N.; Takahashi, H. CLE-CLAVATA1 Peptide-Receptor Signaling Module Regulates the Expansion of Plant Root Systems in a Nitrogen-Dependent Manner. Proc. Natl. Acad. Sci. USA 2014, 111, 2029–2034. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Xu, Y.Y.; Li, J.; Powell, R.A.; Xu, Z.H.; Chong, K. Transgenic Rice Plants Ectopically Expressing AtBAK1 Are Semi-Dwarfed and Hypersensitive to 24-Epibrassinolide. J. Plant Physiol. 2007, 164, 655–664. [Google Scholar] [CrossRef]
- Hu, L.; Ye, M.; Kuai, P.; Ye, M.; Erb, M.; Lou, Y. OsLRR-RLK1, an Early Responsive Leucine-Rich Repeat Receptor-like Kinase, Initiates Rice Defense Responses against a Chewing Herbivore. New Phytol. 2018, 219, 1097–1111. [Google Scholar] [CrossRef] [Green Version]
- Brotman, Y.; Landau, U.; Pnini, S.; Lisec, J.; Balazadeh, S.; Mueller-Roeber, B.; Zilberstein, A.; Willmitzer, L.; Chet, I.; Viterbo, A. The LysM Receptor-like Kinase LysM RLK1 Is Required to Activate Defense and Abiotic-Stress Responses Induced by Overexpression of Fungal Chitinases in Arabidopsis Plants. Mol. Plant 2012, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Cao, B.; Kang, J.; Wang, X.; Han, X.; Jiang, W.; Shi, X.; Zhang, L.; Cui, L.; Hu, Z.; et al. Fine-Tuning Stomatal Movement through Small Signaling Peptides. Front. Plant Sci. 2019, 10, 69. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.S.; Gunupuru, L.R.; Kumar, G.B.S.; Khan, M.; Scofield, S.; Nicholson, P.; Doohan, F.M. Plant Disease Resistance Is Augmented in Uzu Barley Lines Modified in the Brassinosteroid Receptor BRI1. BMC Plant Biol. 2014, 14, 227. [Google Scholar] [CrossRef] [PubMed]

| Numbers of LRR-RLK | Plant Species | Genome Size | Presence of Tandem Duplication | LRR-RLK Organizations | References |
|---|---|---|---|---|---|
| 0 | Chlamydomonas reinhardtii | 120 Mb | - | - | [34] |
| 60 | Sedum alfredii | 39.1 Mb | - | motif conservation | [35] |
| 67–81 | Selaginella moellendorffii | 100 Mb | present | motif conservation | [34] |
| 94 | Amborella trichopoda | 870 Mb | present | motif conservation | [36] |
| 134 | Physcomitrella patens | 500 Mb | - | [34,37] | |
| 176 | Phoenix dactylifera | 658 Mb | present | - | [37,38] |
| 180 | Cucumis sativus | 367 Mb | present | segmental duplication | [37,39] |
| 201 | Fragaria vesca | 240 Mb | present | - | [40] |
| 211 | Medicago truncatula | 465 Mb | present | exon/intron organization, motif conservation | [37,41] |
| 215 | Vitis vinifera | 500 Mb | present | segmental duplication | [37,42] |
| 226 | Arabidopsis thaliana | 133 Mb | present | - | [37,43] |
| 227 | Jatropha curcas | 320 Mb | present | - | [37,44] |
| 230–236 | Vernicia fordii Vernicia montana | 1310 Mb | Present (V. fordii) | motif conservation (both), segmental duplications (V. fordii) | [45,46] |
| 239 | Solanum lycopersicum | 900 Mb | present | - | [37,47] |
| 247 | Theobroma cacao | 430 Mb | present | motif conservation | [37,48] |
| 250 | Zea mays | 2400 Mb | present | random chromosomal distribution | [37,49] |
| 267 | Prunus mume | 280 Mb | present | - | [40] |
| 268 | Solanum tuberosum | 840 Mb | present | lineage-specific expansion | [37,50] |
| 268 | Prunus persica | 265 Mb | present | - | [37,40] |
| 292 | Raphanus sativus | 574 Mb | present | motif conservation | [51] |
| 298 | Gossypium arboreum | 1750 Mb | present | conserved exon/intron organization | [52] |
| 297–300 | Citrus clementina Citrus sinensis | 370 Mb 380 Mb | present | - | [53] |
| 300 | Brassica rapa | 455 Mb | present | intron/exon pattern organization, motif conservation | [37,54] |
| 310 | Setaria italica | 515 Mb | present | motif conservation, segmental duplication | [37,55] |
| 312 | Brassica nigra | 522 Mb | present | intra-chromosomal duplication, conserved loci duplication | [56] |
| 332 | Oryza sativa | 430 Mb | present | exon duplication, mutation, and exon shuffling | [37,57] |
| 367 | Glycine latifolia | 939 Mb | present | - | [58] |
| 384 | Gossypium raimondii | 761 Mb | present | motif conservation | [37,59] |
| 427 | Pyrus bretschneideri | 512 Mb | present | exon/intron organization, motif conservation | [60] |
| 441 | Populus trichocarpa | 500 Mb | present | segmental duplication, exon/intron organization and motif conservation | [37,61] |
| 484 | Brassica juncea | 920 Mb | present | segmental duplications, intra-/inter-genomic duplications | [62] |
| 485 | Malus domestica | 750 Mb | present | - | [37,40] |
| 494 | Glycine max | 1100–1150 Mb | present | exon/intron organizations, motif arrangements | [37,63] |
| 531 | Triticum aestivum | 17,000 Mb | present | segmental duplications, intra-genomic duplications | [64] |
| 548 | Arachis hypogaea | 2700 Mb | present | segmental duplication, exon/intron organization and motif conservation | [65] |
| 589 | Thinopyrum elongatum | 4780 Mb | present | segmental duplications | [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soltabayeva, A.; Dauletova, N.; Serik, S.; Sandybek, M.; Omondi, J.O.; Kurmanbayeva, A.; Srivastava, S. Receptor-like Kinases (LRR-RLKs) in Response of Plants to Biotic and Abiotic Stresses. Plants 2022, 11, 2660. https://doi.org/10.3390/plants11192660
Soltabayeva A, Dauletova N, Serik S, Sandybek M, Omondi JO, Kurmanbayeva A, Srivastava S. Receptor-like Kinases (LRR-RLKs) in Response of Plants to Biotic and Abiotic Stresses. Plants. 2022; 11(19):2660. https://doi.org/10.3390/plants11192660
Chicago/Turabian StyleSoltabayeva, Aigerim, Nurbanu Dauletova, Symbat Serik, Margulan Sandybek, John Okoth Omondi, Assylay Kurmanbayeva, and Sudhakar Srivastava. 2022. "Receptor-like Kinases (LRR-RLKs) in Response of Plants to Biotic and Abiotic Stresses" Plants 11, no. 19: 2660. https://doi.org/10.3390/plants11192660