
Parallel Differentiation and Plastic Adjustment of Leaf Anatomy in Alpine Arabidopsis arenosa Ecotypes
 , , , , and
, , , , and 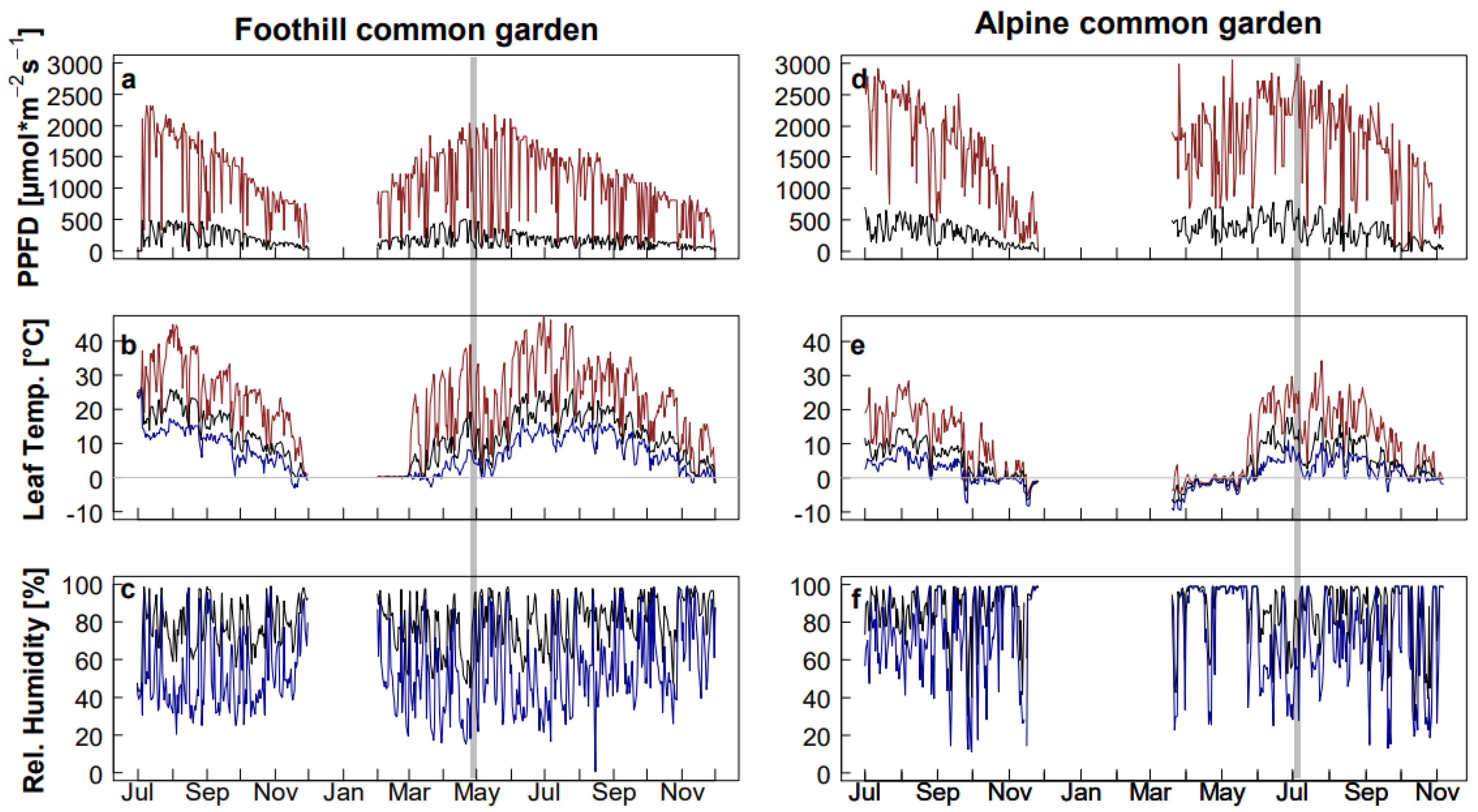{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Environmental Conditions in the Common Gardens Used for Reciprocal Transplantation
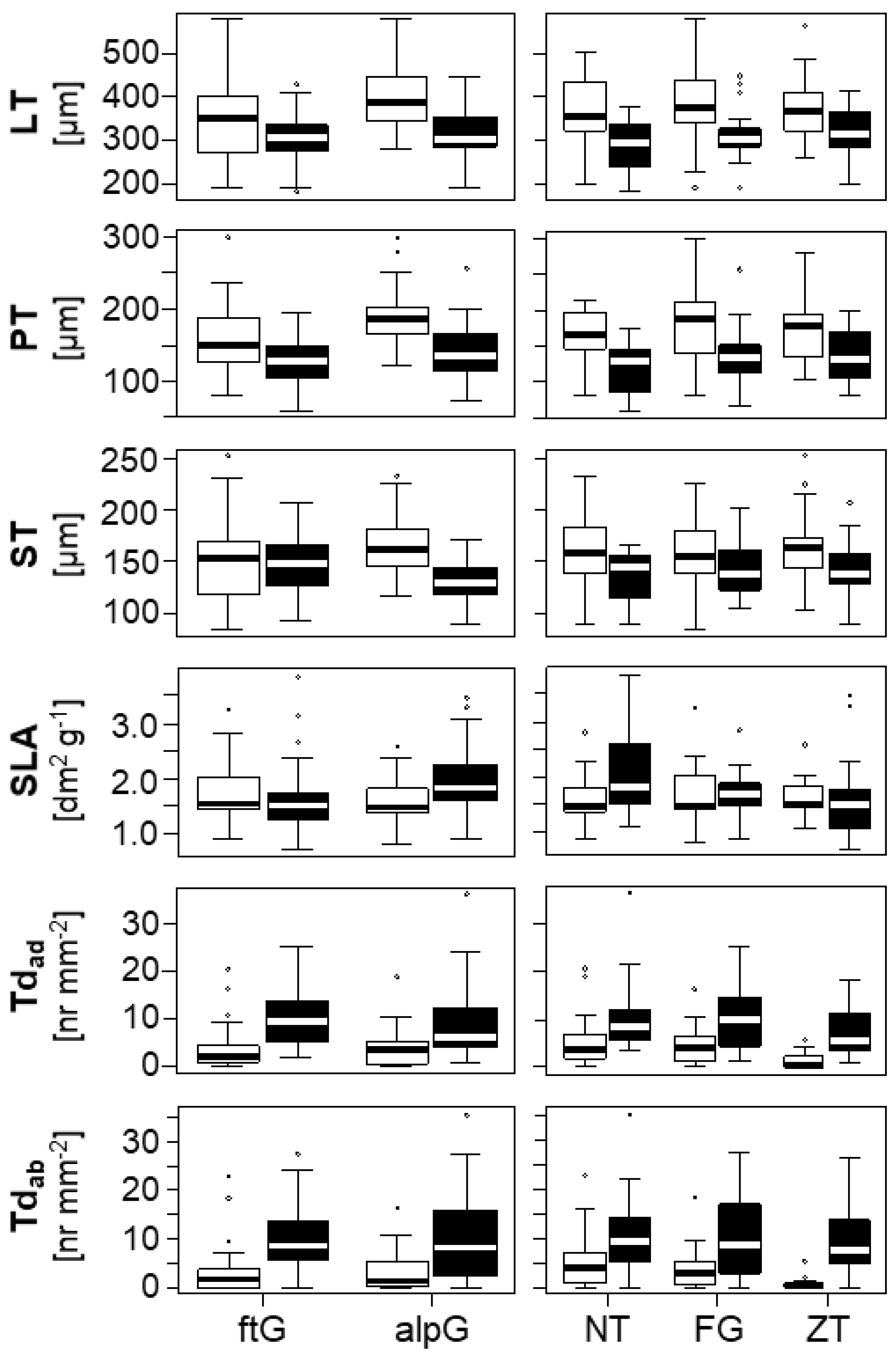
2.2. Differences in Leaf Traits of Ecotypes Grown in the Alpine and Foothill Common Gardens
2.3. Ecotype Plasticity
2.4. The Effect of Ploidy on Leaf Traits
2.5. Chloroplast Ultrastructure
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Biological Material
4.2. Microclimate
4.3. Sampling and Leaf Anatomy
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Körner, C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems, 2nd ed.; Springer: Heidelberg/Berlin, Germany, 2003; ISBN 978-3-642-18970-8. [Google Scholar]
- Larcher, W. Ökophysiologie der Pflanzen. Leben, Leistung und Streßbewältigung der Pflanzen in ihrer Umwelt; Ulmer: Stuttgart, Germany, 2001; Volume 6, ISBN 3825280748. [Google Scholar]
- Sakai, A.; Larcher, W. Low temperature and frost as environmental factors. In Frost Survival of Plants; Ecological Studies; Springer: Heidelberg/Berlin, Germany, 1987; Volume 62, pp. 1–20. ISBN 978-3-642-71747-5. [Google Scholar]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Ackerly, D.D.; Reich, P.B. Convergence and correlations among leaf size and function in seed plants: A comparative test using independent contrasts. Am. J. Bot. 1999, 86, 1272–1281. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.-F.; Li, S.-F.; Li, S.-F.; Xie, W.-J.; Song, J. How do leaf anatomies and photosynthesis of three Rhododendron species relate to their natural environments? Bot. Stud. 2014, 55, 36. [Google Scholar] [CrossRef] [Green Version]
- Rôças, G.; Barros, C.F.; Scarano, F.R. Leaf anatomy plasticity of Alchornea triplinervia (Euphorbiaceae) under distinct light regimes in a Brazilian montane Atlantic rain forest. Trees Struct. Funct. 1997, 11, 469–473. [Google Scholar] [CrossRef]
- Körner, C. The nutritional status of plants from high altitudes: A worldwide comparison. Oecologia 1989, 81, 379–391. [Google Scholar] [CrossRef]
- Morecroft, M.D.; Woodward, F.I. Experiments on the causes of altitudinal differences in the leaf nutrient contents, size and δ13C of Alchemilla alpina. New Phytol. 1996, 134, 471–479. [Google Scholar] [CrossRef]
- Kammer, P.M.; Steiner, J.S.; Schöb, C. Arabis alpina and Arabidopsis thaliana have different stomatal development strategies in response to high altitude pressure conditions. Alp. Bot. 2015, 125, 101–112. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Beck, E.; Müller-Hohenstein, K. Pflanzenökologie; Spektrum Akademischer Verlag: Heidelberg, Germany, 2002; ISBN 382740987X. [Google Scholar]
- Lösch, R. Wasserhaushalt der Pflanzen, 1st ed.; Quelle & Meyer: Wiebelsheim, Germany, 2001; ISBN 978-3-8252-8141-0. [Google Scholar]
- Chartzoulakis, K.; Patakas, A.; Kofidis, G.; Bosabalidis, A.; Nastou, A. Water stress affects leaf anatomy, gas exchange, water relations and growth of two avocado cultivars. Sci. Hortic. 2002, 95, 39–50. [Google Scholar] [CrossRef]
- Rury, P.M.; Dickison, W.C. Structural correlations among wood, leaves and plant habit. In Contemporary Problems in Plant Anatomy; White, R.A., Dickison, W.C., Eds.; Academic Press: San Diego, CA, USA, 1984; pp. 495–540. ISBN 978-0-12-746620-0. [Google Scholar]
- Scoffoni, C.; Chatelet, D.S.; Pasquet-Kok, J.; Rawls, M.; Donoghue, M.J.; Edwards, E.J.; Sack, L. Hydraulic basis for the evolution of photosynthetic productivity. Nat. Plants 2016, 2, 16072. [Google Scholar] [CrossRef]
- Bohutínská, M.; Vlček, J.; Yair, S.; Laenen, B.; Konečná, V.; Fracassetti, M.; Slotte, T.; Kolář, F. Genomic basis of parallel adaptation varies with divergence in Arabidopsis and its relatives. Proc. Natl. Acad. Sci. USA 2021, 118, e2022713118. [Google Scholar] [CrossRef]
- Pfennig, D.W.; Wund, M.A.; Snell-Rood, E.C.; Cruickshank, T.; Schlichting, C.D.; Moczek, A.P. Phenotypic plasticity’s impacts on diversification and speciation. Trends Ecol. Evol. 2010, 25, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Bonduriansky, R.; Crean, A.J.; Day, T. The implications of nongenetic inheritance for evolution in changing environments. Evol. Appl. 2012, 5, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Hufford, K.M.; Mazer, S.J. Plant ecotypes: Genetic differentiation in the age of ecological restoration. Trends Ecol. Evol. 2003, 18, 147–155. [Google Scholar] [CrossRef]
- Lowry, D.B. Ecotypes and the controversy over stages in the formation of new species. Biol. J. Linn. Soc. 2012, 106, 241–257. [Google Scholar] [CrossRef] [Green Version]
- Elmer, K.R.; Meyer, A. Adaptation in the age of ecological genomics: Insights from parallelism and convergence. Trends Ecol. Evol. 2011, 26, 298–306. [Google Scholar] [CrossRef] [Green Version]
- Barrett, R.D.; Schluter, D. Adaptation from standing genetic variation. Trends Ecol. Evol. 2008, 23, 38–44. [Google Scholar] [CrossRef]
- Preite, V.; Sailer, C.; Syllwasschy, L.; Bray, S.; Ahmadi, H.; Kramer, U.; Yant, L. Convergent evolution in Arabidopsis halleri and Arabidopsis arenosa on calamine metalliferous soils. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180243. [Google Scholar] [CrossRef] [Green Version]
- Trucchi, E.; Frajman, B.; Haverkamp, T.H.A.; Schönswetter, P.; Paun, O. Genomic analyses suggest parallel ecological divergence in Heliosperma pusillum (Caryophyllaceae). New Phytol. 2017, 216, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Knotek, A.; Konečná, V.; Wos, G.; Požárová, D.; Šrámková, G.; Bohutínská, M.; Zeisek, V.; Marhold, K.; Kolář, F. Parallel alpine differentiation in Arabidopsis arenosa. Front. Plant Sci. 2020, 11, 561526. [Google Scholar] [CrossRef]
- Antonovics, J. The nature of limits to natural selection. Ann. Mo. Bot. Gard. 1976, 63, 224–247. [Google Scholar] [CrossRef]
- Kawecki, T.J.; Ebert, D. Conceptual issues in local adaptation. Ecol. Lett. 2004, 7, 1225–1241. [Google Scholar] [CrossRef] [Green Version]
- Szukala, A.; Lovegrove-Walsh, J.; Luqman, H.; Fior, S.; Wolfe, T.M.; Frajman, B.; Schönswetter, P.; Paun, O. Polygenic routes lead to parallel altitudinal adaptation in Heliosperma pusillum (Caryophyllaceae). Mol. Ecol. 2022, 1–16. [Google Scholar] [CrossRef]
- van Kleunen, M.; Fischer, M. Constraints on the evolution of adaptive phenotypic plasticity in plants. New Phytol. 2005, 166, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Brochmann, C.; Borgen, L.; Stabbetorp, O.E. Multiple diploid hybrid speciation of the Canary Island endemic Argyranthemum sundingii (Asteraceae). Plant Syst. Evol. 2000, 220, 77–92. [Google Scholar] [CrossRef]
- Berglund, A.B.N.; Dahlgren, S.; Westerbergh, A. Evidence for parallel evolution and site-specific selection of serpentine tolerance in Cerastium alpinum during the colonization of Scandinavia. New Phytol. 2004, 161, 199–209. [Google Scholar] [CrossRef]
- Foster, S.A.; McKinnon, G.E.; Steane, D.A.; Potts, B.M.; Vaillancourt, R.E. Parallel evolution of dwarf ecotypes in the forest tree Eucalyptus globulus. New Phytol. 2007, 175, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Roda, F.; Ambrose, L.; Walter, G.M.; Liu, H.L.; Schaul, A.; Lowe, A.; Pelser, P.B.; Prentis, P.; Rieseberg, L.H.; Ortiz-Barrientos, D. Genomic evidence for the parallel evolution of coastal forms in the Senecio lautus complex. Mol. Ecol. 2013, 22, 2941–2952. [Google Scholar] [CrossRef]
- Schat, H.; Vooijs, R.; Kuiper, E. Identical major gene loci for heavy metal tolerances that have independently evolved in different local populations and subspecies of Silene vulgaris. Evolution 1996, 50, 1888–1895. [Google Scholar] [CrossRef]
- Kolář, F.; Fuxová, G.; Záveská, E.; Nagano, A.J.; Hyklová, L.; Lučanová, M.; Kudoh, H.; Marhold, K. Northern glacial refugia and altitudinal niche divergence shape genome-wide differentiation in the emerging plant model Arabidopsis arenosa. Mol. Ecol. 2016, 25, 3929–3949. [Google Scholar] [CrossRef]
- Kolář, F.; Lučanová, M.; Záveská, E.; Fuxová, G.; Mandáková, T.; Španiel, S.; Senko, D.; Svitok, M.; Kolník, M.; Gudžinskas, Z.; et al. Ecological segregation does not drive the intricate parapatric distribution of diploid and tetraploid cytotypes of the Arabidopsis arenosa group (Brassicaceae). Biol. J. Linn. Soc. 2016, 119, 673–688. [Google Scholar] [CrossRef] [Green Version]
- Wos, G.; Arc, E.; Hülber, K.; Konečná, V.; Knotek, A.; Požárová, D.; Bertel, C.; Kaplenig, D.; Mandáková, T.; Neuner, G.; et al. Parallel local adaptation to an alpine environment in Arabidopsis arenosa. J. Ecol. 2022. [Google Scholar] [CrossRef]
- Geng, Y.P.; Pan, X.Y.; Xu, C.Y.; Zhang, W.J.; Li, B.; Chen, J.K.; Lu, B.R.; Song, Z.P. Phenotypic plasticity rather than locally adapted ecotypes allows the invasive alligator weed to colonize a wide range of habitats. Biol. Invasions 2007, 9, 245–256. [Google Scholar] [CrossRef]
- Mešícĕk, J.; Goliašová, K. Cardaminopsis (C. A. Mey.) Hayek. In Flóra Slovenska; Goliašová, K., Šípošová, H., Eds.; Veda: Bratislava, Slovakia, 2002; pp. 388–415. ISBN 9788022407106. [Google Scholar]
- Körner, C.; Larcher, W. Plant life in cold climates. Symp. Soc. Exp. Biol. 1988, 42, 25–57. [Google Scholar] [PubMed]
- Scherrer, D.; Schmid, S.; Körner, C. Elevational species shifts in a warmer climate are overestimated when based on weather station data. Int. J. Biometeorol. 2011, 55, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Levis, N.A.; Pfennig, D.W. Evaluating ‘plasticity-first’ evolution in nature: Key criteria and empirical approaches. Trends Ecol. Evol. 2016, 31, 563–574. [Google Scholar] [CrossRef]
- Dewitt, T.J.; Sih, A.; Wilson, D.S. Costs and limits of phenotypic plasticity. Trends Ecol. Evol. 1998, 13, 77–81. [Google Scholar] [CrossRef]
- Pál, C.; Miklós, I. Epigenetic inheritance, genetic assimilation and speciation. J. Theor. Biol. 1999, 200, 19–37. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, J.; Dudley, S.A. Testing the adaptive plasticity hypothesis for plant responses to neighbors. Plant Species Biol. 1996, 11, 59–67. [Google Scholar] [CrossRef]
- Dudley, S.A.; Schmitt, J. Testing the adaptive plasticity hypothesis: Density-dependent selection on manipulated stem length in Impatiens capensis. Am. Nat. 1996, 147, 445–465. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Segal, D.L.; Hoyle, G.L.; Schrey, A.W.; Verhoeven, K.J.; Richards, C.L. Adaptive plasticity and epigenetic variation in response to warming in an Alpine plant. Ecol. Evol. 2015, 5, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Solé-Medina, A.; Robledo-Arnuncio, J.J.; Ramírez-Valiente, J.A. Multi-trait genetic variation in resource-use strategies and phenotypic plasticity correlates with local climate across the range of a Mediterranean oak (Quercus faginea). New Phytol. 2022, 234, 462–478. [Google Scholar] [CrossRef] [PubMed]
- Stotz, G.C.; Salgado-Luarte, C.; Escobedo, V.M.; Valladares, F.; Gianoli, E. Global trends in phenotypic plasticity of plants. Ecol. Lett. 2021, 24, 2267–2281. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B.; Lande, R.; Slatkin, M. A neo-Darwinian commentary on macroevolution. Evolution 1982, 36, 474–498. [Google Scholar] [CrossRef] [PubMed]
- Björkman, O. Responses to different quantum flux densities. In Physiological Plant Ecology I: Responses to the Physical Environment; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1981; pp. 57–107. ISBN 978-3-642-68090-8. [Google Scholar]
- Boardman, N.K. Comparative photosynthesis of sun and shade plants. Annu. Rev. Plant Physiol. 1977, 28, 355–377. [Google Scholar] [CrossRef]
- Terashima, I.; Hanba, Y.T.; Tholen, D.; Niinemets, U. Leaf functional anatomy in relation to photosynthesis. Plant Physiol. 2011, 155, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, T.C.; Nishio, J.N.; Smith, W.K. Leaves and light capture: Light propagation and gradients of carbon fixation within leaves. Trends Plant Sci. 1996, 1, 65–70. [Google Scholar] [CrossRef]
- Gratani, L.; Crescente, M.F.; D’Amato, V.; Ricotta, C.; Frattaroli, A.R.; Puglielli, G. Leaf traits variation in Sesleria nitida growing at different altitudes in the Central Apennines. Photosynthetica 2014, 52, 386–396. [Google Scholar] [CrossRef]
- Bréhélin, C.; Kessler, F.; van Wijk, K.J. Plastoglobules: Versatile lipoprotein particles in plastids. Trends Plant Sci. 2007, 12, 260–266. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Chow, W.S.; De Las Rivas, J. Dynamic flexibility in the structure and function of photosystem II in higher plant thylakoid membranes: The grana enigma. Photosynth. Res. 2008, 98, 575–587. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure-function coordination. J. For. Res. 2020, 31, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Press, M.C. The functional significance of leaf structure: A search for generalizations. New Phytol. 1999, 143, 213–219. [Google Scholar] [CrossRef]
- Pellissier, L.; Roger, A.; Bilat, J.; Rasmann, S. High elevation Plantago lanceolata plants are less resistant to herbivory than their low elevation conspecifics: Is it just temperature? Ecography 2014, 37, 950–959. [Google Scholar] [CrossRef] [Green Version]
- Buckley, J.; Widmer, A.; Mescher, M.C.; De Moraes, C.M. Variation in growth and defence traits among plant populations at different elevations: Implications for adaptation to climate change. J. Ecol. 2019, 107, 2478–2492. [Google Scholar] [CrossRef]
- Zhang, N.N.; Tonsor, S.J.; Traw, M.B. A geographic cline in leaf salicylic acid with increasing elevation in Arabidopsis thaliana. Plant Signal. Behav. 2015, 10, e992741. [Google Scholar] [CrossRef] [Green Version]
- Rasmann, S.; Buri, A.; Gallot-Lavallee, M.; Joaquim, J.; Purcell, J.; Pellissier, L. Differential allocation and deployment of direct and indirect defences by Vicia sepium along elevation gradients. J. Ecol. 2014, 102, 930–938. [Google Scholar] [CrossRef]
- Løe, G.; Toräng, P.; Gaudeul, M.; Ågren, J. Trichome production and spatiotemporal variation in herbivory in the perennial herb Arabidopsis lyrata. Oikos 2007, 116, 134–142. [Google Scholar] [CrossRef]
- Pellissier, L.; Fiedler, K.; Ndribe, C.; Dubuis, A.; Pradervand, J.N.; Guisan, A.; Rasmann, S. Shifts in species richness, herbivore specialization, and plant resistance along elevation gradients. Ecol. Evol. 2012, 2, 1818–1825. [Google Scholar] [CrossRef]
- Erelli, M.C.; Ayres, M.P.; Eaton, G.K. Altitudinal patterns in host suitability for forest insects. Oecologia 1998, 117, 133–142. [Google Scholar] [CrossRef]
- Descombes, P.; Marchon, J.; Pradervand, J.N.; Bilat, J.; Guisan, A.; Rasmann, S.; Pellissier, L. Community-level plant palatability increases with elevation as insect herbivore abundance declines. J. Ecol. 2017, 105, 142–151. [Google Scholar] [CrossRef]
- Callis-Duehl, K.; Vittoz, P.; Defossez, E.; Rasmann, S. Community-level relaxation of plant defenses against herbivores at high elevation. Plant Ecol. 2017, 218, 291–304. [Google Scholar] [CrossRef]
- Franks, P.J.; Drake, P.L.; Beerling, D.J. Plasticity in maximum stomatal conductance constrained by negative correlation between stomatal size and density: An analysis using Eucalyptus globulus. Plant Cell Environ. 2009, 32, 1737–1748. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.A. Polyploidy and novelty in flowering plants. Am. Nat. 1983, 122, 1–25. [Google Scholar] [CrossRef]
- Doyle, J.J.; Coate, J.E. Polyploidy, the nucleotype, and novelty: The impact of genome doubling on the biology of the cell. Int. J. Plant Sci. 2019, 180, 1–52. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Nuclear volume control by nucleoskeletal DNA, selection for cell volume and cell growth rate, and the solution of the DNA C-value paradox. J. Cell Sci. 1978, 34, 247–278. [Google Scholar] [CrossRef]
- Byrne, M.C.; Nelson, C.J.; Randall, D.D. Ploidy effects on anatomy and gas exchange of tall Fescue Leaves. Plant Physiol. 1981, 68, 891–893. [Google Scholar] [CrossRef] [Green Version]
- Bomblies, K. When everything changes at once: Finding a new normal after genome duplication. Proc. R. Soc. B Biol. Sci. 2020, 287, 20202154. [Google Scholar] [CrossRef]
- Hodgson, J.G.; Sharafi, M.; Jalili, A.; Diaz, S.; Montserrat-Marti, G.; Palmer, C.; Cerabolini, B.; Pierce, S.; Hamzehee, B.; Asri, Y.; et al. Stomatal vs. genome size in angiosperms: The somatic tail wagging the genomic dog? Ann. Bot. 2010, 105, 573–584. [Google Scholar] [CrossRef]
- Ennos, R.A.; French, G.C.; Hollingsworth, P.M. Conserving taxonomic complexity. Trends Ecol. Evol. 2005, 20, 164–168. [Google Scholar] [CrossRef]
- Kaplenig, D.; Bertel, C.; Arc, E.; Villscheider, R.; Ralser, M.; Kolář, F.; Wos, G.; Hülber, K.; Kranner, I.; Neuner, G. Repeated colonization of alpine habitats by Arabidopsis arenosa viewed through freezing resistance and ice management strategies. Plant Biol. 2022, 24, 939–949. [Google Scholar] [CrossRef]
- Wos, G.; Bohutínská, M.; Nosková, J.; Mandáková, T.; Kolář, F. Parallelism in gene expression between foothill and alpine ecotypes in Arabidopsis arenosa. Plant J. 2021, 105, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Kubínová, L. Recent stereological methods for measuring leaf anatomical characteristics—Estimation of the number and sizes of stomata and mesophyll-cells. J. Exp. Bot. 1994, 45, 119–127. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Holzinger, A.; Roleda, M.Y.; Lütz, C. The vegetative arctic freshwater green alga Zygnema is insensitive to experimental UV exposure. Micron 2009, 40, 831–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Dray, S.; Dufour, A.B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertel, C.; Kaplenig, D.; Ralser, M.; Arc, E.; Kolář, F.; Wos, G.; Hülber, K.; Holzinger, A.; Kranner, I.; Neuner, G. Parallel Differentiation and Plastic Adjustment of Leaf Anatomy in Alpine Arabidopsis arenosa Ecotypes. Plants 2022, 11, 2626. https://doi.org/10.3390/plants11192626
Bertel C, Kaplenig D, Ralser M, Arc E, Kolář F, Wos G, Hülber K, Holzinger A, Kranner I, Neuner G. Parallel Differentiation and Plastic Adjustment of Leaf Anatomy in Alpine Arabidopsis arenosa Ecotypes. Plants. 2022; 11(19):2626. https://doi.org/10.3390/plants11192626
Chicago/Turabian StyleBertel, Clara, Dominik Kaplenig, Maria Ralser, Erwann Arc, Filip Kolář, Guillaume Wos, Karl Hülber, Andreas Holzinger, Ilse Kranner, and Gilbert Neuner. 2022. "Parallel Differentiation and Plastic Adjustment of Leaf Anatomy in Alpine Arabidopsis arenosa Ecotypes" Plants 11, no. 19: 2626. https://doi.org/10.3390/plants11192626