
Palyno-Morphological Characteristics as a Systematic Approach in the Identification of Halophytic Poaceae Species from a Saline Environment

Abstract
:1. Introduction
2. Results
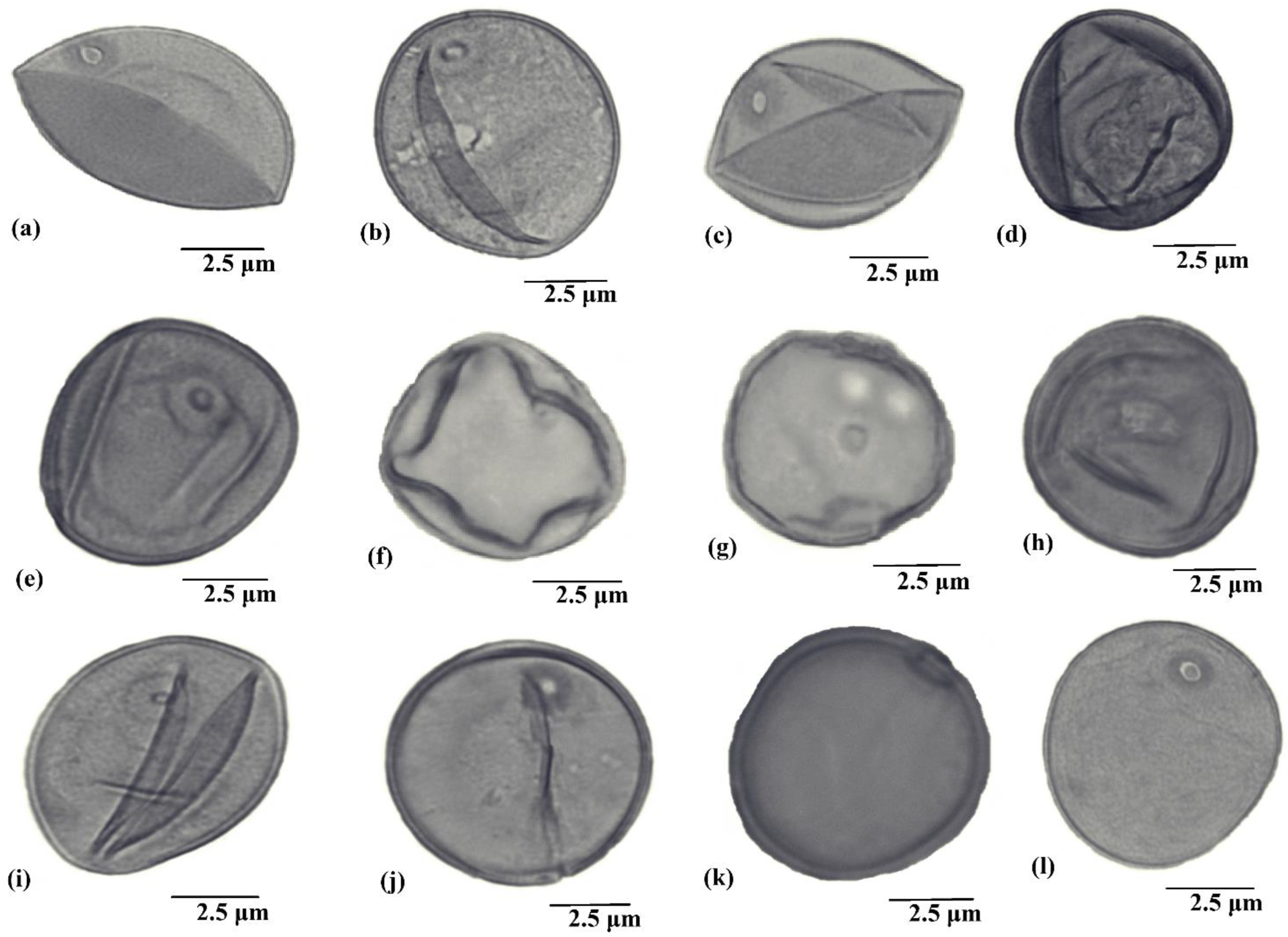
2.1. Diversity in Pollen Size and Shape
2.2. Pollen Aperture
2.3. Exine Ornamentation
2.4. Pollen Viability
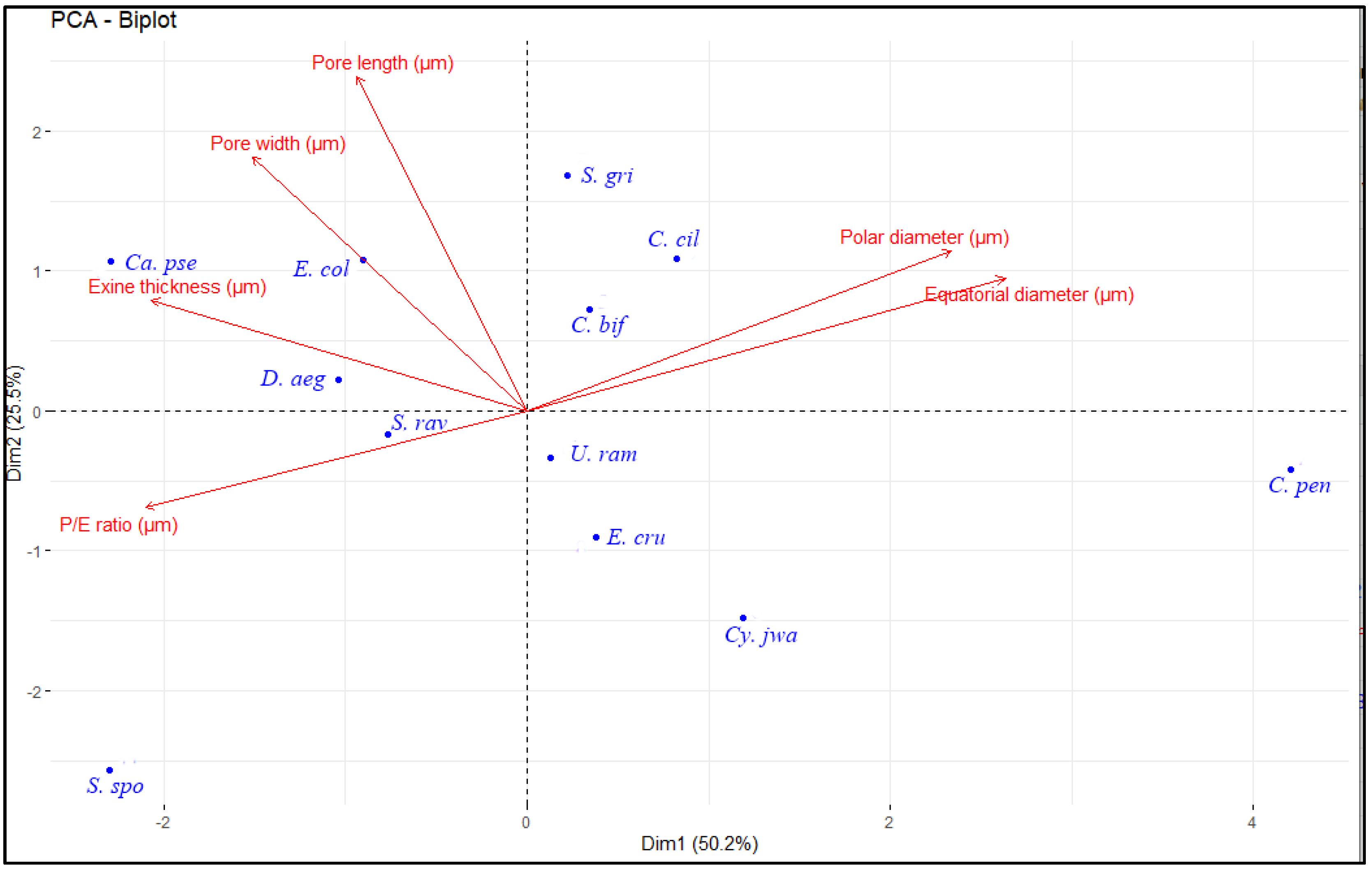
2.5. Principal Component Analysis
3. Discussion
4. Materials and Methods
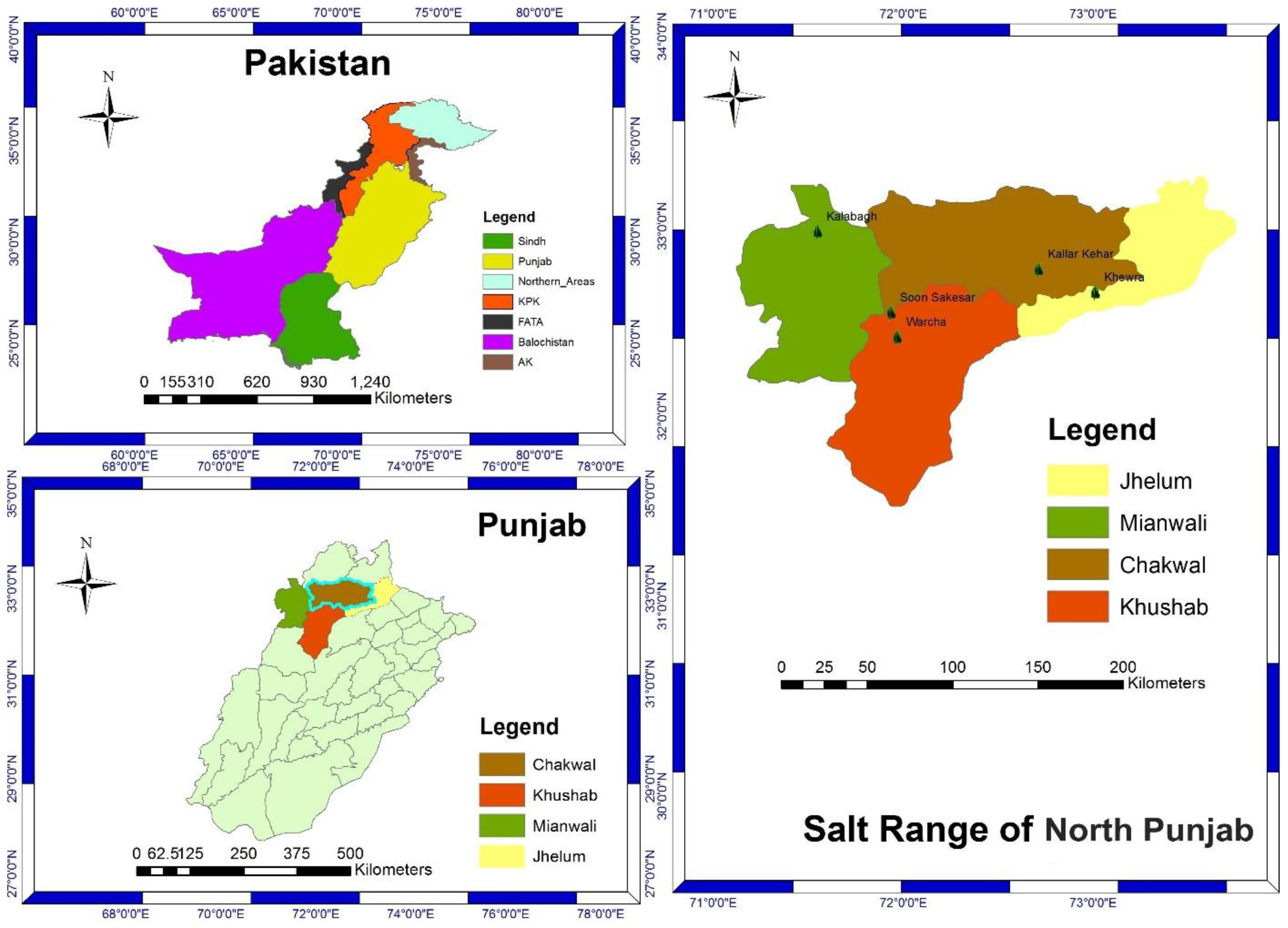
4.1. Collection and Identification
4.2. Light Microscopic Analysis of Pollen in Salt-Tolerant Grasses
4.3. Scanning Electron Microscopic Analysis of Pollen in Salt-Tolerant Grasses
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schüler, L.; Behling, H. Poaceae pollen grain size as a tool to distinguish past grasslands in South America: A new methodological approach. Veg. Hist. Archaeobotany 2011, 20, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Perveen, A.; Qaiser, M. Pollen flora of Pakistan–LXIX. Poacae. Pak. J. Bot. 2012, 44, 747–756. [Google Scholar]
- Edwards, E.J.; Smith, S.A. Phylogenetic analyses reveal the shady history of C4 grasses. Proc. Natl. Acad. Sci. USA 2010, 107, 2532–2537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, F.; Khan, M.A.; Ahmad, M.; Zafar, M.; Nazir, A.; Marwat, S.K. Taxonomic studies of grasses and their indigenous uses in the salt range area of Pakistan. Afr. J. Biotechnol. 2009, 8, 231–249. [Google Scholar]
- Khan, R.; Ullah, A.; Khan, B.; Khan, S.M.; Rashid, A. Comparative foliar micro morphological studies of tribe Arundinacea, Aristideae and Chlorideae from Malakand Agency, Khyber Pakhtunkhwa, Pakistan. Pak. J. Bot. 2017, 49, 33–42. [Google Scholar]
- Knox, R.; Taylor, P.; Smith, P.; Hough, T.; Ong, E.; Suphioglu, C.; Lavithis, M.; Davies, S.; Avjioglu, A.; Singh, M. Pollen allergens: Botanical aspects. In Molecular Biology and Immunology of Allergens; CRC Press: Boca Raton, FL, USA, 1993; pp. 31–34. [Google Scholar]
- Singh, A.; Parabia, M. The floral diversity of Gujarat State: A review. Indian For. 2003, 129, 1461–1469. [Google Scholar]
- Gould, F.W. Grass Systematics; Mc Graw-Hill Book Company: New York, NY, USA, 1968. [Google Scholar]
- Sarir, M.; Marwat, K.; Khattak, J. Studies on some halophytes of Peshawar District. Pak. J. Bot. 1984, 16, 49–51. [Google Scholar]
- Kearney, T.H.; Briggs, L.; Shantz, H.; McLane, J.; Piemeisel, R. Indicator significance of vegetation in Tooele Valley, Utah. J. Agric. Res. 1914, 1, 365–417. [Google Scholar]
- Hilgard, E.W. Soils: Their Formation, Properties, Composition, and Relations to Climate and Plant Growth in the Humid and Arid Regions; Macmillan: New York, NY, USA, 1921. [Google Scholar]
- Naz, N.; Hameed, M.; Nawaz, T.; Batool, R.; Ashraf, M.; Ahmad, F.; Ruby, T. Structural adaptations in the desert halophyte Aeluropus lagopoides (Linn.) Trin. ex Thw. under high salinity. J. Biol. Res. 2013, 19, 150–164. [Google Scholar]
- Farooq, A.; Hameed, M.; Ahmad, K.S.; Ashraf, M. Significance of Anatomical Markers in Tribe Paniceae (Poaceae) from the Salt Range, Pakistan. Int. J. Agric. Biol. 2015, 17, 271–279. [Google Scholar]
- Monteverdi, C.M.; Lauteri, M.; Valentini, R. Biodiversity of plant species and adaptation to drought and salt conditions. Selection of species for sustainable reforestation activity to combat desertification. In Biosaline Agriculture and High Salinity Tolerance; Springer: Berlin/Heidelberg, Germany, 2008; pp. 197–206. [Google Scholar]
- Khan, M.A.; Qaiser, M. Halophytes of Pakistan: Characteristics, distribution and potential economic usages. In Sabkha Ecosystems; Springer: Berlin/Heidelberg, Germany, 2006; pp. 129–153. [Google Scholar]
- Khan, R.; Abidin, S.Z.U.; Ahmad, M.; Zafar, M.; Liu, J.; Amina, H. Palyno-morphological characteristics of gymnosperm flora of Pakistan and its taxonomic implications with LM and SEM methods. Microsc. Res. Tech. 2018, 81, 74–87. [Google Scholar] [CrossRef]
- Belhadj, S.; Derridj, A.; Aigouy, T.; Gers, C.; Gauquelin, T.; Mevy, J.P. Comparative morphology of leaf epidermis in eight populations of Atlas pistachio (Pistacia atlantica Desf., Anacardiaceae). Microsc. Res. Tech. 2007, 70, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhao, N.-X.; Hao, G. Pollen morphology of the Chloridoideae (Gramineae). Grana 2004, 43, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Dórea, M.D.C. Morfologia Polínica, Fenologia Reprodutiva E Biologia Floral de Espécies Florestais de Poaceae; Universidade Estadual de Feira de Santana: Feira de Santana, Brazil, 2011. [Google Scholar]
- Skvarla, J.J.; Rowley, J.R.; Hollowell, V.C.; Chissoe, W.F. Annulus–pore relationship in Gramineae (Poaceae) pollen: The pore margin of Pariana. Am. J. Bot. 2003, 90, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Mander, L.; Punyasena, S.W. Grass pollen surface ornamentation: A review of morphotypes and taxonomic utility. J. Micropalaeontol. 2016, 35, 121–124. [Google Scholar] [CrossRef]
- Nazir, A.; Khan, M.; Abbasi, A.; Zahidullah. Palynological studies in Tribe Aveneae (Poaceae) from Potohar of Pakistan. Int. J. Sci. 2013, 10, 120–125. [Google Scholar]
- Jan, F. Palynological Studies and Holocene Ecosystem Dynamics in North Western Khyber Pakhtunkhwa Province of Pakistan in the Hindu Kush Himalayan Region. Ph.D. Thesis, Georg-August Universität, Göttingen, Germany, 2015. [Google Scholar]
- Joly, C.; Barillé, L.; Barreau, M.; Mancheron, A.; Visset, L. Grain and annulus diameter as criteria for distinguishing pollen grains of cereals from wild grasses. Rev. Palaeobot. Palynol. 2007, 146, 221–233. [Google Scholar] [CrossRef]
- Katsiotis, A.; Forsberg, R. Pollen grain size in four ploidy levels of genusAvena. Euphytica 1995, 83, 103–108. [Google Scholar] [CrossRef]
- Faegri, K.; Kaland, P.E.; Krzywinski, K. Textbook of Pollen Analysis; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 1989. [Google Scholar]
- Andersen, T.S.; Bertelsen, F. Scanning electron microscope studies of pollen of cereals and other grasses. Grana 1972, 12, 79–86. [Google Scholar] [CrossRef]
- Arora, A.; Modi, A. An acetolysis technique for pollen slide preparation. Indian J. Aerobiol. 2008, 21, 90–91. [Google Scholar]
- Quamar, M.; Ali, S.N.; Pandita, S.K.; Singh, Y. Modern pollen rain from Udhampur (Jammu and Kashmir), India: Insights into pollen production, dispersal, transport and preservation. Palynology 2018, 42, 55–65. [Google Scholar] [CrossRef]
- Aftab, R.; Perveen, A. A palynological study of some cultivated trees from Karachi. Pak. J. Bot. 2006, 38, 15–28. [Google Scholar]
- Ahmad, F.; Khan, M.A.; Ahmad, M.; Zafar, M.; Khan, A.; Iqbal, Z. Palynological studies in tribe Chlorideae (Poaceae) from salt range of Pakistan. Afr. J. Biotechnol. 2011, 10, 8909–8913. [Google Scholar]
- Guohua, M.; Xuelin, H.; Qiusheng, X.; Bunn, E. Multiporate pollen and apomixis in Panicoideae. Pak. J. Bot. 2009, 41, 2073–2082. [Google Scholar]
- Perveen, A. A contribution to the pollen morphology of family Gramineae. World Appl. Sci. J. 2006, 1, 60–65. [Google Scholar]
- Correa, A.S.; Guimaraes, M.M.; Cruz-Barros, M.D.; Begale, F. Flora polínica da reserva do parque estadual das fontes do ipiranga (São Paulo, Brasil). Familia 2005, 176, 269–282. [Google Scholar]
- Aboulaich, N.; Bouziane, H.; Kadiri, M.; del Mar Trigo, M.; Riadi, H.; Kazzaz, M.; Merzouki, A. Pollen production in anemophilous species of the Poaceae family in Tetouan (NW Morocco). Aerobiologia 2009, 25, 27–38. [Google Scholar] [CrossRef]
- Bush, M.B. On the interpretation of fossil Poaceae pollen in the lowland humid neotropics. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 177, 5–17. [Google Scholar] [CrossRef]
- Beug, H. Leitfaden der Pollenbestimmung, Lieferung 1; Fischer: Stuttgart, Germany, 1961. [Google Scholar]
- Salgado-Labouriau, M.; Rinaldi, M. Measurements of Gramineae pollen of the Venezuelan mountains. Rev. Bras. Biol. 1989, 50, 115–122. [Google Scholar]
- Strömberg, C.A. Evolution of grasses and grassland ecosystems. Annu. Rev. Earth Planet. Sci. 2011, 39, 517–544. [Google Scholar] [CrossRef]
- Piperno, D.R. Phytoliths: A Comprehensive Guide for Archaeologists and Paleoecologists; Rowman Altamira: Lanham, MD, USA, 2006. [Google Scholar]
- Ahmad, F.; Khan, M.A.; Ahmad, M.; Hameed, M.; Tareen, R.B.; Zafar, M.; Jabeen, A. Taxonomic application of foliar anatomy in grasses of Tribe Eragrostideae (Poaceae) from Salt Range of Pakistan. Pak. J. Bot. 2011, 43, 2277–2284. [Google Scholar]
- Chaturvedi, M.; Datta, K. Pollen morphology in saccharum l.(poaceae)—Wild and cultivated sugar cane species. Feddes Repert. 2001, 112, 387–390. [Google Scholar] [CrossRef]
- Radaeski, J.N.; Evaldt, A.C.P.; Bauermann, S.G.; de Lima, G.L. Diversidade de grãos de pólen e esporos dos Campos do sul do Brasil: Descrições morfológicas e implicações paleoecológicas. Iheringia Série Botânica 2014, 69, 107–132. [Google Scholar]
- Radaeski, J.N.; Bauermann, S.G.; Evaldt, A.C.P.; de Lima, G.L. Grãos de pólen das formações campestres sul-brasileiras. Rev. Iniciação Científica ULBRA 2011, 1, 59–67. [Google Scholar]
- Morgado, L.N.; Gonçalves-Esteves, V.; Resendes, R.; Ventura, M.A.M. Pollen morphology of Poaceae (Poales) in the Azores, Portugal. Grana 2015, 54, 282–293. [Google Scholar] [CrossRef]
- Radaeski, J.N.; Bauermann, S.G.; Pereira, A.B. Poaceae pollen from Southern Brazil: Distinguishing Grasslands (Campos) from Forests by Analyzing a Diverse Range of Poaceae Species. Front. Plant Sci. 2016, 7, 1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dwari, S.; Mondal, A.K. Systematic studies (morphology, anatomy, palynology) of an economically important grass Sporobolus diandrus (Retz.) P. Beauv.(Poaceae). Ann. Plant Sci. 2018, 7, 2226–2230. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, H.; Waseem, M. Conservation status of some medicinal plants of the Salt Range. Zonas Áridas 2006, 8, 40–47. [Google Scholar]
- Erdtman, G. The acetolysis method-a revised description. Sven. Bot. Tidskr. 1960, 54, 516–564. [Google Scholar]
- Ronald, O. Pollen and spores. Am. Assoc. Strat. Palynol. Ed. 2000, 2, 13–21. [Google Scholar]
- Barth, O.M.; Melhem, T.S. Glossário Ilustrado de Palinologia; Editora da Unicamp, Unicamp: Campinas, Brazil, 1988. [Google Scholar]
- Punt, W.; Hoen, P.; Blackmore, S.; Nilsson, S.; Le Thomas, A. Glossary of pollen and spore terminology. Rev. Palaeobot. Palynol. 2007, 143, 1–81. [Google Scholar] [CrossRef]
- Hesse, M.; Halbritter, H.; Zetter, R.; Weber, M.; Buchner, R.; Frosch-Radivo, R.; Ulrich, S. Pollen Terminology: An Illustrated Handbook; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Ullah, F.; Zafar, M.; Ahmad, M.; Dilbar, S.; Shah, S.N.; Sohail, A.; Zaman, W.; Iqbal, M.; Bahadur, S.; Tariq, A. Pollen morphology of subfamily Caryophylloideae (Caryophyllaceae) and its taxonomic significance. Microsc. Res. Tech. 2018, 81, 704–715. [Google Scholar] [PubMed]
- Dafni, A.; Firmage, D. Pollen viability and longevity: Practical, ecological and evolutionary implications. In Pollen and Pollination; Springer: Berlin/Heidelberg, Germany, 2000; Volume 222, pp. 113–132. [Google Scholar]
- Lozano-Vega, G.; Benezeth, Y.; Marzani, F.; Boochs, F. Classification of pollen apertures using bag of words. In Proceedings of the International Conference on Image Analysis and Processing, Naples, Italy, 9–13 September 2013; pp. 712–721. [Google Scholar]
- Mazari, P.; Khansup, M.A.; Alisup, B.; Buxsup, H.; Khansup, K.Y.; Mughalsup, S.; Ahmadsup, M.; Zafarsup, M.; Akhtersup, A. Palynological diversity in selected medicinal plant species of Asteraceae (Compositae) from flora of Kaghan Valley. J. Med. Plants Res. 2012, 6, 2747–2753. [Google Scholar]
- Sufyan, M.; Badshah, I.; Ahmad, M.; Zafar, M.; Bahadur, S.; Rashid, N. Identification of Medicinally Used Flora Using Pollen Features Imaged in the Scanning Electron Microscopy in the Lower Margalla Hills Islamabad Pakistan. Microsc. Microanal. 2018, 24, 292–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mbagwu, F.; Chime, E.; Unamba, C. Palynological studies on five species of Asteraceae. Afr. J. Biotechnol. 2009, 8, 1222–1225. [Google Scholar]
- Hanif, U.; Mazhar, M.; Sardar, A.A. Palynomorphological studies of some Ornamental plants of mall road, Lahore. Biologia 2013, 59, 147–156. [Google Scholar]
- Corrêa, A.M.D.S.; Fonseca, K. Pollen Flora of Reserva do Parque Estadual das Fontes do Ipiranga (São Paulo, São Paulo State, Brazil). Hoehnea 2015, 42, 597–602. [Google Scholar] [CrossRef]
- Mazari, P.; Liu, Q.; Khan, M.A.; Sadia, S.; Ahmad, L. Pollen Morphology and Pollen Fertility Estimation of Three Medicinal Plant Species of Hypericum L. from Kaghan Valley, Northern Pakistan. Am. J. Plant Sci. 2017, 8, 3073–3083. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, T.; Qaiser, M. A palynological study of the family Gramineae from Karachi-Pakistan. Pak. J. Bot. 1988, 20, 161–176. [Google Scholar]
- Mander, L.; Li, M.; Mio, W.; Fowlkes, C.C.; Punyasena, S.W. Classification of grass pollen through the quantitative analysis of surface ornamentation and texture. Proc. R. Soc. Lond. B Biol. Sci. 2013, 280, 20131905. [Google Scholar] [CrossRef]
- Linder, H.; Ferguson, I. Notes on the pollen morphology and phylogeny Restionales and Poales. Grana 1985, 24, 65–76. [Google Scholar] [CrossRef]
- Needham, I.; Vorontsova, M.S.; Banks, H.; Rudall, P.J. Pollen of Malagasy grasses as a potential tool for interpreting grassland palaeohistory. Grana 2015, 54, 247–262. [Google Scholar] [CrossRef]
- Neha, K.; Kalkar, S. Pollen morphology of millets-exine surface ultrastructure. Asian J. Exp. Biol. Sci. 2010, 85–90. [Google Scholar]
- Ogebebor. Pollen Morphology and Plant Taxonomy; Edo State University Ekpoma: Edo State, Nigeria, 1996. [Google Scholar]
- Jan, F.; Schüler, L.; Behling, H. Trends of pollen grain size variation in C3 and C4 Poaceae species using pollen morphology for future assessment of grassland ecosystem dynamics. Grana 2014, 54, 129–145. [Google Scholar] [CrossRef]
- Khola, G.; Hanif, U. Palynological study of soil sample collected from an archaeological site (Gulabi Bagh) in Lahore, Pakistan. J. Anim. Plant Sci 2012, 22, 1113–1117. [Google Scholar]
- Woodehouse, R.P. Pollen Grains: Their Structure, Identification and Significance in Science and Medicine; Mc Graw-Hill Publishing Co., Ltd.: London, UK, 1935; 574p. [Google Scholar]
- Perveen, A.; Qaiser, M. Pollen flora of Pakistan-LI-Caryophyllaceae. Pak. J. Bot. 2006, 38, 901. [Google Scholar]
- Hameed, M.; Naz, N.; Ahmad, M.S.A.; Islam-ud-Din, R.A. Morphological adaptations of some grasses from the salt range, Pakistan. Pak. J. Bot. 2008, 40, 1571–1578. [Google Scholar]
- Mbagwu, F.; Okafor, V.; Okolo, A. Comparative palynological characters of four species of family Poaceae. Int. Sci. Res. J. 2008, 1, 127–129. [Google Scholar]
- Shaheen, S.; Ramzan, S.; Haroon, N.; Hussain, K. Ethnopharmacological And Systematic Studies Of Selected Medicinal Plants Of Pakistan. Pak. J. Sci. 2014, 66, 175–180. [Google Scholar]
- Zafar, M.; Khan, M.A.; Ahmad, M.; Sultana, S. Palynological and taxonomic studies of some weeds from flora of Rawalpindi. Pak. J. Weed Sci. Res. 2006, 12, 99–109. [Google Scholar]
- Tellari, M. Comparative palynological analysis of species of MedicagoL.,Melilotus Mill and Trifolium L.(Leguminoseae) in the phytographic pampas region n (Argentina). Ind. Apic. 1991, 1, 4–13. [Google Scholar]
- Awan, A.; Qureshi, S.; Bano, S.; Khan, M. Study of Pollen fertility of the genus Crepis and Tragopogon from Pakistan. Pak. J. Biol. Sci. 2001, 4, 487–488. [Google Scholar]
- Ashfaq, S.; Ahmad, M.; Zafar, M.; Sultana, S.; Bahadur, S.; Ullah, F.; Zaman, W.; Ahmed, S.N.; Nazish, M. Foliar micromorphology of Convolvulaceous species with special emphasis on trichome diversity from the arid zone of Pakistan. Flora 2019, 255, 110–124. [Google Scholar] [CrossRef]
- Kenfack, D. Resurrection in Carapa (Meliaceae): A reassessment of morphological variation and species boundaries using multivariate methods in a phylogenetic context. Bot. J. Linn. Soc. 2011, 165, 186–221. [Google Scholar] [CrossRef] [Green Version]
- Lopes, R.C.; Andreata, R.H.P.; Cartaxo-Pinto, S.; Trovó, M.; Gonçalves-Esteves, V. Pollen morphology and wall structure of Neotropical species of Herreria and Clara (Asparagaceae-Agavoideae) and its taxonomic implications. Plant Syst. Evol. 2013, 299, 25–34. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Name | Pollen Size | Pollen Shape | Annulus | Operculum | Aperturate | Polar Diameter (µm) Min (Mean ± Std Error) Max | Equatorial Diameter (µm) Min (Mean ± Std Error) Max | P/E Ratio (µm) | Exine Thickness (µm) Min (Mean ± Std Error) Max | Pore Length (µm) Min (Mean ± Std Error) Max | Pore Width (µm) Min (Mean ± Std Error) Max | Sculpturing (SEM) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Calamagrostis pseudophragmites (Haller f.) Koeler | Medium | Subspheroidal | + | − | Monoporate | 27.25 (27.85 ± 0.48) 29.75 | 25.50 (26.80 ± 0.54) 28.0 | 1.03 | 1.0 (1.1 ± 0.03) 1.2 | 1.7 (1.94 ± 0.09) 2.2 | 1.8 (1.96 ± 0.05) 2.1 | Microechinate-areolate |
| Cenchrus biflorus Roxb. | Medium | Subspheroidal | + | − | Monoporate | 30.50 (35.5 ± 1.66) 40.25 | 28.0 (33.5 ± 1.60) 37.75 | 1.05 | 0.6 (0.72 ± 0.05) 0.9 | 1.6 (1.86 ± 0.08) 2.1 | 1.7 (1.96 ± 0.08) 2.2 | Microechinate-areolate |
| Cenchrus ciliaris L. | Medium | Subspheroidal | + | − | Monoporate | 27.25 (32.75 ± 2.19) 38.0 | 30.25 (34.05 ± 1.83) 40.25 | 0.96 | 0.5 (0.72 ± 0.10) 0.9 | 1.9 (1.99 ± 0.08) 2.3 | 1.8 (1.82 ± 0.06) 2.1 | Microechinate-areolate |
| Cenchrus pennisetiformis Hochst. and Steud. | Medium | Subspheroidal | + | − | Diporate, Monoporate | 32.50 (38.55 ± 1.58) 41.75 | 43.0 (44.55 ± 0.91) 47.75 | 0.86 | 0.6 (0.72 ± 0.05) 1.1 | 1.6 (1.76 ± 0.05) 1.9 | 1.5 (1.66 ± 0.08) 1.9 | Microechinate-areolate |
| Cymbopogon jwarancusa (Jones) Schult. | Medium | Subspheroidal | + | + | Monoporate | 25.50 (31.20 ± 2.19) 38.25 | 29.75 (31.50 ± 0.77) 34.0 | 0.99 | 0.5 (0.64 ± 0.07) 0.9 | 1.6 (1.80 ± 0.07) 2.0 | 1.5 (1.72 ± 0.06) 1.9 | Microechinate |
| Dactyloctenium aegyptium (L.) Willd. | Medium | Subspheroidal | + | + | Monoporate | 23.0 (27.85 ± 1.48) 32.25 | 26.25 (28.65 ± 0.69) 30.25 | 0.97 | 0.9 (1.06 ± 0.05) 1.2 | 1.8 (1.92 ± 0.03) 2.0 | 1.7 (1.80 ± 0.03) 1.9 | Microechinate-areolate |
| Echinochloa colona (L.) Link | Medium | Subspheroidal | + | + | Monoporate | 25.50 (29.65 ± 1.84) 35.75 | 28.0 (30.35 ± 0.88) 33.25 | 0.97 | 0.9 (1.04 ± 0.06) 1.2 | 1.5 (1.96 ± 0.10) 2.1 | 1.6 (1.86 ± 0.07) 2.0 | Microechinate-areolate |
| Echinochloa crus-galli (L.) P. Beauv. | Medium | Subspheroidal | + | − | Monoporate | 22.75 (27.35 ± 1.65) 32.75 | 24.50 (30.05 ± 1.77) 33.50 | 0.91 | 0.6 (0.78 ± 0.09) 1.1 | 1.8 (1.78 ± 0.08) 2.2 | 1.7 (1.84 ± 0.03) 1.9 | Microechinate-areolate |
| Saccharum griffithii Munro ex Boiss. | Medium | Subspheroidal | + | − | Monoporate | 26.25 (31.75 ± 1.97) 35.50 | 31.25 (33.85 ± 1.01) 37.25 | 0.93 | 0.5 (0.84 ± 0.12) 1.1 | 1.6 (1.96 ± 0.11) 2.3 | 1.8 (1.94 ± 0.05) 2.1 | Microechinate-areolate |
| Saccharum ravennae (L.) L. | Medium | Subspheroidal | + | − | Monoporate | 24.75 (29.80 ± 2.48) 38.5 | 23.0 (28.70 ± 2.57) 38.0 | 1.03 | 0.6 (0.92 ± 0.10) 1.2 | 1.7 (1.90 ± 0.08) 2.1 | 1.6 (1.80 ± 0.07) 2.0 | Microechinate-areolate |
| Saccharum spontaneum L. | Medium | Subspheroidal | + | − | Monoporate | 20.25 (23.10 ± 1.04) 25.5 | 15.25 (20.95 ± 1.94) 25.25 | 1.10 | 0.7 (0.9 ± 0.08) 1.1 | 1.6 (1.76 ± 0.05) 1.9 | 1.7 (1.78 ± 0.03) 1.9 | Microechinate-areolate |
| Urochloa ramosa (L.) T.Q. Nguyen | Medium | Subspheroidal | + | − | Monoporate | 22.25 (27.85 ± 2.88) 32.25 | 25.50 (29.60 ± 1.65) 33.75 | 0.94 | 0.6 (0.78 ± 0.05) 0.9 | 1.7 (1.90 ± 0.07) 2.1 | 1.6 (1.78 ± 0.07) 2.0 | Microechinate-areolate |
| Botanical Name | No. of Viable Pollen | No. of Non-Viable Pollen | Viability (%) | Non-Viability (%) |
|---|---|---|---|---|
| Calamagrostis pseudophragmites (Haller f.) Koeler | 95 | 16 | 85.58 | 14.41 |
| Cenchrus biflorus Roxb. | 104 | 23 | 81.88 | 18.11 |
| Cenchrus ciliaris L. | 93 | 12 | 88.57 | 11.42 |
| Cenchrus pennisetiformis Hochst. and Steud. | 113 | 28 | 80.14 | 19.85 |
| Cymbopogon jwarancusa (Jones) Schult. | 67 | 8 | 89.33 | 10.66 |
| Dactyloctenium aegyptium (L.) Willd. | 69 | 7 | 90.78 | 9.21 |
| Echinochloa colona (L.) Link | 113 | 25 | 81.88 | 18.11 |
| Echinochloa crus-galli (L.) P. Beauv. | 56 | 15 | 78.87 | 21.12 |
| Saccharum griffithii Munro ex Boiss. | 102 | 17 | 85.71 | 14.28 |
| Saccharum ravennae (L.) L. | 62 | 11 | 84.93 | 15.06 |
| Saccharum spontaneum L. | 98 | 9 | 91.58 | 8.41 |
| Urochloa ramosa (L.) T.Q. Nguyen | 87 | 19 | 82.07 | 17.92 |
| Variables | PC1 | PC2 | PC3 | PC4 | PC5 | PC6 |
|---|---|---|---|---|---|---|
| Polar diameter (µm) | 0.4744565 | 0.3249055 | −0.32621870 | 0.43538549 | −0.17548822 | −0.58527955 |
| Equatorial diameter (µm) | 0.5349302 | 0.2709428 | 0.01092927 | 0.25624982 | 0.03831308 | 0.75709210 |
| P/E ratio (µm) | −0.4265074 | −0.1965046 | −0.60566071 | 0.35522178 | −0.45418376 | 0.28317327 |
| Exine thickness (µm) | −0.4201325 | 0.2252430 | 0.53173029 | 0.69077481 | 0.10909227 | −0.03076033 |
| Pore length (µm) | −0.1900636 | 0.6811253 | 0.15652833 | −0.36410765 | −0.58410765 | 0.04107229 |
| Pore width (µm) | −0.3069148 | 0.5174421 | −0.46840025 | −0.09406787 | 0.63903852 | 0.03793646 |
| Eigenvalue | 3.011799883 | 1.532333989 | 0.758921938 | 0.360648574 | 0.333815144 | 0.002480472 |
| Variation (%) | 50.1966647 | 25.5388998 | 12.6486990 | 6.0108096 | 5.5635857 | 0.0413412 |
| Cumulative variance (%) | 50.19666 | 75.73556 | 88.38426 | 94.39507 | 99.95866 | 100.00000 |
| Taxon | Voucher Specimen Number | Collectors | Altitudes (m) | Flowering Period | Locality | District |
|---|---|---|---|---|---|---|
| Calamagrostispseudophragmites (Haller f.) Koeler | ISL.129931 | Moona Nazish and Farhat Ullah | 181.00 | May–October | Kalabagh | Mianwali |
| Cenchrus biflorus Roxb. | ISL.129955 | Moona Nazish | 181.03 | July–September | Khewra | Jhelum |
| Cenchrus ciliaris L. | ISL.129944 | Moona Nazish | 152.90 | March–October | Khewra | Jhelum |
| Cenchrus pennisetiformis Hochst. and Steud. | ISL.129959 | Moona Nazish and Farhat Ullah | 153.00 | April–August | Sakesar | Khushab |
| Cymbopogon jwarancusa (Jones) Schult. | ISL.129971 | Moona Nazish | 152.90 | June–September | Soon Valley | Khushab |
| Dactyloctenium aegyptium (L.) Willd. | ISL.129953 | Moona Nazish | 182.08 | April–October | Kalabagh | Mianwali |
| Echinochloa colona (L.) Link | ISL.129980 | Moona Nazish | 169.00 | March–November | Warcha | Khushab |
| Echinochloa crus-galli (L.) P. Beauv. | ISL.129970 | Moona Nazish | 151.70 | June–September | Soon Valley | Khushab |
| Saccharum griffithii Munro ex Boiss. | ISL.129961 | Moona Nazish | 643.00 | March–August | Kallar Kahar | Chakwal |
| Saccharum ravennae (L.) L. | ISL.129937 | Moona Nazish and Farhat Ullah | 181.10 | April–September | Khewra | Jhelum |
| Saccharum spontaneum L. | ISL.129981 | Moona Nazish | 153.01 | March–September | Sakesar | Khushab |
| Urochloa ramosa (L.) T.Q. Nguyen | ISL.129973 | Moona Nazish | 169.01 | July–October | Warcha | Khushab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazish, M.; Althobaiti, A.T. Palyno-Morphological Characteristics as a Systematic Approach in the Identification of Halophytic Poaceae Species from a Saline Environment. Plants 2022, 11, 2618. https://doi.org/10.3390/plants11192618
Nazish M, Althobaiti AT. Palyno-Morphological Characteristics as a Systematic Approach in the Identification of Halophytic Poaceae Species from a Saline Environment. Plants. 2022; 11(19):2618. https://doi.org/10.3390/plants11192618
Chicago/Turabian StyleNazish, Moona, and Ashwaq T. Althobaiti. 2022. "Palyno-Morphological Characteristics as a Systematic Approach in the Identification of Halophytic Poaceae Species from a Saline Environment" Plants 11, no. 19: 2618. https://doi.org/10.3390/plants11192618