
Novel Antiviral Efficacy of Hedyotis diffusa and Artemisia capillaris Extracts against Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Infection and Immunoregulatory Cytokine Signatures


Abstract
:1. Introduction
2. Materials and Methods
2.1. Herbs
2.2. Cell Lines
2.3. Viruses
2.4. Cell Cytotoxicity Assay and CC50 Determination
2.5. Virus Infection
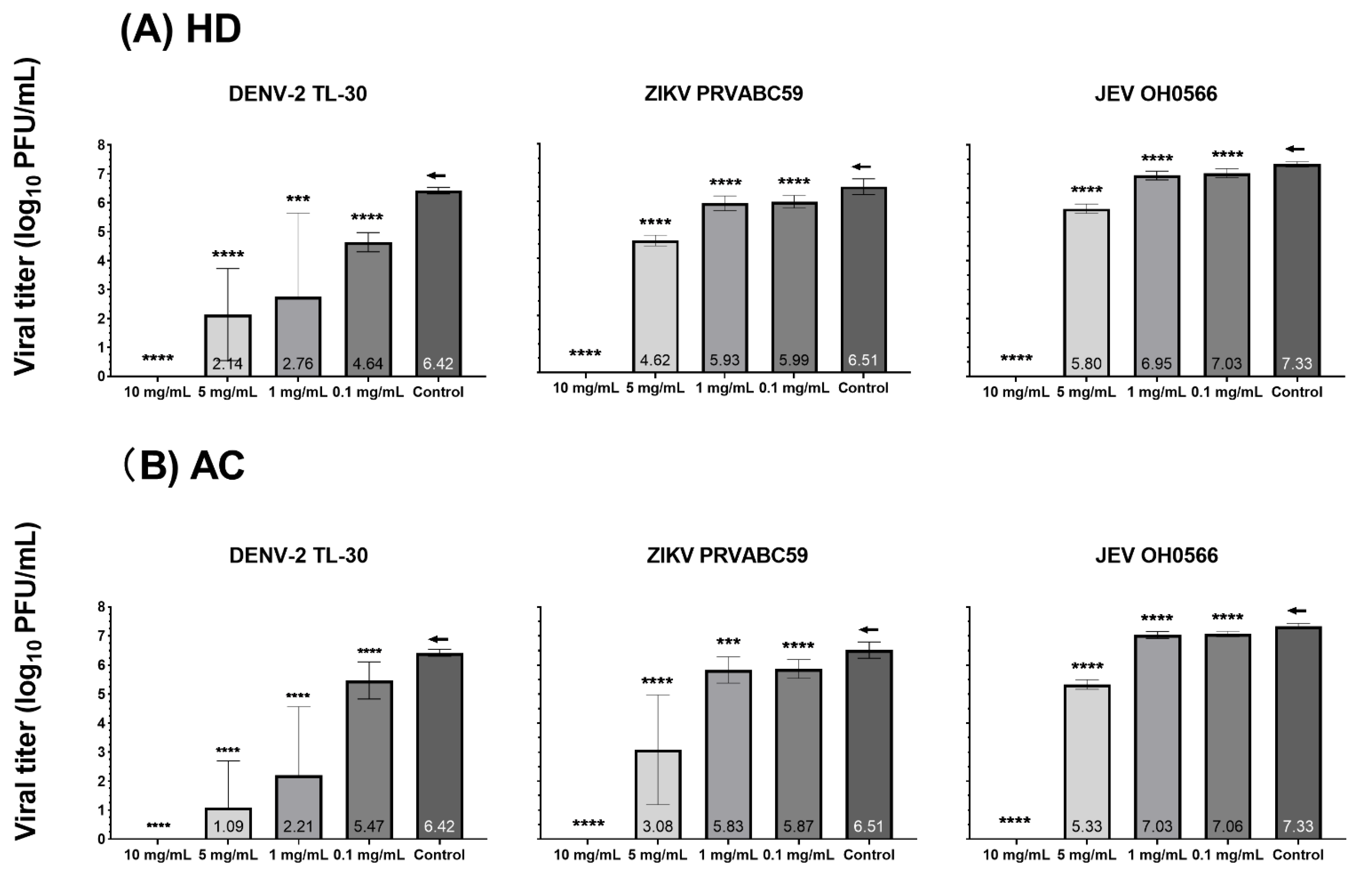
2.6. Plaque Assay
2.7. Antiviral Assay and EC50 Determination
2.8. Viral RNA Extraction
2.9. Real-Time Quantitative Reverse-Transcription Polymerase Chain Reaction
2.10. Time of Drug Addition Assay
2.11. RT2 Profiler PCR Arrays
2.12. SYBR Green Real-Time Quantitative Polymerase Chain Reaction
2.13. Statistical Analysis
3. Results
3.1. Optimizing Multiplicities of Infection (MOI) and Viral Supernatant Harvesting Time Points
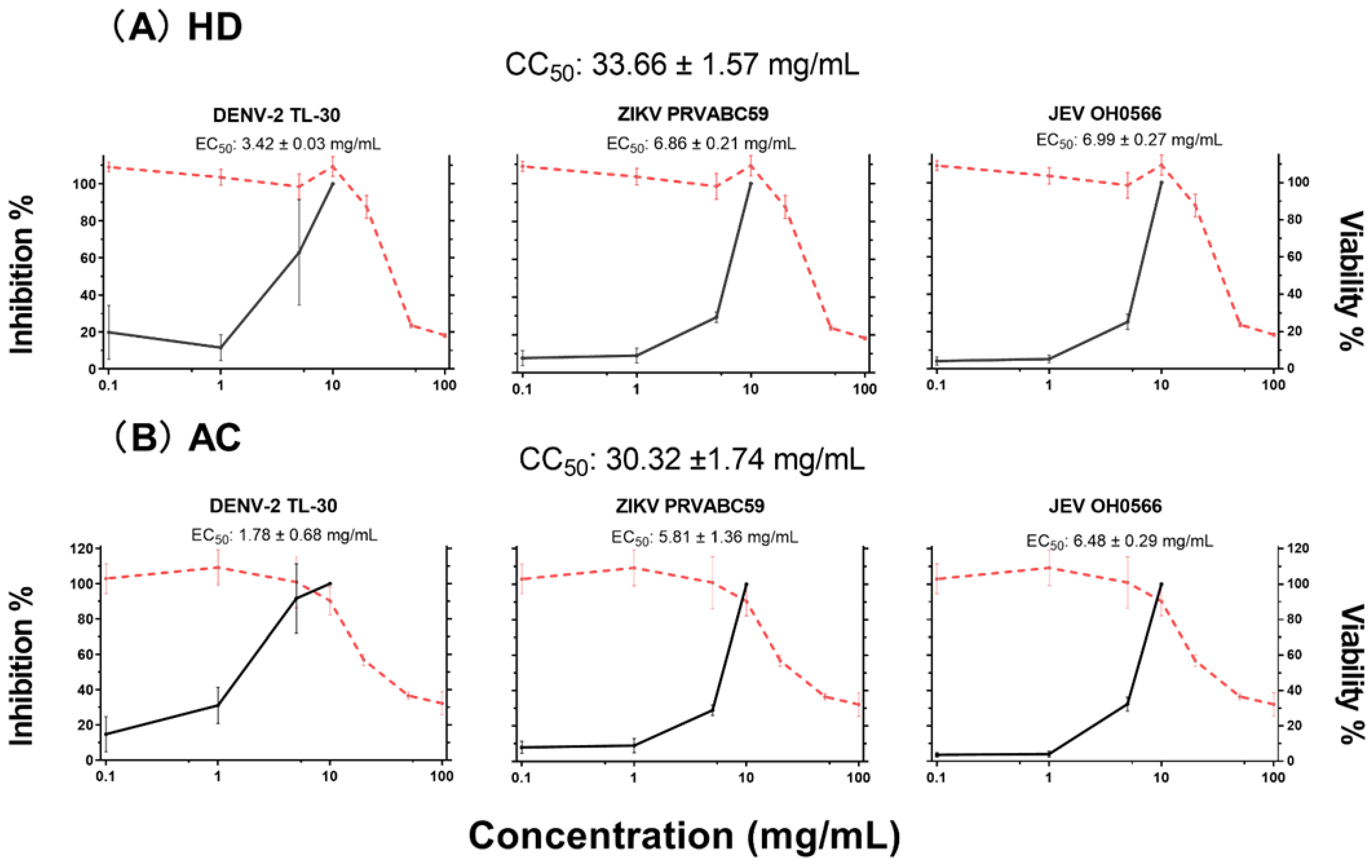
3.2. Antiviral Efficacy and Cytotoxicity in Vero 9013 Cells
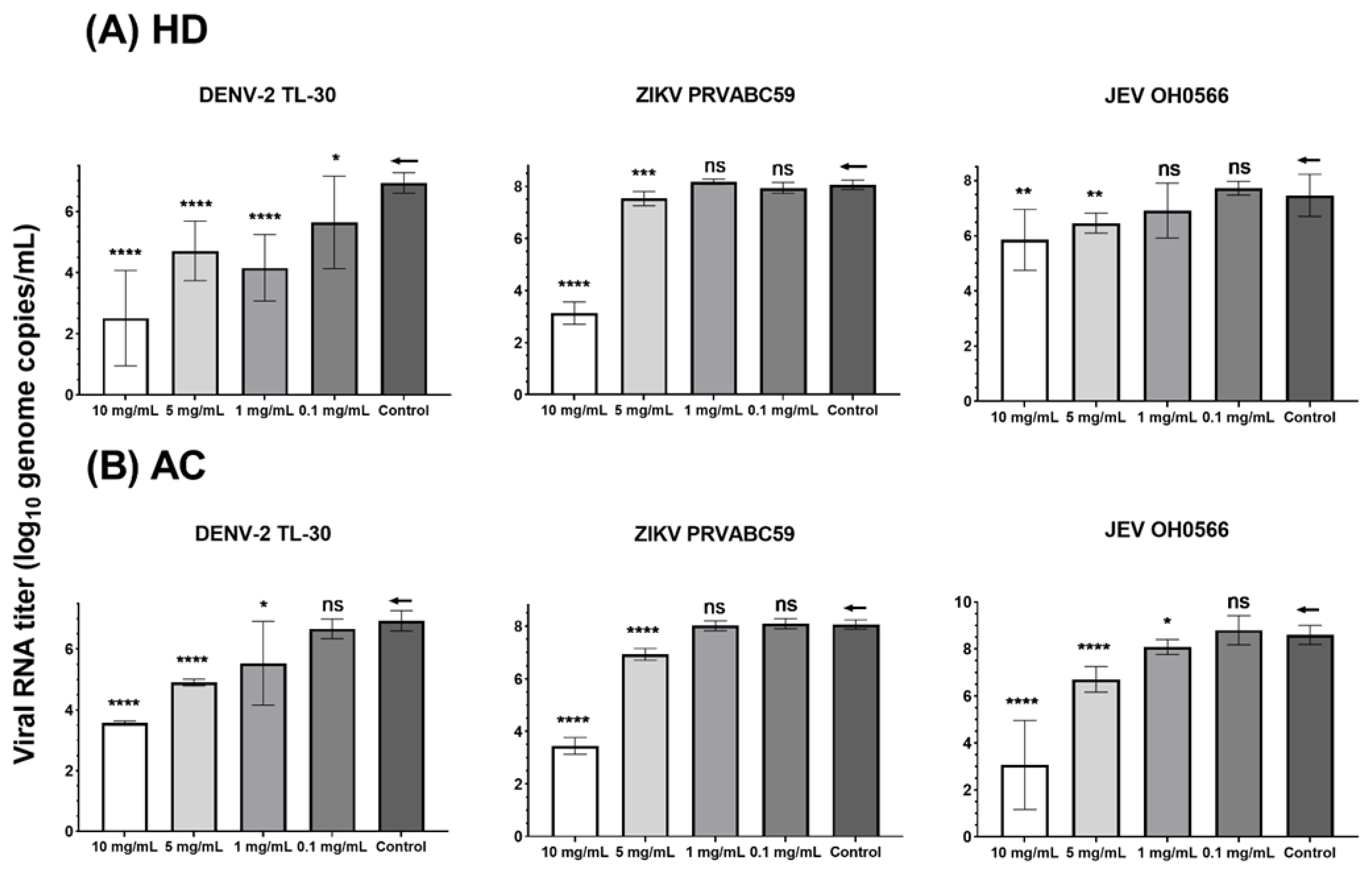
3.3. Quantification of Viral RNA Genome Copy by RT-qPCR
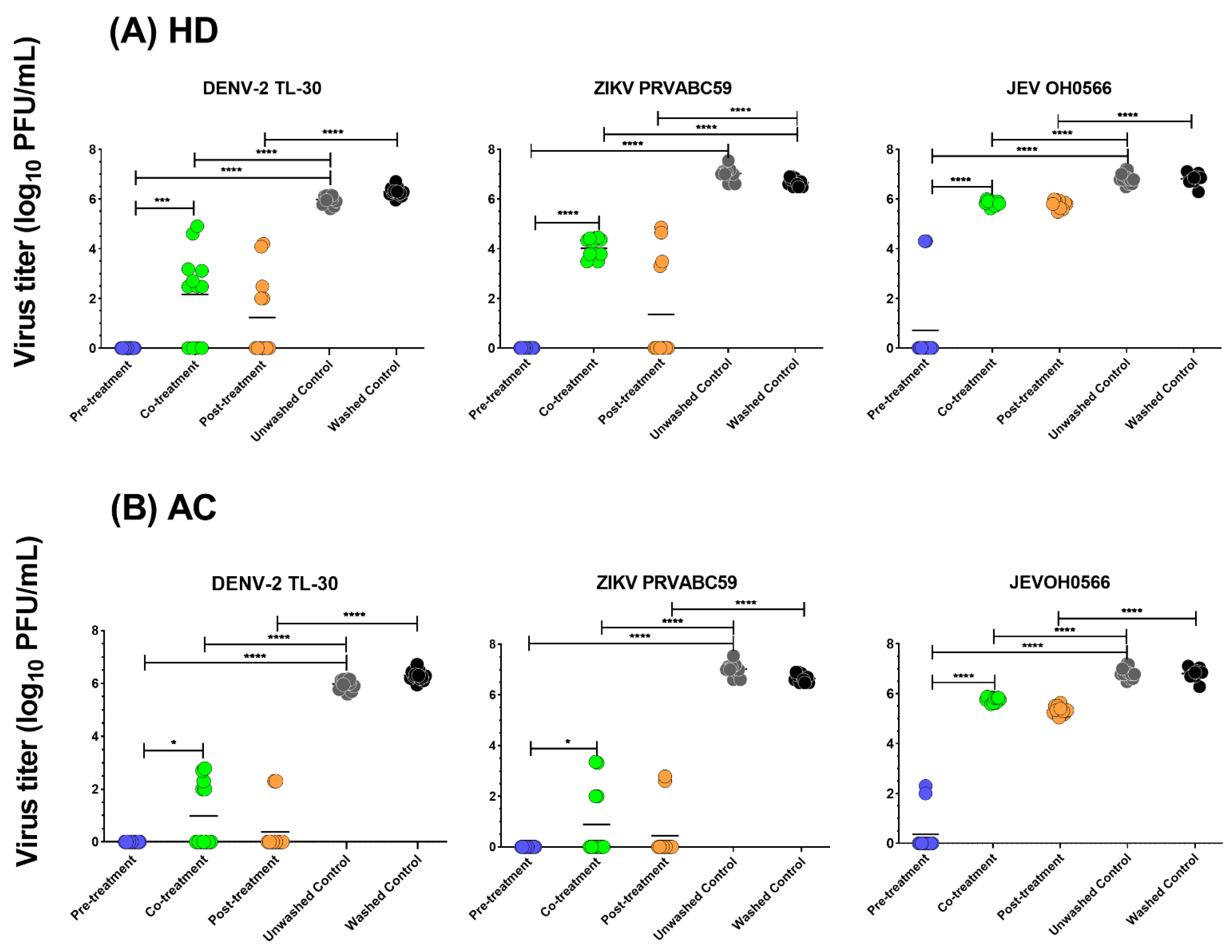
3.4. Time of Drug Addition
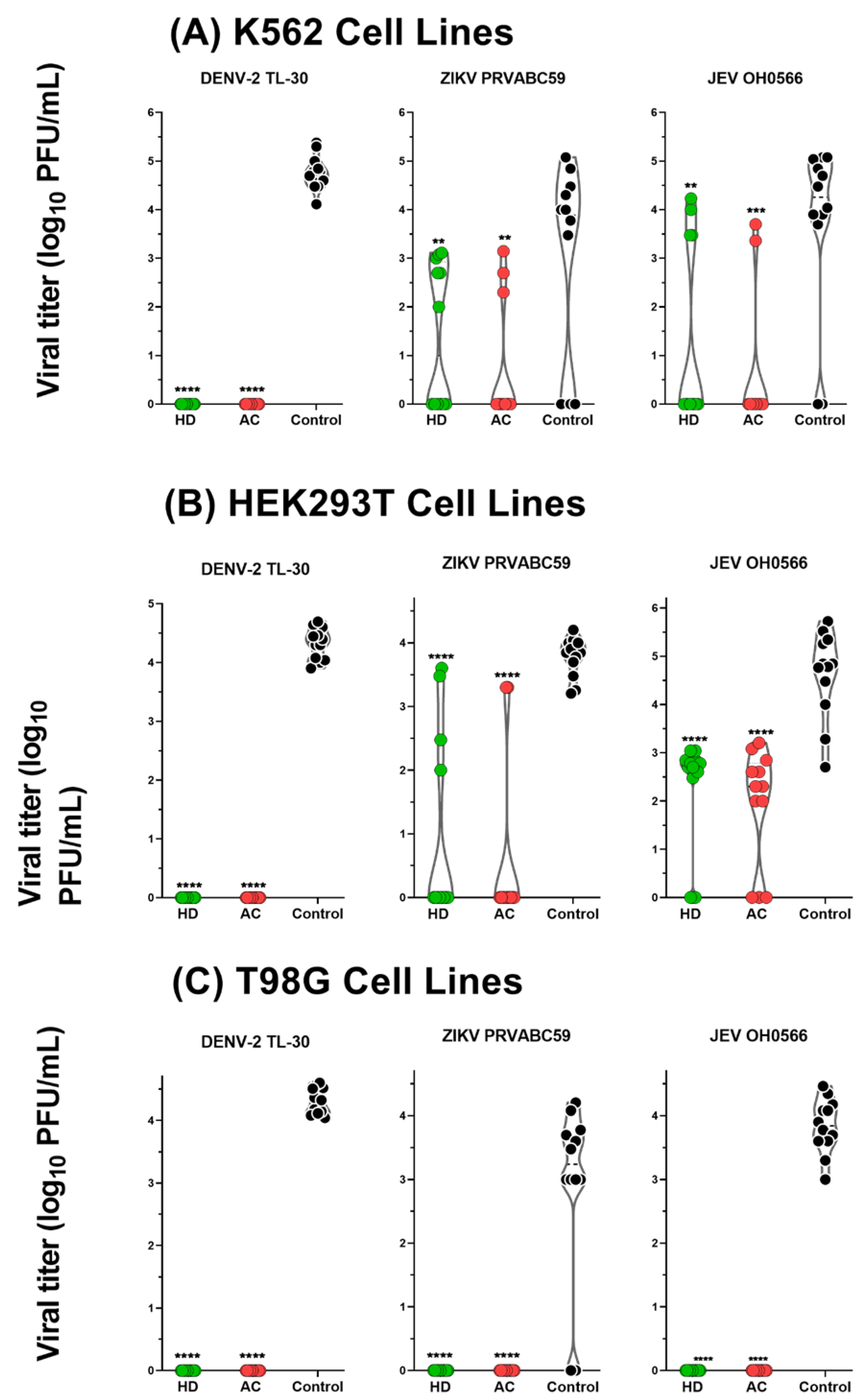
3.5. Antiviral Activity of HD and AC in Human-derived Cell Lines
3.6. Screening of Differentially Expressed Genes (DEGs)
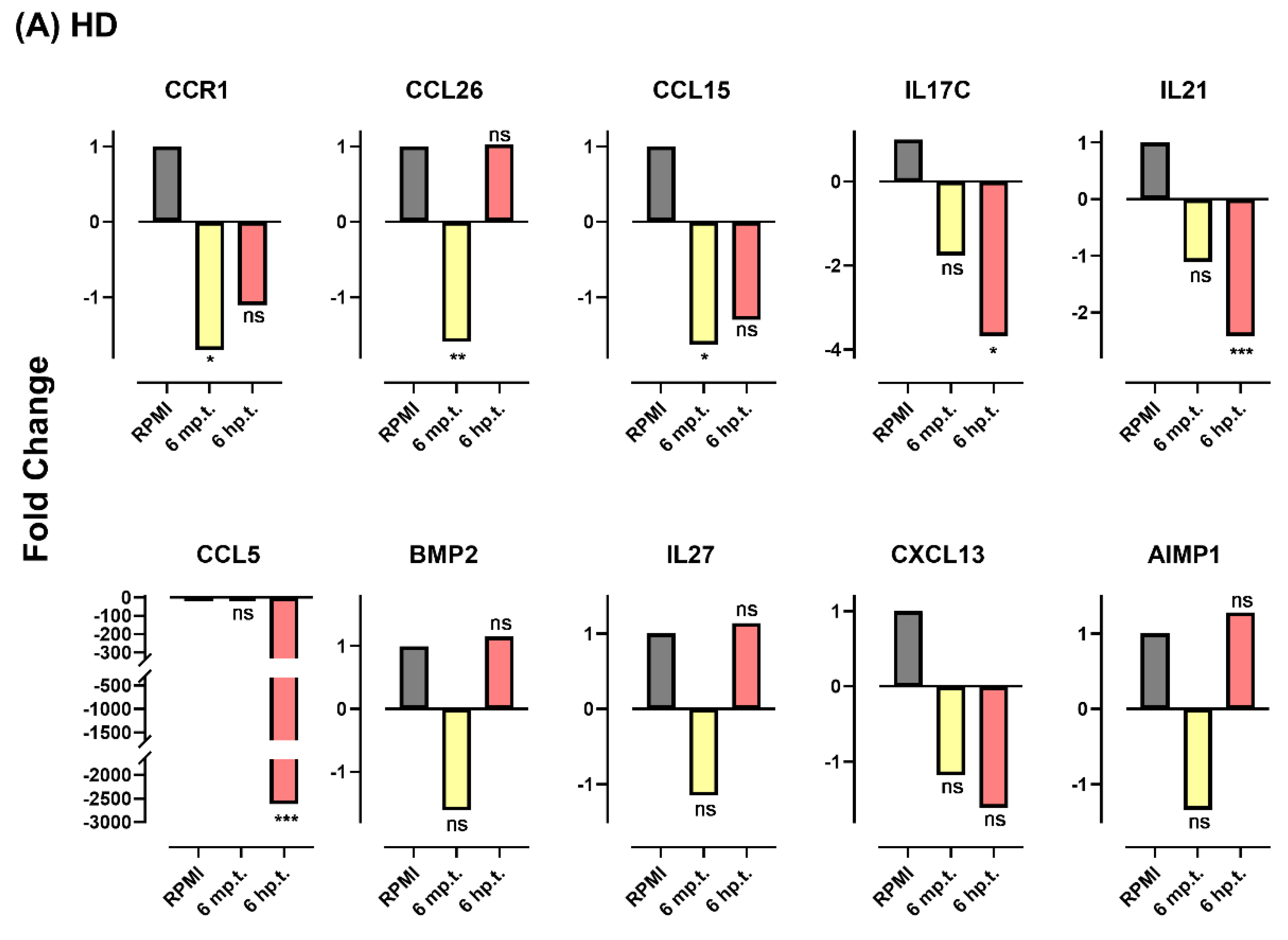
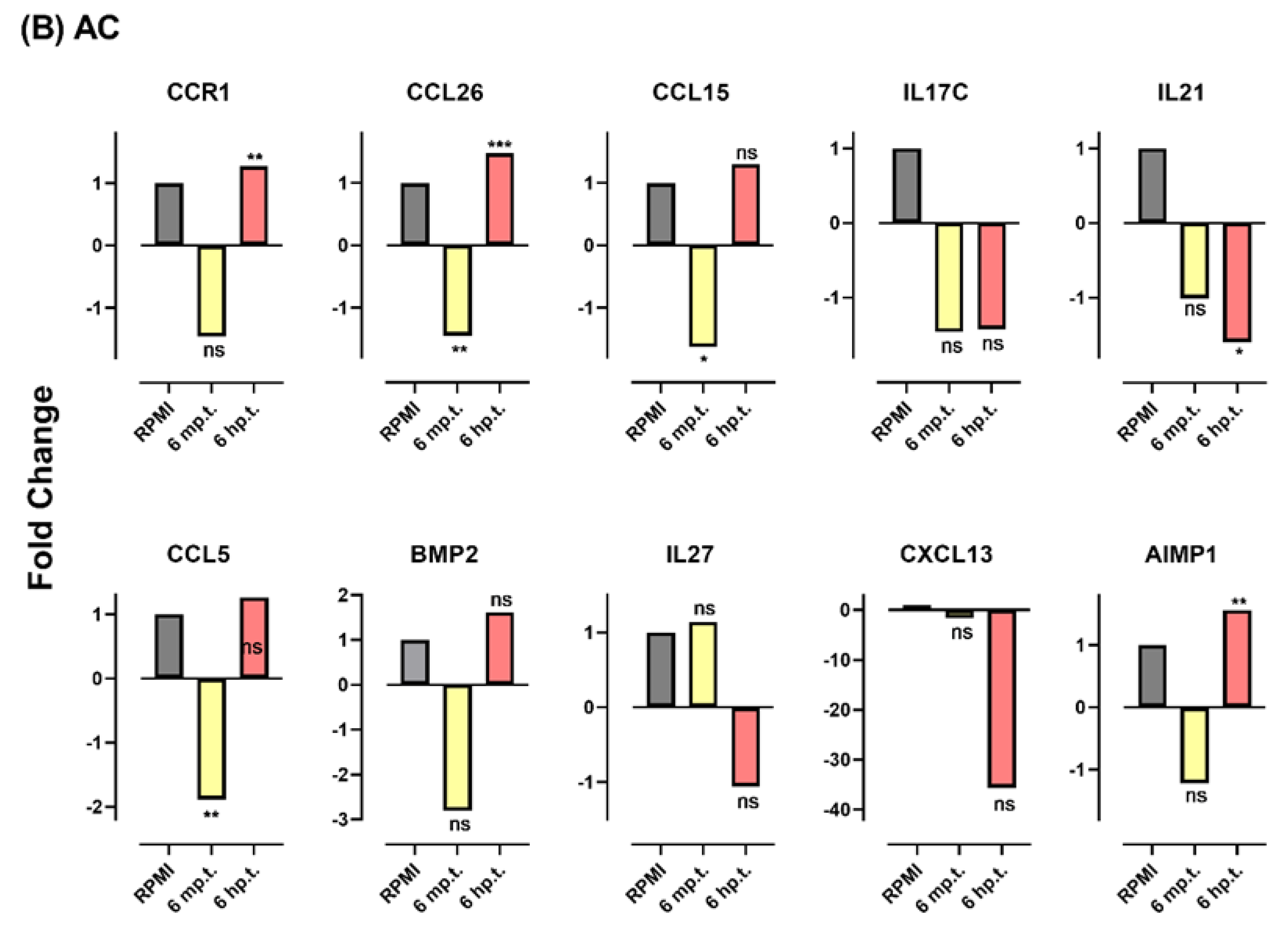
3.7. Herbs Altered Gene Expression Profile Response to Viral Infection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | Artemisia capillaris |
| AIMP1 | Aminoacyl tRNA synthetase complex interacting multifunctional protein 1 |
| BMP2 | Bone morphogenetic protein 2 |
| CC50 | Half maximal cytotoxic concentration |
| CCL15 | C-C motif chemokine ligand 15 |
| CCL26 | C-C motif chemokine ligand 26 |
| CCL5 | C-C motif chemokine ligand 5 |
| CCMG | Concentrated Chinese Medicine Granules |
| CCR1 | C-C motif chemokine receptor 1 |
| CNS | Central Nervous System |
| CXCL13 | C-X-C motif chemokine ligand 13 |
| DEGs | Differentially expressed gene |
| DENV | Dengue virus |
| DMEM | Dulbecco’s Modified Eagle Medium |
| EC50 | Half maximal effective concentration |
| EMEM | Eagle’s minimum essential medium |
| FCS | Fetal calf serum |
| GDC | Genomic DNA control |
| h p.i. | Hour (s) post-infection |
| h p.t. | Hour (s) post-treatment |
| HD | Hedyotis Diffusa |
| HEK-293T cell lines | Human embryonic kidney cell lines |
| HKG | Housekeeping gene |
| IL17C | Interleukin 17C |
| IL21 | Interleukin 21 |
| IL27 | Interleukin 27 |
| JEV | Japanese encephalitis virus |
| K562 cell lines | Human chronic myeloid leukemia cell lines |
| LRV | Log reduction value |
| m p.t. | Minute (s) post the treatment |
| MOI | Multiplicities of infection |
| OD | Optical density |
| PFU | Plaque-forming units |
| PPC | Positive PCR control |
| RPMI | Roswell Park Memorial Institute |
| RT-qPCR | Quantitative real-time polymerase chain reaction |
| SI | Selectivity index |
| SYBR green qPCR | SYBR green real-time quantitative polymerase chain reaction |
| T98G cell lines | Human glioblastoma cell lines |
| TCM | Traditional Chinese medicine |
| ZIKV | Zika virus |
References
- Barnard, T.R.; Abram, Q.H.; Lin, Q.F.; Wang, A.B.; Sagan, S.M. Molecular Determinants of Flavivirus Virion Assembly. Trends Biochem. Sci. 2021, 46, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Pierson, T.C.; Diamond, M.S. The continued threat of emerging flaviviruses. Nat. Microbiol. 2020, 5, 796–812. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.Q.; Fukuta, M.; Balingit, J.C.; Nguyen, T.T.N.; Nguyen, C.T.; Inoue, S.; Nguyen, T.T.T.; Nguyen, L.K.H.; Minakawa, N.; Morita, K.; et al. Direct Viral RNA Detection of SARS-CoV-2 and DENV in Inactivated Samples by Real-Time RT-qPCR: Implications for Diagnosis in Re-source Limited Settings with Flavivirus Co-Circulation. Pathogens 2021, 10, 1558. [Google Scholar] [CrossRef] [PubMed]
- Christy, M.P.; Uekusa, Y.; Gerwick, L.; Gerwick, W.H. Natural Products with Potential to Treat RNA Virus Pathogens Including SARS-CoV-2. J. Nat. Prod. 2021, 84, 161–182. [Google Scholar] [CrossRef] [PubMed]
- Imad, H.A.; Phadungsombat, J.; Nakayama, E.E.; Chatapat, L.; Pisutsan, P.; Matsee, W.; Piyaphanee, W.; Nguitragool, W.; Shioda, T. A Cluster of Dengue Cases in Travelers: A Clinical Series from Thailand. Trop. Med. Infect. Dis. 2021, 6, 152. [Google Scholar] [CrossRef]
- Deng, S.-Q.; Yang, X.; Wei, Y.; Chen, J.-T.; Wang, X.-J.; Peng, H.-J. A Review on Dengue Vaccine Development. Vaccines 2020, 8, 63. [Google Scholar] [CrossRef]
- Mazeaud, C.; Freppel, W.; Chatel-Chaix, L. The Multiples Fates of the Flavivirus RNA Genome During Pathogenesis. Front. Genet. 2018, 9, 595. [Google Scholar] [CrossRef]
- Harapan, H.; Ryan, M.; Yohan, B.; Abidin, R.S.; Nainu, F.; Rakib, A.; Jahan, I.; Emran, T.B.; Ullah, I.; Panta, K.; et al. COVID-19 and dengue: Double punches for dengue-endemic countries in Asia. Rev. Med. Virol. 2020, 31, e2161. [Google Scholar] [CrossRef]
- Aguas, R.; Dorigatti, I.; Coudeville, L.; Luxemburger, C.; Ferguson, N.M. Cross-serotype interactions and disease outcome prediction of dengue infections in Vietnam. Sci. Rep. 2019, 9, 9395. [Google Scholar] [CrossRef]
- Moi, M.L.; Lim, C.K.; Kotaki, A.; Takasaki, T.; Kurane, I. Discrepancy in Dengue Virus Neutralizing Antibody Titers between Plaque Reduction Neutralizing Tests with Fcγ Receptor (FcγR)-Negative and FcγR-Expressing BHK-21 Cells. Clin. Vaccine Immunol. 2010, 17, 402–407. [Google Scholar] [CrossRef] [Green Version]
- Bhatt, P.; Sabeena, S.P.; Varma, M.; Arunkumar, G. Current Understanding of the Pathogenesis of Dengue Virus Infection. Curr. Microbiol. 2021, 78, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Zhang, R.; Cheng, G. Progress towards understanding the pathogenesis of dengue hemorrhagic fever. Virol. Sin. 2017, 32, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Umakanth, M.; Suganthan, N. Unusual Manifestations of Dengue Fever: A Review on Expanded Dengue Syndrome. Cureus 2020, 12, e10678. [Google Scholar] [CrossRef]
- Wang, W.H.; Urbina, A.N.; Chang, M.R.; Assavalapsakul, W.; Lu, P.L.; Chen, Y.H.; Wang, S.F. Dengue hemorrhagic fever—A systemic literature review of current perspectives on pathogenesis, prevention and control. J. Microbiol. Immunol. Infect. 2020, 53, 963–978. [Google Scholar] [CrossRef]
- Mutheneni, S.R.; Morse, A.P.; Caminade, C.; Upadhyayula, S.M. Dengue burden in India: Recent trends and importance of climatic parameters. Emerg. Microbes. Infect. 2017, 6, e70. [Google Scholar] [CrossRef]
- Bhatt, S.; Gething, P.W.; Brady, O.J.; Messina, J.P.; Farlow, A.W.; Moyes, C.L.; Drake, J.M.; Brownstein, J.S.; Hoen, A.G.; Sankoh, O.; et al. The global distribution and burden of dengue. Nature 2013, 496, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Martinez, D.R.; Metz, S.W.; Baric, R.S. Dengue Vaccines: The Promise and Pitfalls of Antibody-Mediated Protection. Cell Host Microbe 2021, 29, 13–22. [Google Scholar] [CrossRef]
- Young, E.; Carnahan, R.H.; Andrade, D.V.; Kose, N.; Nargi, R.S.; Fritch, E.J.; Munt, J.E.; Doyle, M.P.; White, L.; Baric, T.J.; et al. Identification of Dengue Virus Serotype 3 Specific Antigenic Sites Targeted by Neutralizing Human Antibodies. Cell Host Microbe 2020, 27, 710–724.e7. [Google Scholar] [CrossRef]
- Reich, N.G.; Shrestha, S.; King, A.; Rohani, P.; Lessler, J.; Kalayanarooj, S.; Yoon, I.-K.; Gibbons, R.V.; Burke, D.S.; Cummings, D.A.T. Interactions between serotypes of dengue highlight epidemiological impact of cross-immunity. J. R. Soc. Interface 2013, 10, 20130414. [Google Scholar] [CrossRef]
- Brown, M.G.; Hermann, L.L.; Issekutz, A.C.; Marshall, J.S.; Rowter, D.; Al-Afif, A.; Anderson, R. Dengue Virus Infection of Mast Cells Triggers Endothelial Cell Activation. J. Virol. 2011, 85, 1145–1150. [Google Scholar] [CrossRef] [Green Version]
- Modhiran, N.; Watterson, D.; Muller, D.A.; Panetta, A.K.; Sester, D.P.; Liu, L.; Hume, D.A.; Stacey, K.J.; Young, P.R. Dengue virus NS1 protein activates cells via Toll-like receptor 4 and disrupts endothelial cell monolayer integrity. Sci. Transl. Med. 2015, 7, 142–304. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.A.; Pyke, A.T.; Sonja, H.M.; Andrew, D.; Mores, C.N.; Christofferson, R.C.; Gubler, D.J.; Bennett, S.N.; Hurk, A.F. An explosive epidemic of denv-3 in cairns, australia. PLoS ONE. 2013, 8, e68137. [Google Scholar] [CrossRef]
- Shukla, R.; Ramasamy, V.; Shanmugam, R.K.; Ahuja, R.; Khanna, N. Antibody-Dependent Enhancement: A Challenge for Developing a Safe Dengue Vaccine. Front. Cell. Infect. Microbiol. 2020, 10, 572681. [Google Scholar] [CrossRef]
- Song, B.H.; Yun, S.I.; Woolley, M.; Lee, Y.M. Zika virus: History, epidemiology, transmission, and clinical presentation. J Neuroimmunol. 2017, 308, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.C. Microcephaly and Zika virus infection. Lancet 2016, 387, 2070–2072. [Google Scholar] [CrossRef]
- Dick, G.W. Zika virus. II. Pathogenicity and physical properties. Trans. R. Soc. Trop. Med. Hyg. 1952, 46, 521–534. [Google Scholar] [CrossRef]
- Pielnaa, P.; Al-Saadawe, M.; Saro, A.; Dama, M.F.; Zhou, M.; Huang, Y.; Huang, J.; Xia, Z. Zika virus-spread, epidemiology, genome, transmission cycle, clinical manifestation, associated challenges, vaccine and antiviral drug development. Virology 2020, 543, 34–42. [Google Scholar] [CrossRef]
- Basu, R.; Tumban, E. Zika Virus on a Spreading Spree: What we now know that was unknown in the 1950’s. Virol. J. 2016, 13, 165. [Google Scholar] [CrossRef]
- Masmejan, S.; Musso, D.; Vouga, M.; Pomar, L.; Dashraath, P.; Stojanov, M.; Panchaud, A.; Baud, D. Zika Virus. Pathogens 2020, 9, 898. [Google Scholar] [CrossRef]
- Gasco, S.; Muñoz-Fernández, M. A Review on the Current Knowledge on ZIKV Infection and the Interest of Organoids and Nanotechnology on Development of Effective Therapies against Zika Infection. Int. J. Mol. Sci. 2020, 22, 35. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Shi, W.F.; Qin, C.F. The evolution of Zika virus from Asia to the Americas. Nat. Rev. Microbiol. 2019, 17, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.I.; Lee, Y.M. Japanese encephalitis: The virus and vaccines. Hum. Vaccin. Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Barrett, A.D.T. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Turtle, L.; Solomon, T. Japanese encephalitis—The prospects for new treatments. Nat. Rev. Neurol. 2018, 14, 298–313. [Google Scholar] [CrossRef]
- Simon, L.V.; Sandhu, D.S.; Goyal, A.; Kruse, B. Japanese Encephalitis. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022; PMID: 29262148. [Google Scholar]
- Xing, J.; Zhang, Y.; Lin, Z.; Liu, L.; Xu, Q.; Liang, J.; Yuan, Z.; Huang, C.; Liao, M.; Qi, W. 3′UTR SL-IV and DB1 Regions Contribute to Japanese Encephalitis Virus Replication and Pathogenicity. Front. Veter Sci. 2021, 8, 703147. [Google Scholar] [CrossRef]
- Mileno, M.D. Japanese Encephalitis Vaccine. R. I. Med. J. 2020, 103, 49–50. [Google Scholar]
- Connor, B.A.; Hamer, D.H.; Kozarsky, P.; Jong, E.; Halstead, S.B.; Key Stone, J.; Mileno, M.D.; Dawood, R.; Bonnie Rogers, B.; Bunn, W.B. Japanese encephalitis vaccine for travelers: Risk-benefit recon sidered. J. Travel Med. 2019, 26, taz037. [Google Scholar] [CrossRef]
- Chong, H.Y.; Leow, C.Y.; Abdul Majeed, A.B.; Leow, C.H. Flavivirus infection-A review of immunopathogenesis, immunological response, and immunodiagnosis. Virus Res. 2019, 274, 197770. [Google Scholar] [CrossRef]
- Kok, W.M. New developments in flavivirus drug discovery. Expert Opin. Drug Discov. 2016, 11, 433–445. [Google Scholar] [CrossRef]
- Liu, M.; Chen, X. COVID-19 basics and vaccine development with a Canadian perspective. Can. J. Microbiol. 2021, 67, 112–118. [Google Scholar] [CrossRef]
- Blázquez, A.-B.; Saiz, J.-C. Potential for Protein Kinase Pharmacological Regulation in Flaviviridae Infections. Int. J. Mol. Sci. 2020, 21, 9524. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Lin, J.; Wei, L.; Zhang, L.; Wang, L.; Zhan, Y.; Zeng, J.; Xu, W.; Shen, A.; Hong, Z.; et al. Hedyotis diffusa Willd Inhibits Colorectal Cancer Growth in Vivo via Inhibition of STAT3 Signaling Pathway. Int. J. Mol. Sci. 2012, 13, 6117–6128. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-J.; He, J.; Li, G.-Z.; Fang, P.-P.; Xie, J.-D.; Ding, Y.-W.; Mao, Y.-Q.; Hu, K.-F. Analyzing hedyotis diffusa mechanisms of action from the genomics perspective. Comput. Methods Programs Biomed. 2019, 174, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xin, P.; Wang, Y.; Zhou, X.; Wei, D.; Deng, C.; Sun, S. Iridoids and sfingolipids from Hedyotis diffusa. Fitoterapia 2018, 124, 152–159. [Google Scholar] [CrossRef]
- He, C.-S.; Yue, H.-Y.; Xu, J.; Xue, F.; Liu, J.; Li, Y.-Y.; Jing, H.-E. Protective effects of Artemisia capillarispolysaccharide on oxidative injury to the liver in rats with obstructive jaundice. Exp. Ther. Med. 2012, 4, 645–648. [Google Scholar] [CrossRef]
- Balingit, J.C.; Ly, M.H.P.; Matsuda, M.; Suzuki, R.; Hasebe, F.; Morita, K.; Moi, M.L. A Simple and High-Throughput ELISA-Based Neutralization Assay for the Determination of Anti-Flavivirus Neutralizing Antibodies. Vaccines 2020, 8, 297. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Takasaki, T.; Yamada, K.-I.; Nerome, R.; Tajima, S.; Kurane, I. Development and Evaluation of Fluorogenic TaqMan Reverse Transcriptase PCR Assays for Detection of Dengue Virus Types 1 to 4. J. Clin. Microbiol. 2004, 42, 5935–5937. [Google Scholar] [CrossRef] [PubMed]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and Serologic Properties of Zika Virus Associated with an Epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Maeki, T.; Tajima, S.; Kyaw, A.K.; Matsumoto, F.; Miura, K.; Yamashita, A.; Yoshikawa, A.; Negishi, K.; Noguchi, Y.; Tadokoro, K.; et al. Comparison of Neutralizing Antibody Titers against Japanese Encephalitis Virus Genotype V Strain with Those against Genotype I and III Strains in the Sera of Japanese Encephalitis Patients in Japan in 2016. Jpn. J. Infect. Dis. 2018, 71, 360–364. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef]
- Osada, N.; Kohara, A.; Yamaji, T.; Hirayama, N.; Kasai, F.; Sekizuka, T.; Kuroda, M.; Hanada, K. The Genome Landscape of the African Green Monkey Kidney-Derived Vero Cell Line. DNA Res. 2014, 21, 673–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okemoto-Nakamura, Y.; Someya, K.; Yamaji, T.; Saito, K.; Takeda, M.; Hanada, K. Poliovirus-nonsusceptible Vero cell line for the World Health Organization global action plan. Sci. Rep. 2021, 11, 6746. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Salehi, B.; Schnitzler, P.; Ayatollahi, S.A.; Kobarfard, F.; Fathi, M.; Eisazadeh, M.; Sharifi-Rad, M. Susceptibility of herpes simplex virus type 1 to monoterpenes thymol, carvacrol, p-cymene and essential oils of Sinapis arvensis L.; Lallemantia royleana Benth. and Pulicaria vulgaris Gaertn. Cell. Mol. Biol. 2017, 63, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Amoros, M.; Simõs, C.M.O.; Girre, L.; Sauvager, F.; Cormier, M. Synergistic Effect of Flavones and Flavonols Against Herpes Simplex Virus Type 1 in Cell Culture. Comparison with the Antiviral Activity of Propolis. J. Nat. Prod. 1992, 55, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Nelson, A.R.; Betsholtz, C.; Zlokovic, B.V. Establishment and Dysfunction of the Blood-Brain Barrier. Cell 2015, 163, 1064–1078. [Google Scholar] [CrossRef]
- Banks, W.A.; Kovac, A.; Morofuji, Y. Neurovascular unit crosstalk: Pericytes and astrocytes modify cytokine secretion patterns of brain endothelial cells. J. Cereb. Blood Flow. Metab. 2018, 38, 1104–1118. [Google Scholar] [CrossRef]
- Suzuki, A.; Stern, S.A.; Bozdagi, O.; Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-Neuron Lactate Transport Is Required for Long-Term Memory Formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef]
- Habbas, S.; Santello, M.; Becker, D.; Stubbe, H.; Zappia, G.; Liaudet, N.; Klaus, F.R.; Kollias, G.; Fontana, A.; Pryce, C.R.; et al. Neuroinflammatory TNFα Impairs Memory via Astrocyte Signaling. Cell 2015, 163, 1730–1741. [Google Scholar] [CrossRef]
- Marinho, P.E.S.; De Oliveira, D.B.; Candiani, T.M.S.; Crispim, A.P.C.; Alvarenga, P.P.M.; Castro, F.C.D.S.; Abrahão, J.S.; Rios, M.; Coimbra, R.S.; Kroon, E.G. Meningitis Associated with Simultaneous Infection by Multiple Dengue Virus Serotypes in Children, Brazil. Emerg. Infect. Dis. 2017, 23, 115–118. [Google Scholar] [CrossRef]
- Lossia, O.V.; Conway, M.J.; Tree, M.O.; Williams, R.J.; Goldthorpe, S.C.; Srinageshwar, B.; Dunbar, G.L.; Rossignol, J. Zika virus induces astrocyte differentiation in neural stem cells. J. NeuroVirology 2017, 24, 52–61. [Google Scholar] [CrossRef]
- Jorgačevski, J.; Korva, M.; Potokar, M.; Lisjak, M.; Avšič-Županc, T.; Zorec, R. ZIKV Strains Differentially Affect Survival of Human Fetal Astrocytes versus Neurons and Traffic of ZIKV-Laden Endocytotic Compartments. Sci. Rep. 2019, 9, 8069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patabendige, A.; Michael, B.; Craig, A.; Solomon, T. Brain microvascular endothelial-astrocyte cell responses following Japanese encephalitis virus infection in an in vitro human blood-brain barrier model. Mol. Cell. Neurosci. 2018, 89, 60–70. [Google Scholar] [CrossRef] [PubMed]
- Baez-Jurado, E.; Vega, G.G.; Aliev, G.; Tarasov, V.V.; Esquinas, P.; Echeverria, V.; Barreto, G.E. Blockade of Neuroglobin Reduces Protection of Conditioned Medium from Human Mesenchymal Stem Cells in Human Astrocyte Model (T98G) Under a Scratch Assay. Mol. Neurobiol. 2018, 55, 2285–2300. [Google Scholar] [CrossRef] [PubMed]
- Ponia, S.S.; Robertson, S.J.; McNally, K.L.; Subramanian, G.; Sturdevant, G.L.; Lewis, M.; Jessop, F.; Kendall, C.; Gallegos, D.; Hay, A.; et al. Mitophagy antagonism by ZIKV reveals Ajuba as a regulator of PINK1 signaling, PKR-dependent inflammation, and viral invasion of tissues. Cell Rep. 2021, 37, 109888. [Google Scholar] [CrossRef] [PubMed]
- Hassert, M.; Steffen, T.L.; Scroggins, S.; Coleman, A.K.; Shacham, E.; Brien, J.D.; Pinto, A.K. Prior Heterologous Flavivirus Exposure Results in Reduced Pathogenesis in a Mouse Model of Zika Virus Infection. J. Virol. 2021, 95, e00573-21. [Google Scholar] [CrossRef]
- Glass, W.G.; Hickey, M.J.; Hardison, J.L.; Liu, M.T.; Manning, J.E.; Lane, T.E. Antibody Targeting of the CC Chemokine Ligand 5 Results in Diminished Leukocyte Infiltration into the Central Nervous System and Reduced Neurologic Disease in a Viral Model of Multiple Sclerosis. J. Immunol. 2004, 172, 4018–4025. [Google Scholar] [CrossRef]
- Miller, A.L.; Gerard, C.; Schaller, M.; Gruber, A.D.; Humbles, A.A.; Lukacs, N.W. Deletion of CCR1 Attenuates Pathophysiologic Responses during Respiratory Syncytial Virus Infection. J. Immunol. 2006, 176, 2562–2567. [Google Scholar] [CrossRef]
- Liu, L.-Z.; Zhang, Z.; Zheng, B.-H.; Shi, Y.; Duan, M.; Ma, L.-J.; Wang, Z.-C.; Dong, L.-Q.; Dong, P.-P.; Shi, J.-Y.; et al. CCL15 Recruits Suppressive Monocytes to Facilitate Immune Escape and Disease Progression in Hepatocellular Carcinoma. Hepatology 2019, 69, 143–159. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Zhao, T.; Cui, R.; Hu, L.; Chang, L.; Wei, X.; Zhang, J.; Li, Y. Hepatitis B Virus X Protein Modulates Chemokine CCL15 Upregulation in Hepatocellular Carcinoma. Anti Cancer Agents Med. Chem. 2021, 21, 2198–2203. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus Strain | HD | CA | |||||
|---|---|---|---|---|---|---|---|
| Vero9013 Cells | CC50 (mg/mL) | SI | Vero9013 Cells | CC50 (mg/mL) | SI | ||
| EC50 (mg/mL) | EC50 (mg/mL) | ||||||
| JEV OH0566 | 7.0 ± 0.3 | 33.7 ± 1.6 | 4.8 | 6.5 ± 0.3 | 30.3 ±1.7 | 4.7 | |
| ZIKV PRVABC59 | 6.9 ± 0.2 | 4.9 | 5.8 ± 1.4 | 5.2 | |||
| DENV-2 TL-30 | 3.4 ± 0.03 | 9.8 | 1.8 ± 0.7 | 18.9 | |||
| Concentration | HD | AC | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| JEV OH0566 | ZIKV PRVABC59 | DENV-2 TL-30 | JEV OH0566 | ZIKV PRVABC59 | DENV-2 TL-30 | ||||||||||||
| RNA Titer | p Value | RNA Titer | p Value | RNA Titer | p Value | RNA Titer | p Value | RNA Titer | p Value | RNA Titer | p Value | ||||||
| 10 mg/mL | 1.6 ± 1.1 | 0.004 | 4.9 ± 0.4 | <0.0001 | 4.4 ± 1.6 | <0.0001 | 5.5 ± 1.9 | <0.0001 | 4.6 ± 0.31 | <0.0001 | 3.3 ± 0.06 | <0.0001 | |||||
| 5 mg/mL | 1.0 ± 0.3 | 0.005 | 0.5 ± 0.3 | 0.0004 | 2.2 ± 1.0 | <0.0001 | 1.9 ± 0.5 | <0.0001 | 1.1 ± 0.22 | <0.0001 | 2.0 ± 0.1 | <0.0001 | |||||
| 1 mg/mL | NS ** | NS | 2.8 ± 1.1 | <0.0001 | 0.9 ± 0.5 | 0.01 | NS | 1.4 ± 1.4 | 0.02 | ||||||||
| 0.1 mg/mL | NS | NS | 1.3 ± 1.5 | 0.0337 | NS | NS | NS | ||||||||||
| HD | AC | ||||||
|---|---|---|---|---|---|---|---|
| Pre-Treatment | Co-Treatment | Post-Treatment | Pre-Treatment | Co-Treatment | Post-Treatment | ||
| DENV-2 TL-30 | 100.0 ± 0.0 | 63.8 ± 29.7 | 80.4 ± 26.4 | 100.0 ± 0.0 | 83.6 ± 20.6 | 93.9 ± 14.2 | |
| ZIKV PRVABC59 | 100.0 ± 0.0 | 42.8 ± 5.6 | 79.6 ± 30.8 | 100.0 ± 0.0 | 87.3 ± 19.6 | 93.3 ± 15.8 | |
| JEV OH0566 | 89.5 ± 24.5 | 14.5 ± 1.5 | 14.8 ± 2.4 | 94.8 ± 12.3 | 15.5 ± 1.5 | 21.7 ± 2.4 | |
| Symbol | Accession Number | Entrez Gene Name | Treatment Time | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 m p.t. | 6 h p.t. | 6 m p.t. | 6 h p.t. | ||||||||||
| Fold Change | p Value | Fold Change | p Value | Fold Change | p Value | Fold Change | p Value | ||||||
| BMP2 | NM_001200 | Bone morphogenetic protein 2 | NC * | NC | NC | NC | |||||||
| IL17C | NM_013278 | Interleukin 17C | NC | −3.7 | 0.03 | NC | NC | ||||||
| IL27 | NM_145659 | Interleukin 27 | NC | NC | NC | NC | |||||||
| IL21 | NM_021803 | Interleukin 21 | NC | −2.4 | 0.0002 | NC | −1.59 | 0.01 | |||||
| CCL5 | NM_002985 | C-C motif chemokine ligand 5 | NC | −2615.2 | 0.0006 | −1.89 | 0.002 | NC | |||||
| CCR1 | NM_001295 | C-C motif chemokine receptor 1 | −1.7 | 0.015 | NC | NC | +1.27 | 0.008 | |||||
| CXCL13 | NM_006419 | C-X-C motif chemokine ligand 13 | NC | NC | NC | NC | |||||||
| AIMP1 | NM_004757 | Aminoacyl tRNA synthetase complex interacting multifunctional protein 1 | NC | NC | NC | +1.6 | 0.001 | ||||||
| CCL26 | NM_006072 | C-C motif chemokine ligand 26 | −1.59 | 0.004 | NC | −1.45 | 0.004 | +1.5 | 0.0003 | ||||
| CCL15 | NM_032964 | C-C motif chemokine ligand 15 | −1.63 | 0.02 | NC | −1.63 | 0.02 | NC | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, Z.Q.; Minakawa, N.; Moi, M.L. Novel Antiviral Efficacy of Hedyotis diffusa and Artemisia capillaris Extracts against Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Infection and Immunoregulatory Cytokine Signatures. Plants 2022, 11, 2589. https://doi.org/10.3390/plants11192589
Mao ZQ, Minakawa N, Moi ML. Novel Antiviral Efficacy of Hedyotis diffusa and Artemisia capillaris Extracts against Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Infection and Immunoregulatory Cytokine Signatures. Plants. 2022; 11(19):2589. https://doi.org/10.3390/plants11192589
Chicago/Turabian StyleMao, Zhan Qiu, Noboru Minakawa, and Meng Ling Moi. 2022. "Novel Antiviral Efficacy of Hedyotis diffusa and Artemisia capillaris Extracts against Dengue Virus, Japanese Encephalitis Virus, and Zika Virus Infection and Immunoregulatory Cytokine Signatures" Plants 11, no. 19: 2589. https://doi.org/10.3390/plants11192589