

The Diversity of Fungi Involved in Damage to Japanese Quince
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
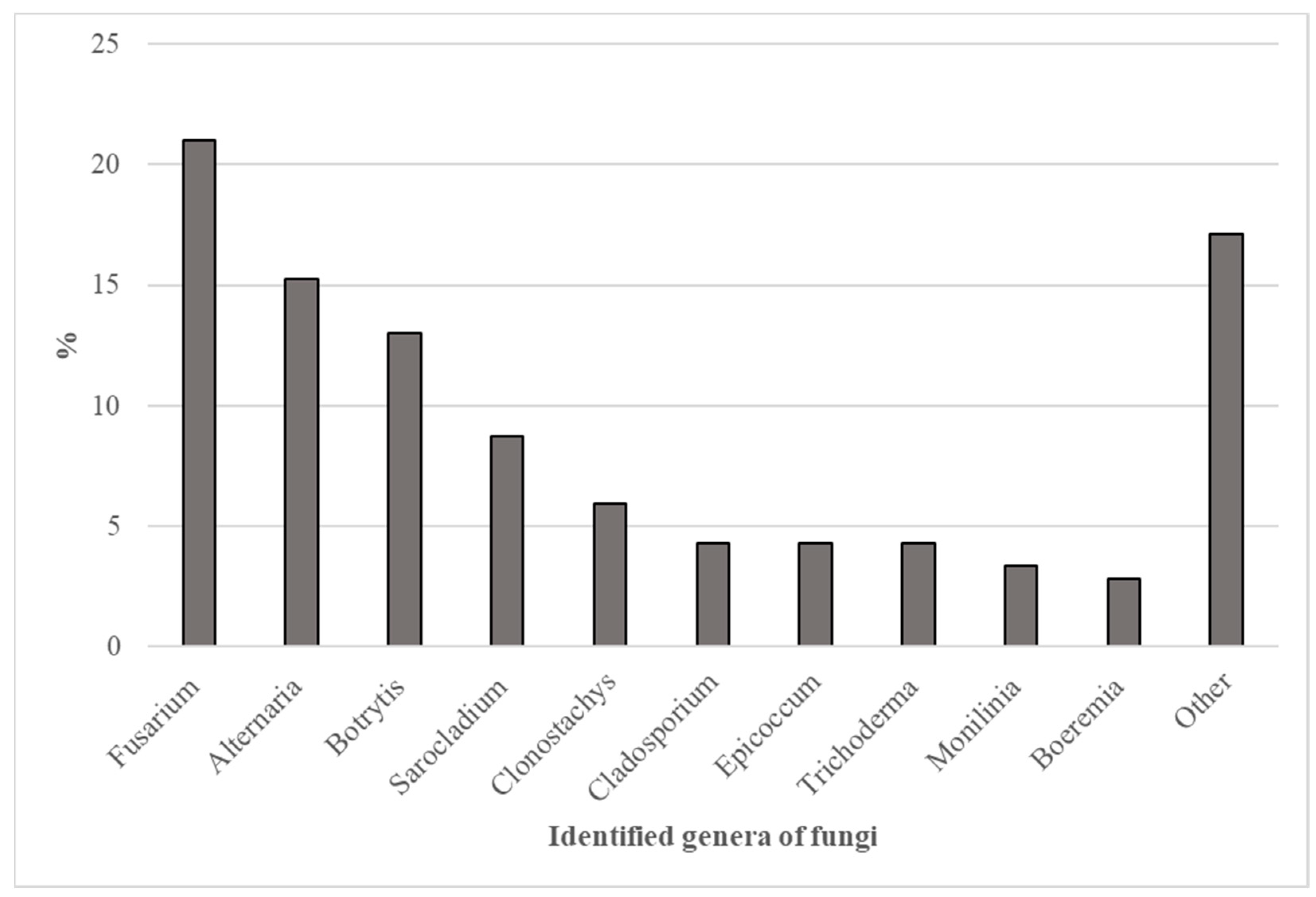
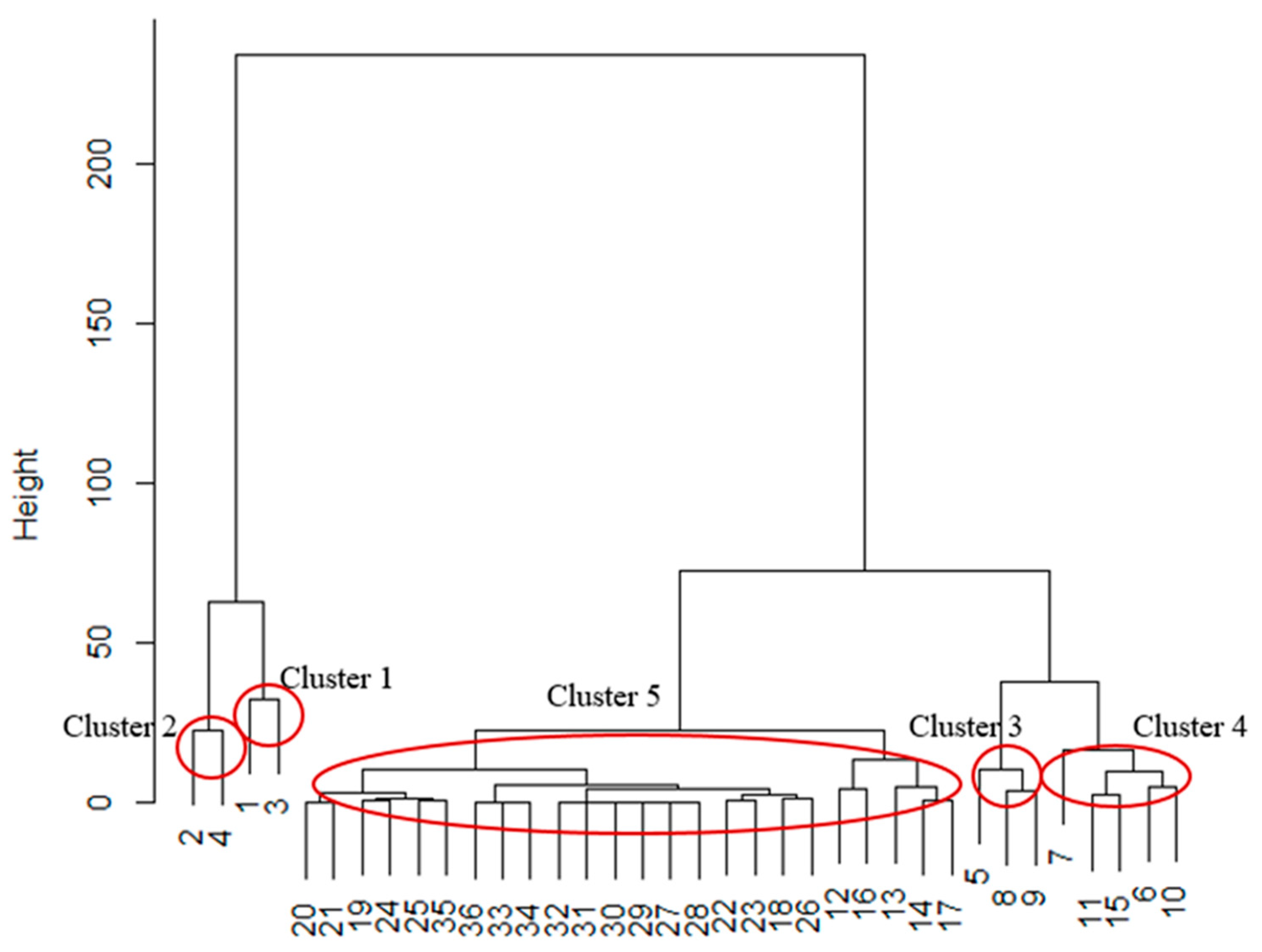
2. Results
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mierina, I.; Serzane, R.; Strele, M.; Moskaluka, J.; Seglina, D.; Jure, M. Extracts of Japanese quince seeds -potential source of antioxidants. In Proceedings of the 6th Baltic Conference on Food Science and Technology FOODBALT-2011. Innovations for Food Science and Production, Jelgava, Latvia, 5–6 May 2011; Faculty of Food Technology, Latvia University of Agriculture: Jelgava, Latvia, 2011; pp. 98–103. [Google Scholar]
- Baranowska-Bosiacka, I.; Bosiacka, B.; Rast, J.; Gutowska, I.; Wolska, J.; Rębacz-Maron, E.; Dębia, K.; Janda, K.; Korbecki, J.; Chlubek, D. Macro- and Microelement Content and Other Properties of Chaenomeles japonica L. Fruit and Protective Effects of Its Aqueous Extract on Hepatocyte Metabolism. Biol. Trace Elem. Res. 2017, 178, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Nahorska, A.; Dzwoniarska, M.; Thiem, B. Fruits of Japanese quince (Chaenomeles japonica (Thunb.) Lindl. Ex Spach) as a source of bioactive compounds. Postępy Fitoter. 2014, 4, 239–246. [Google Scholar]
- Kaufmane, E.; Skrivele, M.; Rubauskis, E.; Strautiņa, S.; Ikase, L.; Lacis, G.; Segliņa, D.; Moročko-Bičevska, I.; Ruisa, S.; Priekule, I. Development of fruit science in Latvia. Proc. Latv. Acad. Sci. Sect. B Nat. Exact. Appl. Sci. 2013, 67, 71–83. [Google Scholar] [CrossRef]
- Rumpunen, K. Chaenomeles: Potential New Fruit Crop for Northern Europe. In Trends in New Crops and New Uses; Whipkey, J., Janick, A., Eds.; ASHS Press: Alexandria, VA, USA, 2002; pp. 385–392. [Google Scholar]
- Repnau, A. Jaapani Ebaküdoonia (Chaenomeles japonica Thunb.) Erinevate Sortide Kasvatamise Perspektiivsus Eesti Tingimustes. Master’s Thesis, Estonian University of Life Sciences, Tartu, Estonia, 2020. [Google Scholar]
- Rumpunen, K.; Trajkovski, V.; Bartish, I.; Garkava, L.; Nybom, H.; Laencina, J.; Ros, J.M.; Jordan, M.J.; Hellin, P.; Tigerstedt, P.M.A.; et al. Domestication of Japanese quince (Chaenomeles japonica). Acta Hortic. 2000, 538, 345–348. [Google Scholar] [CrossRef]
- Bieniasz, M.; Dziedzic, E.; Kaczmarczyk, E. The Effect of storage and Pprocessing on vitamin C content in Japanese quince fruit. Folia Hortic. 2017, 29, 83–93. [Google Scholar] [CrossRef]
- Mihova, T.; Kondakova, V.; Mondeshka, P. Investigations of Chaenomeles japonica (Thunb.) Lindl. in the Region of Central Balkans. Banat. J. Biotechnol. 2012, 2, 43–48. [Google Scholar] [CrossRef]
- Клименкo, C.B.; Недвига, O.H. Хенoмелес: интрoдукция, сoстoяние и перспективы культуры. Інтрoдукція рoслин 1999, 3–4, 125–134. [Google Scholar]
- Panteev, A.V.; Batchilo, A.I.; Grakovich, Z.V. The breeding of Chaenomeles japonica Lindl. (Japanese quince), Cerasus tomentosa Wall. (Felt cherry), and Viburnum opulus L. (Snowball tree) in the Republic of Belarus. Acta Hortic. 1995, 390, 133–136. [Google Scholar] [CrossRef]
- Fedulova, Y.A.; Skripnikova, M.K.; Mezhenskiĭ, V.N. Biological features and economic value of Japanese quince. Sadovod. Vinograd. 2009, 1, 2–3. [Google Scholar]
- Фирсoва, C.B.; Русинoв, A.A. Изучение Айвы япoнскoй в Кирoвскoй oбласти. Аграрная Наука Еврo-Северo-Вoстoка 2013, 1, 19–22. [Google Scholar]
- Jakobija, I.; Bankina, B. Incidence of fruit rot on Japanese quince (Chaenomeles japonica) in Latvia. Res. Rural Dev. 2018, 2, 83–89. [Google Scholar] [CrossRef]
- Jakobija, I.; Bankina, B.; Klūga, A. Leaf spot diseases as an emerging problem in Chaenomeles japonica plantations. In Proceedings of the 3rd International Scientific Virtual Conference, AgroEco 2020 Programme and Abstracts, Kaunas, Lithuania, 2–3 December 2020; Vytautas Magnus University: Lithuania, Kaunas, 2020; p. 27. [Google Scholar]
- Norin, I.; Rumpunen, K. Pathogens on Japanese quince (Chaenomeles japonica) plants. In Japanese Quince—Potential Fruit Crop for Northern Europe.; Rumpunen, K., Ed.; Department of Crop Science, Swedish University of Agricultural Sciences: Uppsala, Sweden, 2003; pp. 37–54. [Google Scholar]
- Grigaliūnaitė, B.; Žilinskaitė, S.; Radaitienė, D. Japoninio svarainio (Chaenomeles japonica) fitosanitarinė būklė Vilniaus Universiteto Botanikos sode. Optim. Ornam. Gard. Plant Assortment Technol. Environ. Sci. Artic. 2012, 3, 25–29. [Google Scholar]
- Fedulova, Y.; Kuklina, A.; Sorokopudov, V.; Sorokopudova, O.; Shlapakova, S.; Lukashov, Y. Screening of phytopathogens and phytopathoges on Chenomeles (Chaenomeles Lindl.) cultivars. In Proceedings of the BIO Web of Conferences, Online, 28 February 2020; EDP Sciences: Les Ulis, France, 2000; Volume 17, pp. 1–4. [Google Scholar] [CrossRef]
- Moral, J.; Muñoz-Díez, C.; Cabello, D.; Arquero, O.; Lovera, M.; Benítez, M.J.; Trapero, A. Characterization of Monilia disease caused by Monilinia linhartiana on quince in Southern Spain. Plant Pathol. 2011, 60, 1128–1139. [Google Scholar] [CrossRef]
- Hong, C.X.; Michailides, T.J.; Holtz, B.A. Mycoflora of stone fruit mummies in California orchards. Plant Dis. 2000, 84, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Thomma, B.P.H.J. Alternaria spp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef]
- Børve, J.; Røen, D.; Stensvand, A. Harvest time influences incidence of storage diseases and fruit quality in organically grown “Aroma” apples. Eur. J. Hortic. Sci. 2013, 78, 232–238. [Google Scholar]
- Weber, R.W.S.; Dralle, N.; Jork, O.; Niedersachsen, L. Fungi associated with blossom-end rot of apples in Germany. Eur. J. Hortic. Sci. 2013, 78, 97–105. [Google Scholar]
- Muñoz, M.; Faust, J.E.; Schnabel, G. Characterization of Botrytis cinerea from commercial cut flower roses. Plant Dis. 2019, 103, 1577–1583. [Google Scholar] [CrossRef]
- Tanovic, B.; Hrustic, J.; Mihajlovic, M.; Grahovac, M.; Delibasic, G. Botrytis cinerea in raspberry in Serbia I: Morphological and molecular characterization. Pestic. Fitomedicina 2014, 29, 237–247. [Google Scholar] [CrossRef]
- Grantina-Ievina, L. Fungi causing storage rot of apple fruit in Integrated Pest Management system and their sensitivity to fungicides. Rural Sustain. Res. 2015, 34, 2–11. [Google Scholar] [CrossRef]
- Rasiukevičiūtė, N.; Moročko-Bičevska, I.; Sasnauskas, A. Characterisation of growth variability and mycelial compatibility of Botrytis cinerea isolates originated from apple and strawberry in Lithuania. Proc. Latv. Acad. Sci. Sect. B Nat. Exact. Appl. Sci. 2017, 71, 217–224. [Google Scholar] [CrossRef]
- Rivera, Y.; Zeller, K.; Srivastava, S.; Sutherland, J.; Galvez, M.; Nakhla, M.; Poniatowska, A.; Schnabel, G.; Sundin, G.; Abad, Z.G. Draft genome resources for the phytopathogenic fungi Monilinia fructicola, M. fructigena, M. polystroma, and M. laxa, the causal agents of brown rot. Phytopathology 2018, 108, 1141–1142. [Google Scholar] [CrossRef]
- Šernaitė, L.; Rasiukevičiūtė, N.; Dambrauskienė, E.; Viškelis, P.; Valiuškaitė, A. Biocontrol of strawberry pathogen Botrytis cinerea using plant extracts and essential oils. Zemdirbyste 2020, 107, 147–152. [Google Scholar] [CrossRef]
- Tran, T.T.; Li, H.; Nguyen, D.Q.; Sivasithamparam, K.; Jones, M.G.K.; Wylie, S.J. Comparisons between genetic diversity, virulence and colony morphology of Monilinia fructicola and Monilinia laxa isolates. J. Plant Pathol. 2020, 102, 743–751. [Google Scholar] [CrossRef]
- Sholberg, P.L.; Harlton, C.; Haag, P.; Lévesque, C.A.; O’Gorman, D.; Seifert, K. Benzimidazole and diphenylamine sensitivity and identity of Penicillium spp. that cause postharvest blue mold of apples using β-tubulin gene sequences. Postharvest Biol. Technol. 2005, 36, 41–49. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, S.W.; Choi, K.H.; Kim, D.A.; Uhm, J.Y. Survey on the occurrence of apple diseases in Korea from 1992 to 2000. Plant Pathol. J. 2006, 22, 375–380. [Google Scholar] [CrossRef]
- Havenga, M.; Gatsi, G.M.; Halleen, F.; Spies, C.F.J.; van der Merwe, R.; Mostert, L. Canker and wood rot pathogens present in young apple trees and propagation material in the Western Cape of South Africa. Plant Dis. 2019, 103, 3129–3141. [Google Scholar] [CrossRef]
- Muñoz, G.; Hinrichsen, P.; Brygoo, Y.; Giraud, T. Genetic characterisation of Botrytis cinerea populations in Chile. Mycol. Res. 2002, 106, 594–601. [Google Scholar] [CrossRef]
- Mazzola, M. Identification and pathogenicity of Rhizoctonia spp. isolated from apple roots and orchard soils. Phytopathology 1997, 87, 582–587. [Google Scholar] [CrossRef]
- Tewoldemedhin, Y.T.; Mazzola, M.; Botha, W.J.; Spies, C.F.J.; McLeod, A. Characterization of fungi (Fusarium and Rhizoctonia) and oomycetes (Phytophthora and Pythium) associated with apple orchards in South Africa. Eur. J. Plant Pathol. 2011, 130, 215–229. [Google Scholar] [CrossRef]
- Kulshrestha, S.; Seth, C.A.; Sharma, M.; Sharma, A.; Mahajan, R.; Chauhan, A. Biology and control of Rosellinia necatrix causing white root rot disease: A Review. J. Pure Appl. Microbiol. 2014, 8, 1803–1814. [Google Scholar]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Gareth Jones, E.B.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Ten Hoopen, G.M.; Krauss, U. Biology and control of Rosellinia bunodes, Rosellinia necatrix and Rosellinia pepo: A Review. Crop Prot. 2006, 25, 89–107. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; et al. Diversity, morphology and molecular phylogeny of dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- De Carvalho, J.O.; Broll, V.; Martinelli, A.H.S.; Lopes, F.C. Endophytic fungi: Positive association with plants. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Al-Ani, L.K.T., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 321–332. [Google Scholar] [CrossRef]
- Hao, Y.; Aluthmuhandiram, J.V.S.; Chethana, K.W.T.; Manawasinghe, I.S.; Li, X.; Liu, M.; Hyde, K.D.; Phillips, A.J.L.; Zhang, W. Nigrospora species associated with various hosts from Shandong Peninsula, China. Mycobiology 2020, 48, 169–183. [Google Scholar] [CrossRef]
- Sun, Z.B.; Li, S.D.; Ren, Q.; Xu, J.L.; Lu, X.; Sun, M.H. Biology and applications of Clonostachys rosea. J. Appl. Microbiol. 2020, 129, 486–495. [Google Scholar] [CrossRef]
- Grantina-Ievina, L.; Stanke, L. Incidence and severity of leaf and fruit diseases of plums in Latvia. Commun. Agric. Appl. Biol. Sci. 2015, 80, 421–433. [Google Scholar]
- Castillo, H.; Rojas, R.; Villalta, M. Gliocladium sp., agente biocontrolador con aplicaciones prometedoras. Rev. Tecnol. Marcha 2016, 29, 65. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; El Hamss, H.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Sutton, J.C.; Li, D.W.; Peng, G.; Yu, H.; Zhang, P.; Valdebenito-Sanhueza, R.M. Gliocladium roseum: A versatile adversary of Botrytis cinerea in crops. Plant Dis. 1997, 81, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.M.; Zhang, X.; Zhang, N.N.; Naklumpa, W.; Zhao, W.Y.; Liang, X.F.; Zhang, R.; Sun, G.Y.; Gleason, M.L. Genera Acremonium and Sarocladium cause brown spot on bagged apple fruit in China. Plant Dis. 2019, 103, 1889–1901. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.H.; Kirschner, R. Sarocladium spinificis, a new endophytic species from the coastal grass Spinifex littoreus in Taiwan. Bot. Stud. 2014, 55, 25. [Google Scholar] [CrossRef]
- Liu, X. Sarocladium brachiariae Sp. Nov., an endophytic fungus isolated from Brachiaria brizantha. Mycosphere 2017, 8, 827–834. [Google Scholar] [CrossRef]
- Giraldo, A.; Gené, J.; Sutton, D.A.; Madrid, H.; de Hoog, G.S.; Cano, J.; Decock, C.; Crous, P.W.; Guarro, J. Phylogeny of Sarocladium (Hypocreales). Persoonia Mol. Phylogeny Evol. Fungi 2015, 34, 10–24. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.; Tudzynski, P.; Van Kan, J.A.L. Botrytis cinerea: The cause of grey mould disease. Molecular Plant Pathology. 2007, 8, 561–580. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhao, W.; Lin, R.; Yao, Y.; Yu, S.; Zhou, Z.; Zhang, X.; Gao, Y.; Huai, W. Fusarium species in declining wild apple forests on the Northern slope of the Tian Shan Mountains in North-Western China. For. Pathol. 2019, 49, e12542. [Google Scholar] [CrossRef]
- Racedo, J.; Salazar, S.M.; Castagnaro, A.P.; Díaz Ricci, J.C. A Strawberry disease caused by Acremonium strictum. Eur. J. Plant Pathol. 2013, 137, 649–654. [Google Scholar] [CrossRef]
- Papavasileiou, A.; Madesis, P.B.; Karaoglanidis, G.S. Identification and differentiation of Monilinia species causing brown rot of pome and stone fruit using High-resolution melting (HRM) analysis. Phytopathology 2016, 106, 1055–1064. [Google Scholar] [CrossRef]
- Madbouly, A.K.; Abo Elyousr, K.A.M.; Ismail, I.M. Biocontrol of Monilinia fructigena, causal agent of brown rot of apple fruit, by using endophytic yeasts. Biol. Control 2020, 144, 104239. [Google Scholar] [CrossRef]
- Hortova, B.; Novotny, D.; Erban, T. Physiological characteristics and pathogenicity of eight Neofabraea isolates from apples in Czechia. Eur. J. Hortic. Sci. 2014, 79, 327–334. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Köhn, H.-F.; Hubert, L.J. Hierarchical Cluster Analysis. In Wiley StatsRef: Statistics Reference Online; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2015; pp. 1–13. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakobija, I.; Bankina, B.; Klūga, A.; Roga, A.; Skinderskis, E.; Fridmanis, D. The Diversity of Fungi Involved in Damage to Japanese Quince. Plants 2022, 11, 2572. https://doi.org/10.3390/plants11192572
Jakobija I, Bankina B, Klūga A, Roga A, Skinderskis E, Fridmanis D. The Diversity of Fungi Involved in Damage to Japanese Quince. Plants. 2022; 11(19):2572. https://doi.org/10.3390/plants11192572
Chicago/Turabian StyleJakobija, Inta, Biruta Bankina, Alise Klūga, Ance Roga, Edmunds Skinderskis, and Dāvids Fridmanis. 2022. "The Diversity of Fungi Involved in Damage to Japanese Quince" Plants 11, no. 19: 2572. https://doi.org/10.3390/plants11192572