
Transcriptomics Using the Enriched Arabidopsis Shoot Apex Reveals Developmental Priming Genes Involved in Plastic Plant Growth under Salt Stress Conditions

Abstract
:1. Introduction
2. Results
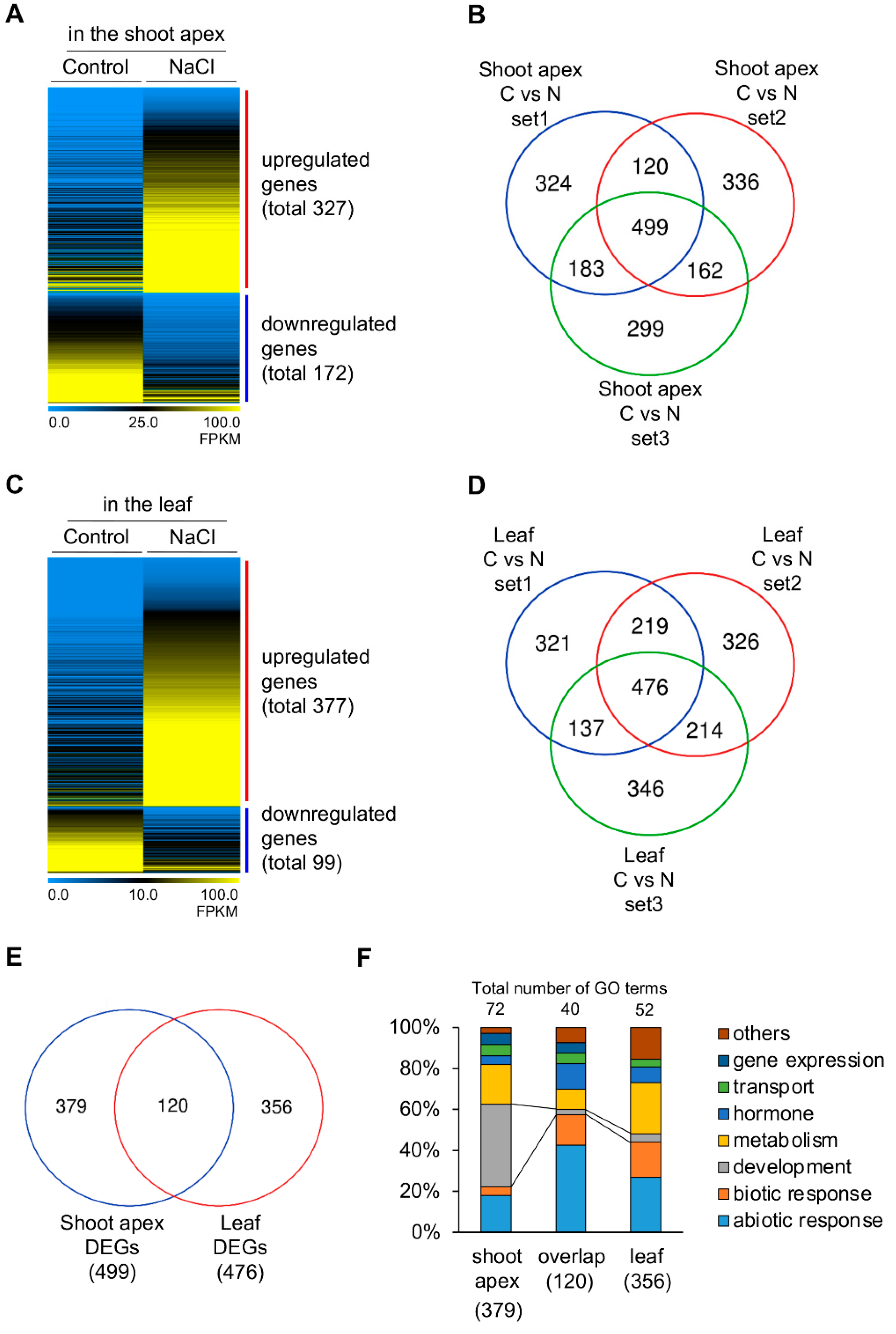
2.1. Developmental Plasticity Involved in the Shoot Growth and Development against Salt Stress Conditions
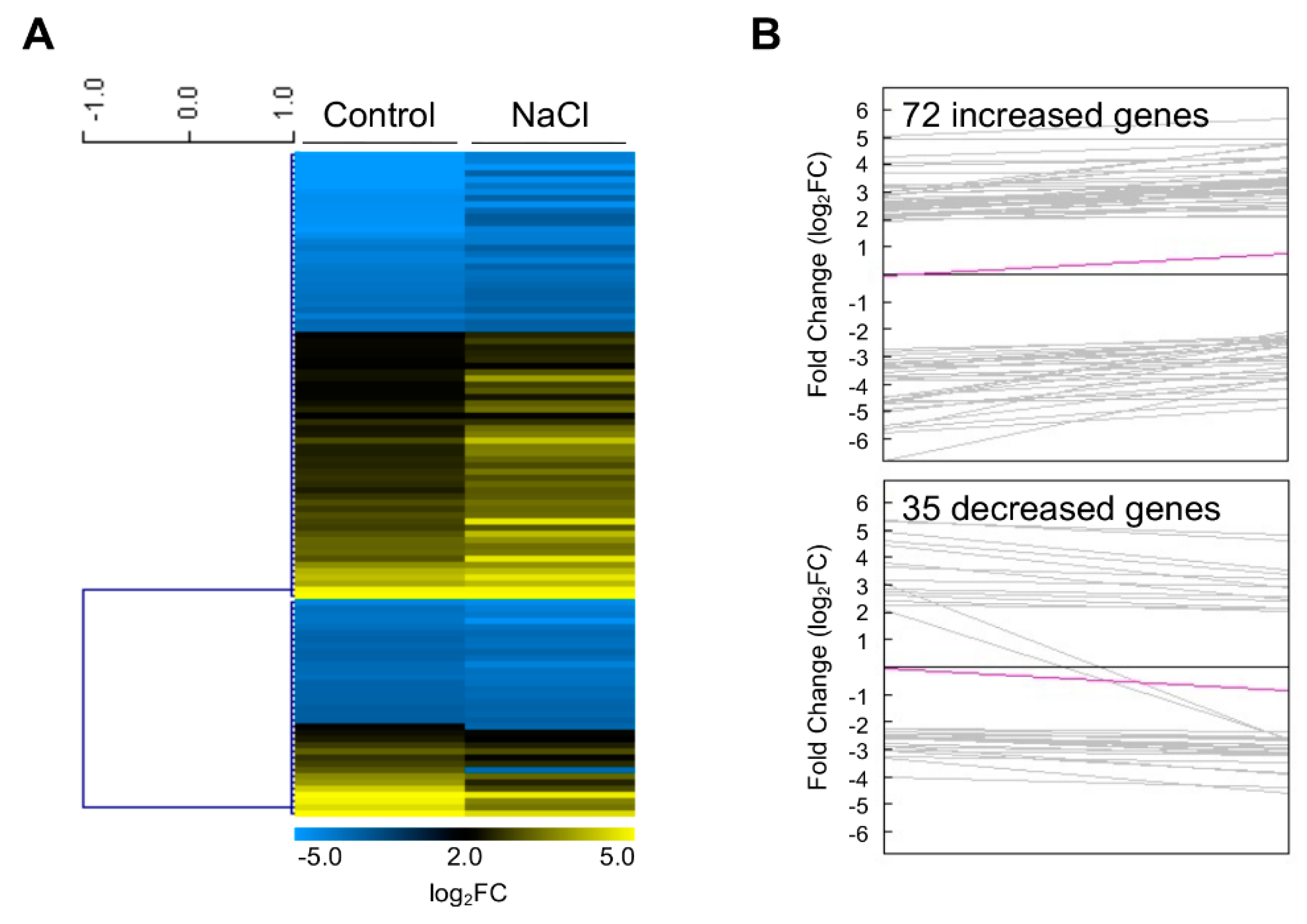
2.2. Identification of Developmental Priming Genes in the Shoot Apex
2.3. Clustering Analysis of Developmental Priming Genes in the Shoot Apex
2.4. Identification of Salt-Inducible Genes Only in the Shoot Apex
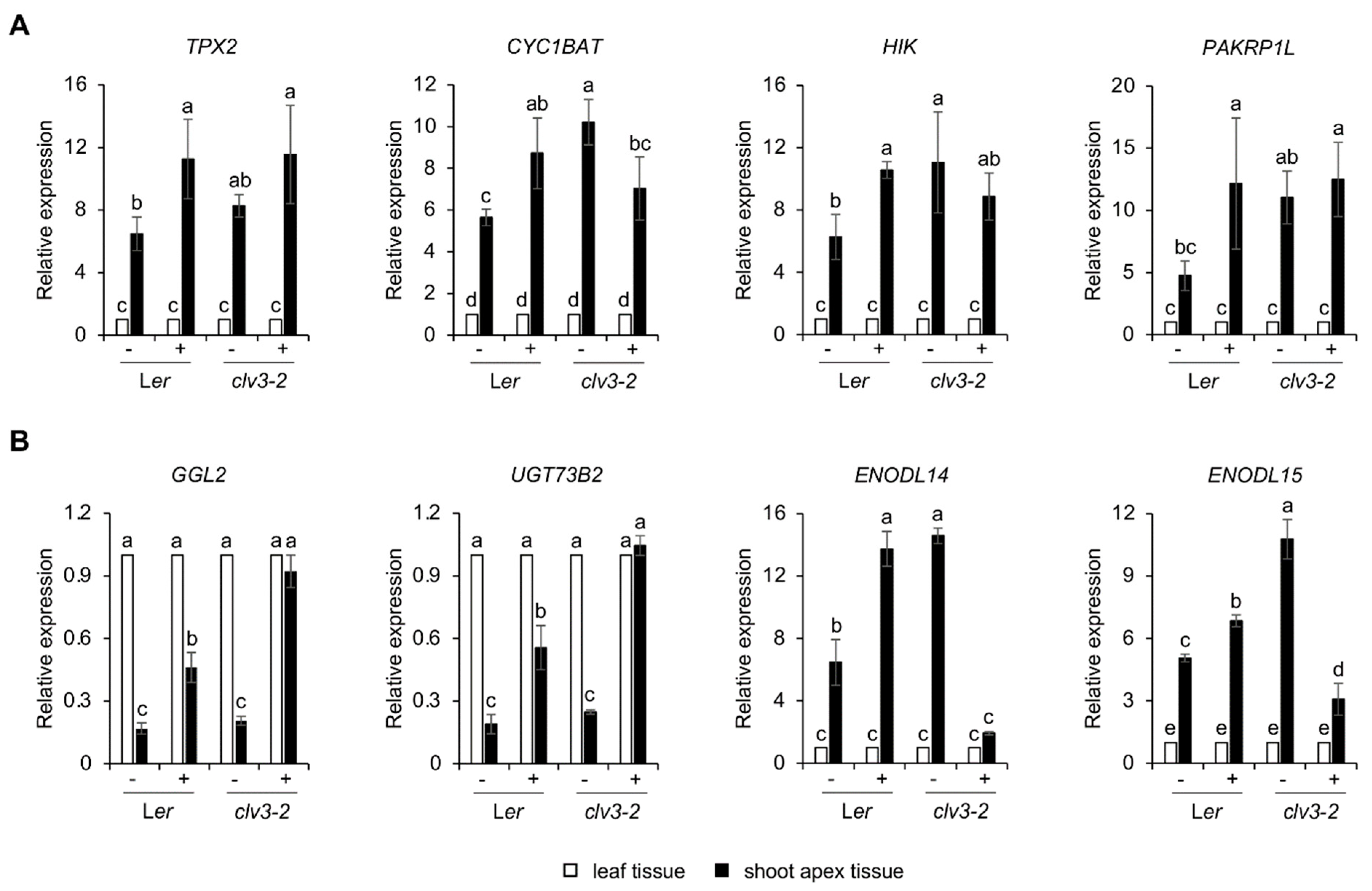
2.5. Identification of Developmental Priming Genes Related to Salt in the Shoot Apex
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. RNA Extraction and RNA Sequencing
4.3. Transcriptome Data Analysis
4.4. cDNA Synthesis and qRT-PCR
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lopes, F.L.; Galvan-Ampudia, C.; Landrein, B. WUSCHEL in the shoot apical meristem: Old player, new tricks. J. Exp. Bot. 2021, 72, 1527–1535. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jun, Y.S.; Cha, O.-K.; Sheen, J. Mitogen-activated protein kinases MPK3 and MPK6 are required for stem cell maintenance in the Arabidopsis shoot apical meristem. Plant Cell Rep. 2019, 38, 311–319. [Google Scholar] [CrossRef] [PubMed]
- De Jong, M.; Leyser, O. Developmental plasticity in plants. Cold Spring Harb. Sym. Quant. Biol. 2012, 77, 63–73. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. The link between flowering time and stress tolerance. J. Exp. Bot. 2016, 67, 47–60. [Google Scholar] [CrossRef]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Wang, Z.; Kim, W.S. Effect of drought stress on shoot growth and physiological response in the cut rose ‘charming black’ at different developmental stages. Hortic. Environ. Biotechnol. 2019, 60, 1–8. [Google Scholar] [CrossRef]
- Trontin, C.; Kiani, S.; Corwin, J.A.; Hématy, K.; Yansouni, J.; Kliebenstein, K.J.; Loudet, O. A pair of receptor-like kinases is responsible for natural variation in shoot growth response to mannitol treatment in Arabidopsis thaliana. Plant J. 2014, 78, 121–133. [Google Scholar] [CrossRef]
- Wilson, M.E.; Mixdorf, M.; Berg, R.H.; Haswell, E.S. Plastid osmotic stress influences cell differentiation at the plant shoot apex. Development 2016, 143, 3382–3393. [Google Scholar]
- Claeys, H.; Van Landeghem, S.; Dubois, M.; Maleux, K.; Inzé, D. What Is Stress? Dose-Response Effects in Commonly Used in Vitro Stress Assays. Plant Physiol. 2014, 165, 519–527. [Google Scholar] [CrossRef]
- Crisp, P.A.; Ganguly, D.; Eichten, S.R.; Borevitz, J.O.; Pogson, B.J. Reconsidering plant memory: Intersections between stress recovery, RNA turnover, and epigenetics. Sci. Adv. 2016, 2, e1501340. [Google Scholar] [CrossRef]
- Khan, A.; Khan, V.; Pandey, K.; Sopory, S.K.; Sanan-Mishra, N. Thermo-Priming Mediated Cellular Networks for Abiotic Stress Management in Plants. Front. Plant Sci. 2022, 13, 866409. [Google Scholar] [CrossRef] [PubMed]
- Grafi, G.; Chalifa-Caspi, V.; Nagar, T.; Plaschkes, I.; Barak, S.; Ransbotyn, V. Plant response to stress meets dedifferentiation. Planta 2011, 233, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Weits, D.A.; Kunkowska, A.B.; Kapms, N.C.W.; Portz, K.M.S.; Packbier, N.K.; Venza, Z.N.; Gaillochet, C.; Lohmann, J.U.; Pedersen, O.; Van Dongen, J.T.; et al. An apical hypoxic niche sets the pace of shoot meristem activity. Nature 2019, 569, 714–717. [Google Scholar] [CrossRef] [PubMed]
- Olas, J.J.; Apelt, F.; Annunziata, M.G.; John, S.; Richard, S.I.; Gupta, S.; Kragler, F.; Balazadeh, S.; Mueller-Roeber, B. Primary carbohydrate metabolism genes participate in heat-stress memory at the shoot apical meristem of Arabidopsis thaliana. Mol. Plant 2021, 14, 1508–1524. [Google Scholar] [CrossRef]
- Jun, Y.S.; Cha, O.-K.; Kim, J.H.; Lee, H. Shoot Meristem Activity is Involved in Salt Tolerance on Arabidopsis Shoot Growth. J. Plant Biol. 2019, 62, 410–418. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Klei, K.; Fokkens, L.; Haring, M.A.; Schranz, M.E.; Testerink, C. Natural variation in rosette size under salt stress conditions corresponds to developmental differences between Arabidopsis accessions and allelic variation in the LRR-KISS gene. J. Exp. Bot. 2016, 67, 2127–2138. [Google Scholar] [CrossRef]
- Yadav, R.K.; Tavakkoli, M.; Xie, M.; Girke, T.; Reddy, G.V. A high-resolution gene expression map of the Arabidopsis shoot meristem stem cell niche. Development 2014, 141, 2735–2744. [Google Scholar] [CrossRef]
- Ribone, P.A.; Capella, M.; Chan, R.L. Functional characterization of the homeodomain leucine zipper I transcription factor AtHB13 reveals a crucial role in Arabidopsis development. J. Exp. Bot. 2015, 66, 5929–5943. [Google Scholar] [CrossRef]
- Cabello, J.V.; Chan, R.L. The homologous homeodomain-leucine zipper transcription factors HaHB1 and AtHB13 confer tolerance to drought and salinity stresses via the induction of proteins that stabilize membrane. Plant Biotechnol. J. 2012, 10, 815–825. [Google Scholar] [CrossRef]
- Gao, D.; Appiano, M.; Huibers, R.P.; Chen, X.; Loonen, A.E.H.M.; Visser, R.G.F.; Wolters, A.-M.A.; Bai, Y. Activation tagging of ATHB13 in Arabidopsis thaliana confers broad-spectrum disease resistance. Plant Mol. Biol. 2014, 86, 641–653. [Google Scholar] [CrossRef]
- Chung, Y.; Zhu, Y.; Wu, M.F.; Simonini, S.; Kuhn, A.; Armenta-Medina, A.; Jin, R.; Ostergaard, L.; Gillmor, C.S.; Wagner, D. Auxin Response Factors promote organogenesis by chromatin-mediated repression of the pluripotency gene SHOOTMERISTEMLESS. Nat. Commun. 2019, 10, 886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Choi, Y.-R.; Seo, P.J. Increased STM expression is associated with drought tolerance in Arabidopsis. J. Plant Physiol. 2016, 201, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Landrein, B.; Kiss, A.; Sassi, M.; Chauvet, A.; Das, P.; Cortizo, M.; Laufs, P.; Takeda, S.; Aida, M.; Traas, J.; et al. Mechanical stress contributes to the expression of the STM homeobox gene in Arabidopsis shoot meristem. eLife 2015, 4, e07811. [Google Scholar] [CrossRef] [PubMed]
- Hake, A.; Smith, H.M.S.; Holtan, H.; Magnani, E.; Mele, G.; Ramirez, J. The role of knox genes in plant development. Annu. Rev. Cell Dev. Biol. 2004, 20, 125–151. [Google Scholar] [CrossRef]
- Wang, L.; Hua, D.; He, J.; Duan, Y.; Chen, Z.; Hong, X.; Gong, Z. Auxin Response Factor2 (ARF3) and Its Regulated Homeodomain Gene HB33 Mediate Abscisic Acid Response in Arabidopsis. PLoS Genet. 2011, 7, e1002172. [Google Scholar] [CrossRef]
- Hong, S.-Y.; Kim, O.-K.; Kim, S.-G.; Yand, M.-S.; Park, C.-M. Nuclear Import and DNA Binding of the ZHD5 Transcription Factor Is Modulated by a Competitive Peptide Inhibitor in Arabidopsis. J. Biol. Chem. 2011, 286, 1659–1668. [Google Scholar] [CrossRef]
- Kim, J.-Y.; Ryu, J.Y.; Baek, K.; Park, C.-M. High temperature attenuates the gravitropism of inflorescence stems by inducing SHOOT GRAVITROPISM 5 alternative splicing in Arabidopsis. New Phytol. 2016, 209, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Lashbrook, C.C. Stamen Abscission Zone Transcriptome Profiling Reveals New Candidates for Abscission Control: Enhanced Retention of Floral Organs in Transgenic Plants Overexpressing Arabidopsis ZINC FINGER PROTEIN2. Plant Physiol. 2008, 146, 1305–1321. [Google Scholar] [CrossRef]
- Gubert, C.M.; Christy, M.E.; Ward, D.L.; Groner, W.D.; Liljegren, S.J. ASYMMETRIC LEAVES1 regulates abscission zone placement in Arabidopsis flowers. BMC Plant Biol. 2014, 14, 195. [Google Scholar] [CrossRef]
- Nurmberg, P.L.; Knox, K.A.; Yun, B.-W.; Morris, P.C.; Shafiei, R.; Hudson, A.; Loake, G.J. The developmental selector AS1 is an evolutionarily conserved regulator of the plant immune response. Proc. Natl. Acad. Sci. USA 2007, 104, 18795–18800. [Google Scholar] [CrossRef]
- Lee, D.-K.; Geisler, M.; Springer, P.S. LATERAL ORGAN FUSION1 and LATERAL ORGAN FUSION2 function in lateral organ separation and axillary meristem formation in Arabidopsis. Development 2009, 136, 2423–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randall, R.S.; Miyashima, S.; Blomster, T.; Zhang, J.; Elo, A.; Karlberg, A.; Immanen, J.; Nieminen, K.; Lee, J.-Y.; Kakimoto, T.; et al. AINTEGUMENTA and the D-type cyclin CYCD3;1 regulate root secondary growth and respond to cytokinins. Biol. Open 2015, 4, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Krizek, B.A.; Bequette, C.J.; Xu, K.; Blakley, I.C.; Fu, Z.Q.; Stratmann, J.W.; Loraine, A.E. RNA-Seq Links the Transcription Factors AINTEGUMENTA and AINTEGUMENTA-LIKE6 to Cell Wall Remodeling and Plant Defense Pathways. Plant Physiol. 2016, 171, 2069–2084. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Feng, C.-Z.; Ye, Q.; Yu, W.-H.; Chen, Y.-F. Arabidopsis WRKY6 Transcription Factor Acts as a Positive Regulator of Abscisic Acid Signaling during Seed Germination and Early Seedling Development. PLoS Genet. 2016, 12, e1005833. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-F.; Li, L.-Q.; Xu, Q.; Kong, Y.-H.; Wang, H.; Wu, W.-H. The WRKY6 Transcription Factor Modulate PHOSPHATE1 Expression in Response to Low Pi Stress in Arabidopsis. Plant Cell 2009, 21, 3554–3566. [Google Scholar] [CrossRef]
- Bernhardt, C.; Zhao, M.; Gonzalez, A.; Lloyd, A.; Schiefelbein, J. The bHLH genes GL3 and EGL3 participate in an intercellular regulatory circuit that controls cell patterning in the Arabidopsis root epidermis. Development 2005, 132, 291–298. [Google Scholar] [CrossRef]
- Long, J.A.; Moan, E.I.; Medford, J.I.; Barton, M.K. A member of the KNOTTED class of homeodomain proteins encoded by the STM gene of Arabidopsis. Nature 1996, 379, 66–69. [Google Scholar] [CrossRef]
- Magnani, E.; Hake, S. KNOX Lost the OX: The Arabidopsis KNATM Gene Defines a Novel Class of KNOX Transcriptional Regulators Missing the Homeodomain. Plant Cell 2008, 20, 875–887. [Google Scholar] [CrossRef]
- Solís-Guzmán, M.G.; Argüello-Astorga, G.; López-Bucio, J.; Ruiz-Herrera, L.F.; López-Meza, J.E.; Sánchez-Calderón, L.; Carreón-Abud, Y.; Martínez-Trujillo, M. Arabidopsis thaliana sucrose phosphate synthase (sps) genes are expressed differentially in organs and tissues, and their transcription is regulated by osmotic stress. Gene Expr. Patterns 2017, 25–26, 92–101. [Google Scholar] [CrossRef]
- Raggi, S.; Ferrarini, A.; Delledonne, M.; Dunand, C.; Ranocha, P.; de Lorenzo, G.; Cervone, F.; Ferrari, S. The Arabidopsis Class III Peroxidase AtPRX71 Negatively Regulates Growth under Physiological Conditions and in Response to Cell Wall Damage. Plant Physiol. 2015, 169, 2513–2525. [Google Scholar] [CrossRef]
- Nikonorova, N.; Van den Broeck, L.; Zhu, S.; van de Cotte, B.; Dubois, M.; Gevaert, K.; Inzé, D.; De Smet, I. Early mannitol-triggered changes in the Arabidopsis leaf (phospho)proteome reveal growth regulators. J. Exp. Bot. 2018, 69, 4591–4607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, H.; Wang, Y.; Cloix, C.; Li, K.; Jenkins, G.I.; Wang, S.; Shang, Z.; Shi, Y.; Yang, S.; Li, X. The Arabidopsis RCC1 Family Protein TCF1 Regulates Freezing Tolerance and Cold Acclimation through Modulating Lignin Biosynthesis. PLoS Genet. 2015, 11, e1005471. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Meier, K.; Dörr, F.; Ravindran, B.M. NSP-Dependent Simple Nitrile Formation Dominates upon Breakdown of Major Aliphatic Glucosinolates in Roots, Seeds, and Seedlings of Arabidopsis thaliana Columbia-0. Front. Plant Sci. 2016, 7, 1821. [Google Scholar] [CrossRef] [PubMed]
- La Camera, S.; Balague, C.; Cobel, C.; Ceoffroy, P.; Legrand, M.; Feussner, I.; Roby, D.; Heitz, T. The Arabidopsis patatin-like protein s (PLP2) plays an essential role in cell death execution and differentially affects biosynthesis of oxylipins and resistance to pathogens. Mol. Plant Microbe Interact. 2009, 22, 469–481. [Google Scholar] [CrossRef]
- Kuhn, H.; Lorek, J.; Kwaaitaal, M.; Consonni, C.; Becker, K.; Micali, C.; van Themaat, E.V.L.; Bednarek, P.; Raaymakers, T.M.; Appiano, M.; et al. Key Components of Different Plant Defense Pathways Are Dispensable for Powdery Mildew Resistance of the Arabidopsis mlo2 mol6 mol12 Triple Mutant. Front. Plant Sci. 2017, 8, 1006. [Google Scholar] [CrossRef]
- Shi, H.; Ye, T.; Chen, F.; Cheng, Z.; Wang, Y.; Yang, P.; Zhang, Y.; Chan, Z. Manipulation of arginase expression modulates abiotic stress tolerance in Arabidopsis: Effect on arginine metabolism and ROS accumulation. J. Exp. Bot. 2013, 64, 1367–1379. [Google Scholar] [CrossRef]
- Casanova-Sáez, R.; Mateo-Bonmatí, E.; Šimura, J.; Pěnčík, A.; Novák, O.; Staswick, P.; Ljung, K. Inactivation of the entire Arabidopsis group II GH3s confers tolerance to salinity and water deficit. New Phytol. 2022, 235, 263–275. [Google Scholar] [CrossRef]
- Pereira, A.M.; Masiero, S.; Sofia, M.; Nobre, S.; Costa, M.L.; Solís, M.-T.; Testillano, P.S.; Sprunck, S.; Coimbra, S. Differential expression patterns of arabinogalactan proteins in Arabidopsis thaliana reproductive tissues. J. Exp. Bot. 2014, 65, 5459–5471. [Google Scholar] [CrossRef]
- Reis, R.S.; Deforges, J.; Sokoloff, T.; Poirier, Y. Modulation of Shoot Phosphate Level and Growth by PHOSPHATE1 Upstream Open Reading Frame. Plant Physiol. 2020, 183, 1145–1156. [Google Scholar] [CrossRef]
- Rani, S.H.; Krishna, T.H.; Saha, S.; Negi, A.S.; Rajasekharan, R. Defective in cuticular ridges (DCR) of Arabidopsis thaliana, a gene associated with surface cutin formation, encodes a soluble diacylglycerol acyltransferase. J. Biol. Chem. 2010, 285, 38337–38347. [Google Scholar] [CrossRef]
- Hirner, A.; Ladwig, F.; Stransky, H.; Okumoto, S.; Keinath, M.; Harms, A.; Frommer, W.B.; Koch, W. Arabidopsis LHT1 Is a High-Affinity Transporter for Cellular Amino Acid Uptake in Both Root Epidermis and Leaf Mesophyll. Plant Cell 2006, 18, 1931–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Tsuda, K.; Nguyen, L.V.; Wang, X.; Lin, J.; Murphy, A.S.; Glazebrook, J.; Thordal-Christensen, H.; Katagiri, F. Physical association of Arabidopsis hypersensitive induced reaction proteins (HIRs) with the immune receptor RPS2. J. Biol. Chem. 2011, 286, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Hoang, X.L.T.; Nhi, D.N.H.; Thu, N.B.A.; Thao, N.P.; Tran, L.S.P. Transcription factors and their roles in signal transduction in plants under abiotic stresses. Curr. Genom. 2017, 18, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Geng, Y.; Wu, R.; Wee, C.W.; Xie, F.; Wei, X.; Chan, P.M.Y.; Tham, C.; Duan, L.; Dinneny, J.R. A Spatio-Temporal Understanding of Growth Regulation during the Salt Stress Response in Arabidopsis. Plant Cell 2013, 25, 2132–2154. [Google Scholar] [CrossRef]
- Endler, A.; Kesten, C.; Froehlich, A.; Zhang, Y.; Ivakov, A.; Froehlich, A.; Funke, N.; Persson, S. A mechanism for sustained cellulose synthesis during salt stress. Cell 2015, 162, 1353–1364. [Google Scholar] [CrossRef]
- Bird, A.W.; Hyman, A.A. Building a spindle of the correct length in human cells requires the interaction between TPX2 and Aurora A. J. Cell Biol. 2008, 182, 289–300. [Google Scholar] [CrossRef]
- Izawa, D.; Pines, J. The mitotic checkpoint complex binds a second CDC20 to inhibit active APC/C. Nature 2015, 517, 631–634. [Google Scholar] [CrossRef]
- Petrovská, B.; Cenklová, V.; Pochylová, Z.; Kourová, H.; Doskočilová, A.; Plíhal, O.; Binarová, L.; Binarová, P. Plant Aurora kinases play a role in maintenance of primary meristems and control of endoreduplication. New Phytol. 2012, 193, 590–604. [Google Scholar] [CrossRef]
- Niu, B.; Wang, L.; Zhang, L.; Ren, D.; Ren, R.; Copenhaver, G.P.; Ma, H.; Wang, Y. Arabidopsis Cell Division Cycle 20.1 Is Required for Normal Meiotic Spindle Assembly and Chromosome Segregation. Plant Cell 2015, 27, 3367–3382. [Google Scholar] [CrossRef]
- Blethrow, J.D.; Glavy, J.S.; Morgan, D.O.; Shokat, K.M. Covalent capture of kinase-specific phosphopeptides reveals Cdk1-cyclin B substrates. Proc. Natl. Acad. Sci. USA 2008, 105, 1442–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitiss, J.L. Investigating the biological functions of DNA topoisomerases in eukaryotic cell. Biochim. Biophys. Acta 1998, 1400, 63–81. [Google Scholar] [CrossRef]
- Parra-Nunez, P.; Cooper, C.; Sanchez-Moran, E. The Role of DNA Topoisomerase Binding Protein 1 (TopBP1) in Genome Stability in Arabidopsis. Plants 2021, 10, 2568. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequencing from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef]
- Sun, J.; Nishiyama, T.; Shimizu, K.; Kadota, K. TCC: An R package for comparing tag count data with robust normalization strategies. BMC Bioinform. 2013, 14, 219. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AGI Number | Synonym | Type of TF | Developmental Function | Stress Function | References |
|---|---|---|---|---|---|
| AT3G30530 | ATbZIP42 | basic leucine zipper | - | - | |
| AT1G69780 | ATHB13 | HD-ZIP family | reproductive organ development | low temperature/drought/salt/disease | [18,19,20] |
| AT1G62360 | STM | KNOX/ELK | SAM initiation/maintenance via auxin | mechanical/drought stress response | [21,22,23,24] |
| AT1G75240 | ATHB33 | ZF-HD family | root and whole plant growth | ABA response | [25,26] |
| AT2G01940 | SGR5 | zinc finger protein | shoot gravitropism | high temperature response | [27] |
| AT5G57520 | ZFP2 | zinc finger protein | abscission of floral organ | - | [28] |
| AT2G37630 | AS1 | MYB protein | abscission of floral organ | defense response | [29,30] |
| AT1G26780 | MYB117 | MYB protein | axillary meristem formation | - | [31] |
| AT4G37750 | ANT | AP2 family protein | cell proliferation in root growth | defense response | [32,33] |
| AT2G34830 | WRKY35 | WRKY protein | - | - | |
| AT1G62300 | WRKY6 | WRKY protein | ABA signaling in seedling development | low Pi stress response | [34,35] |
| AT1G63650 | EGL3 | bHLH family | cell patterning in root epidermis | - | [36] |
| AT1G49830 | - | bHLH family | - | - |
| AGI Number | Synonym | Control | NaCl | Function | References | ||||
|---|---|---|---|---|---|---|---|---|---|
| Avg. log2FC | SD | Primed State | Avg. log2FC | SD | Primed State | ||||
| increased DEGs | |||||||||
| AT4G10120 | ATSPS4F | −5.67 | 0.36 | DOWN | −2.50 | 0.33 | DOWN | osmotic stress | [39] |
| AT5G64120 | PER71 | −6.79 | 0.79 | DOWN | −3.76 | 0.78 | DOWN | negative effects of growth via ROS | [40] |
| AT3G48310 | CYP71A22 | −4.64 | 0.22 | DOWN | −2.10 | 0.36 | DOWN | - | |
| AT3G22060 | CRRSP38 | −4.71 | 0.12 | DOWN | −2.26 | 0.47 | DOWN | regulation of leaf growth | [41] |
| AT5G20230 | ATBCB | −4.47 | 0.56 | DOWN | −2.48 | 0.54 | DOWN | cold acclimation in lignin biosynthesis | [42] |
| AT3G16400 | NSP1 | 2.84 | 0.12 | UP | 4.76 | 0.79 | UP | defense metabolite formation | [43] |
| AT3G16410 | NSP4 | 2.99 | 0.40 | UP | 4.73 | 0.41 | UP | defense metabolite formation | [43] |
| AT2G26560 | PLP2 | −5.49 | 1.15 | DOWN | −3.77 | 1.41 | DOWN | cell death and biotic stress response | [44] |
| AT2G39200 | MLO12 | −5.05 | 0.54 | DOWN | −3.35 | 0.72 | DOWN | defense response | [45] |
| AT4G08870 | ARGAH2 | −4.60 | 0.26 | DOWN | −2.90 | 1.14 | DOWN | abiotic stress tolerance | [46] |
| decreased DEGs | |||||||||
| AT2G23170 | GH3.3 | 3.06 | 0.56 | UP | −2.67 | 0.96 | DOWN | IAA-dependent salt tolerance | [47] |
| AT5G64310 | AGP1 | 2.08 | 0.06 | UP | −2.64 | 0.37 | DOWN | reproductive process | [48] |
| AT3G23430 | PHO1 | 4.44 | 0.38 | UP | 2.89 | 0.12 | UP | shoot phosphate level and growth | [49] |
| AT5G23940 | EMB3009 | 4.90 | 0.65 | UP | 3.55 | 0.53 | UP | cutin Biosynthesis | [50] |
| AT4G27260 | GH3.5 | 3.81 | 0.21 | UP | 2.47 | 0.29 | UP | IAA-dependent salt tolerance | [47] |
| AT5G40780 | LHT1 | −3.33 | 0.32 | DOWN | −4.62 | 0.41 | DOWN | uptake of amino acid | [51] |
| AT1G75780 | TUB1 | 4.61 | 0.48 | UP | 3.36 | 0.44 | UP | - | |
| AT1G62300 | WRKY6 | −2.89 | 0.16 | DOWN | −3.91 | 0.49 | DOWN | low-Pi stress response | [35] |
| AT3G01290 | HIR2 | −3.07 | 0.48 | DOWN | −3.83 | 0.55 | DOWN | hypersensitive response with immunity | [52] |
| AT2G28790 | - | 5.33 | 0.33 | UP | 4.61 | 0.27 | UP | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cha, O.-K.; Yang, S.; Lee, H. Transcriptomics Using the Enriched Arabidopsis Shoot Apex Reveals Developmental Priming Genes Involved in Plastic Plant Growth under Salt Stress Conditions. Plants 2022, 11, 2546. https://doi.org/10.3390/plants11192546
Cha O-K, Yang S, Lee H. Transcriptomics Using the Enriched Arabidopsis Shoot Apex Reveals Developmental Priming Genes Involved in Plastic Plant Growth under Salt Stress Conditions. Plants. 2022; 11(19):2546. https://doi.org/10.3390/plants11192546
Chicago/Turabian StyleCha, Ok-Kyoung, Soeun Yang, and Horim Lee. 2022. "Transcriptomics Using the Enriched Arabidopsis Shoot Apex Reveals Developmental Priming Genes Involved in Plastic Plant Growth under Salt Stress Conditions" Plants 11, no. 19: 2546. https://doi.org/10.3390/plants11192546