

Physiological and Transcriptomic Analyses Reveal the Mechanisms of Compensatory Growth Ability for Early Rice after Low Temperature and Weak Light Stress
 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Experimental Design
2.3. Agronomic Trait Determination
2.4. Determination of Soluble Protein, H2O2 Concentration, and Antioxidant Enzyme Activity
2.5. Determination of NR, GS, and Rubisco Activities
2.6. Determination of Endogenous Hormone Concentrations
2.7. RNA Extraction and Transcriptome Sequencing
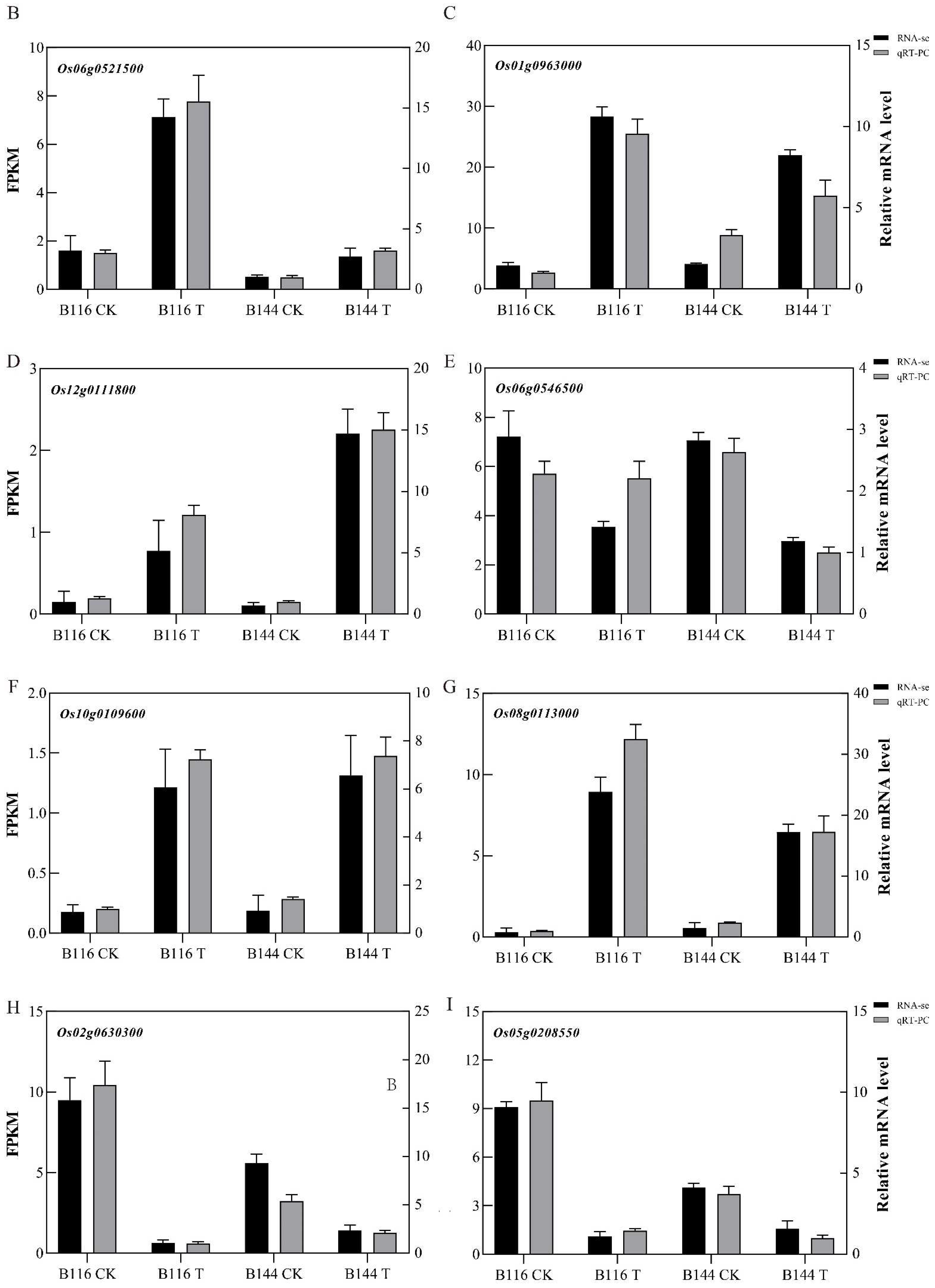
2.8. Quantitative RT-PCR Validations
2.9. Statistical Analysis
3. Results
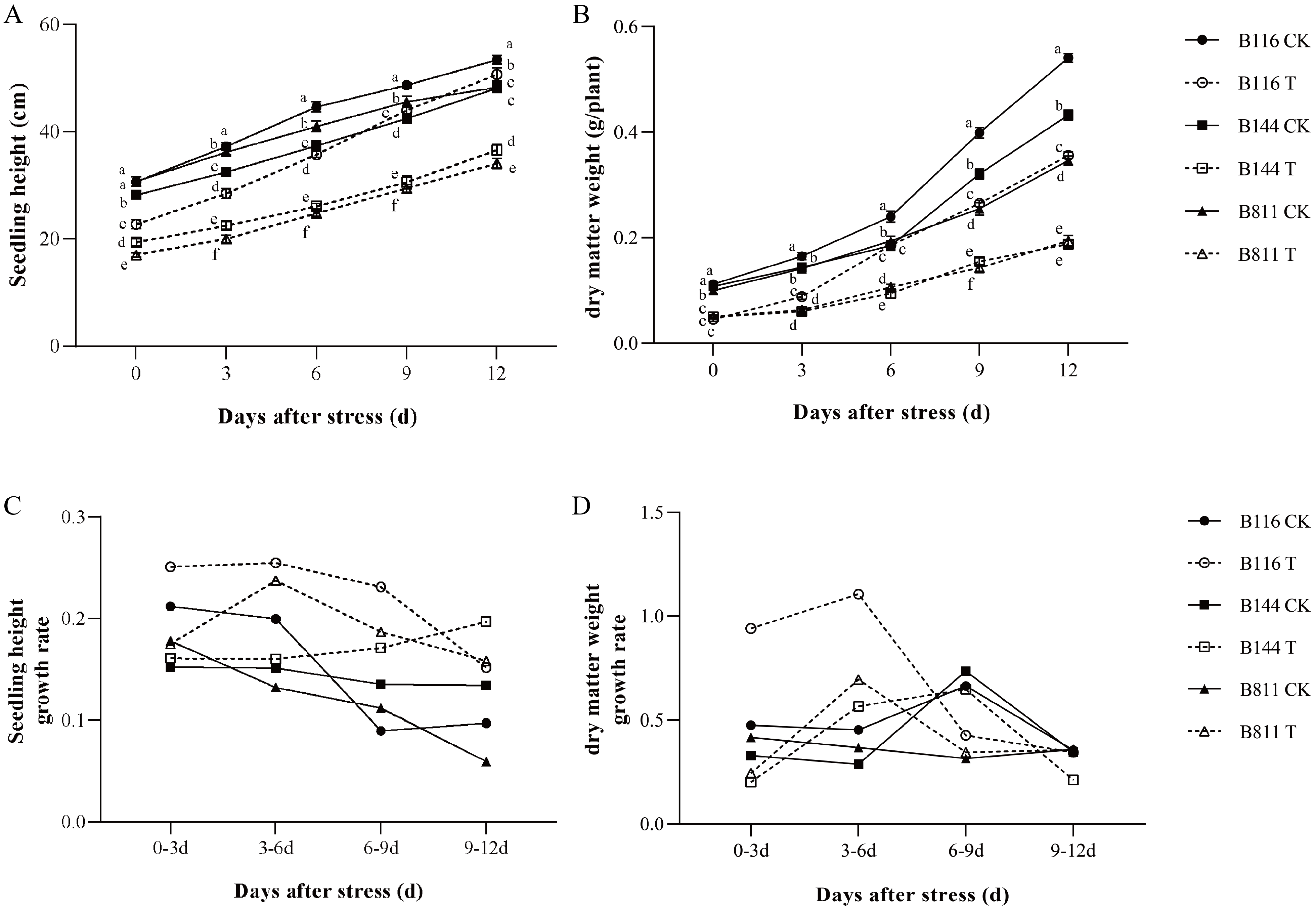
3.1. Differences in Agronomic Traits
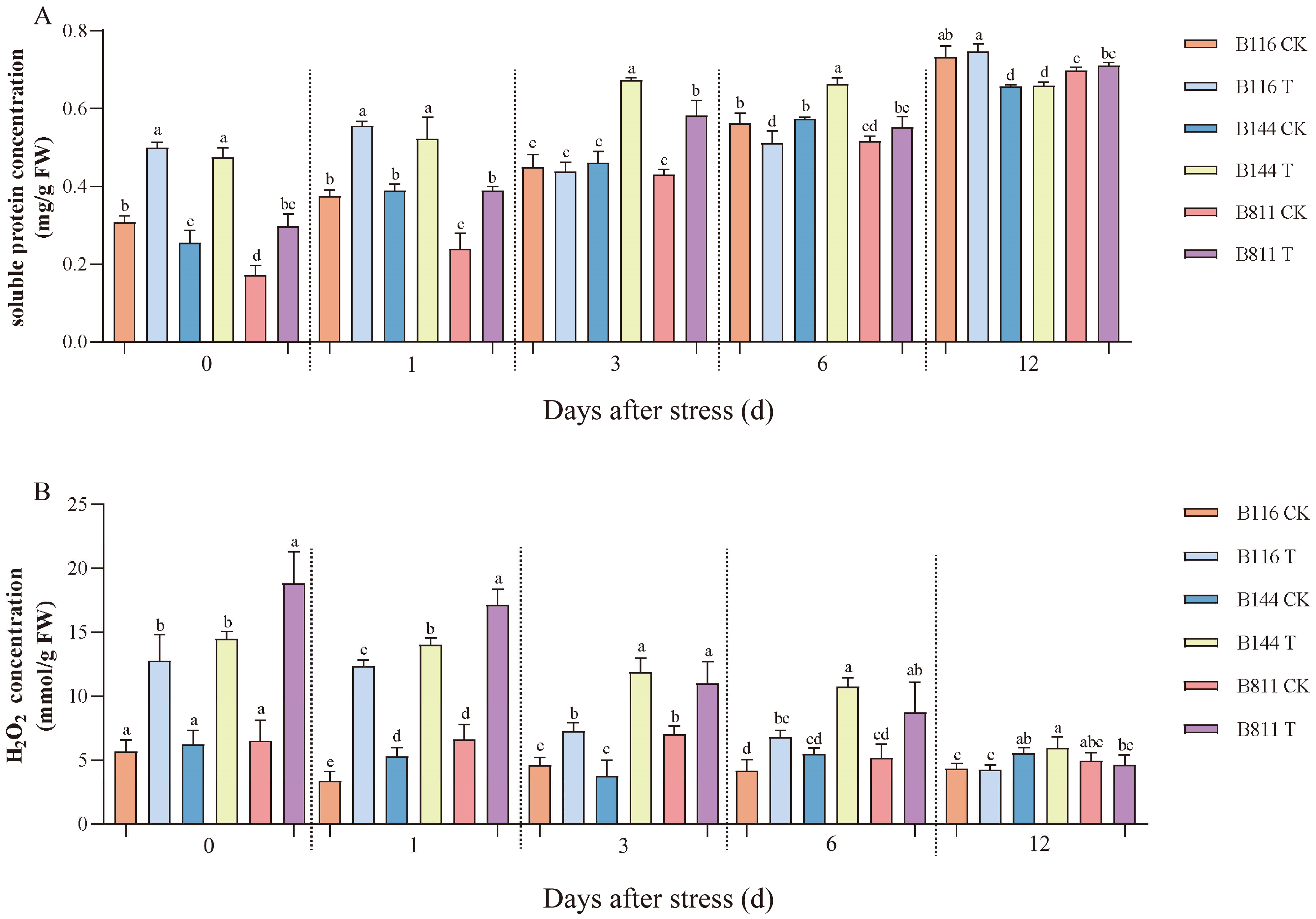
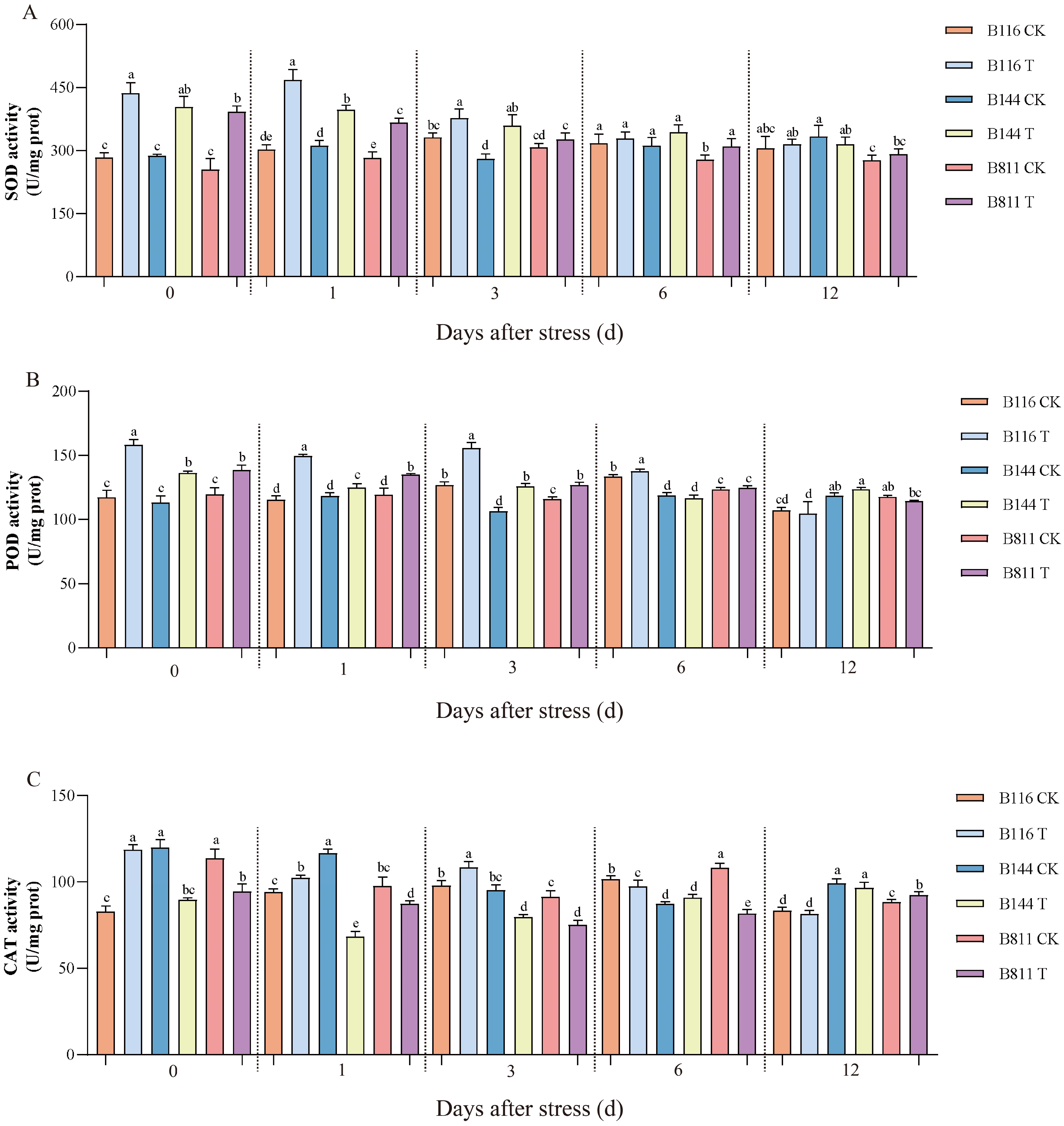
3.2. Differences in Soluble Protein, H2O2 Concentration, and Antioxidant Enzyme Activity
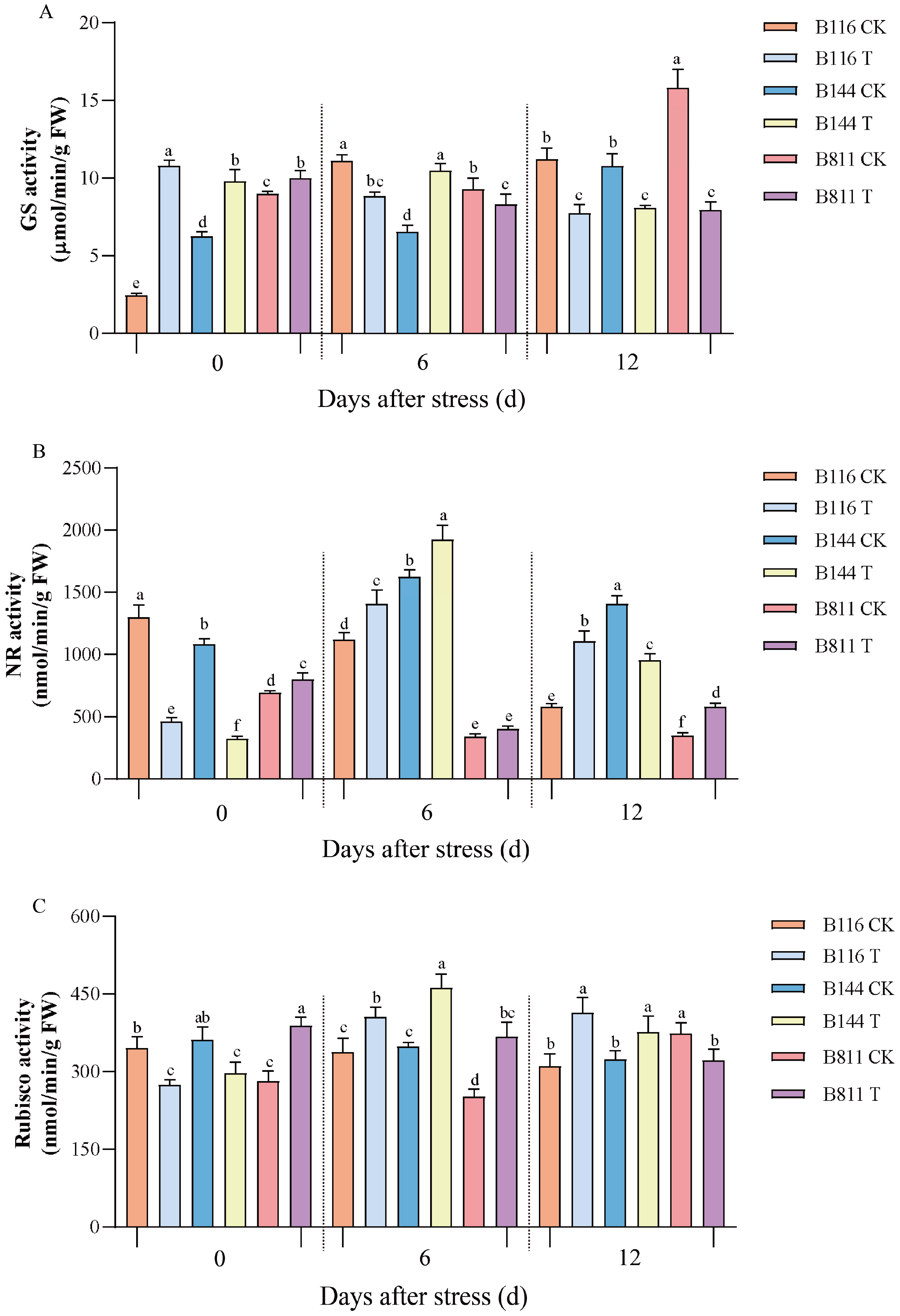
3.3. Differences in NR, GS, and Rubisco Activity
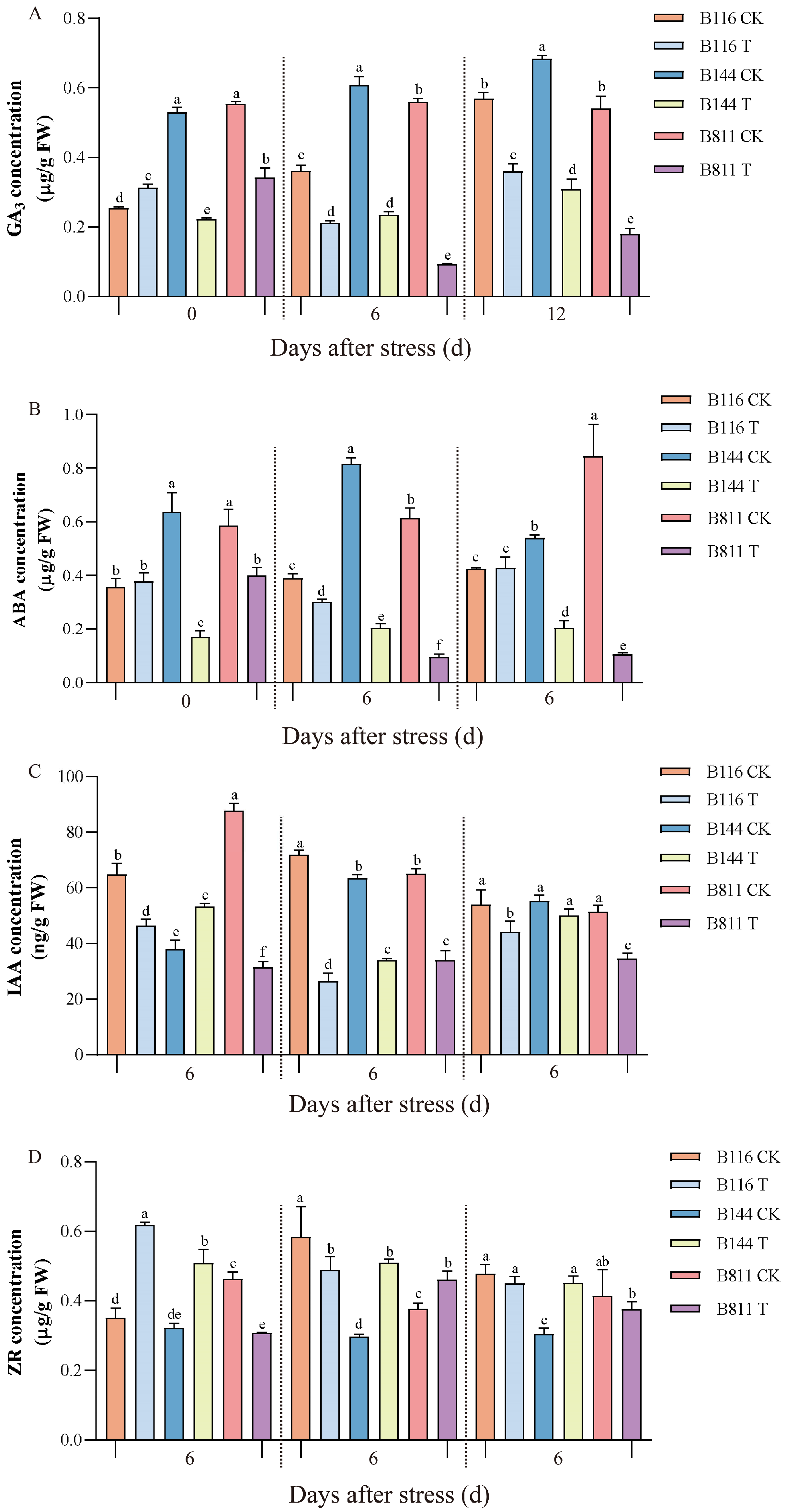
3.4. Differences in GA3, ABA, IAA, and ZR Concentrations
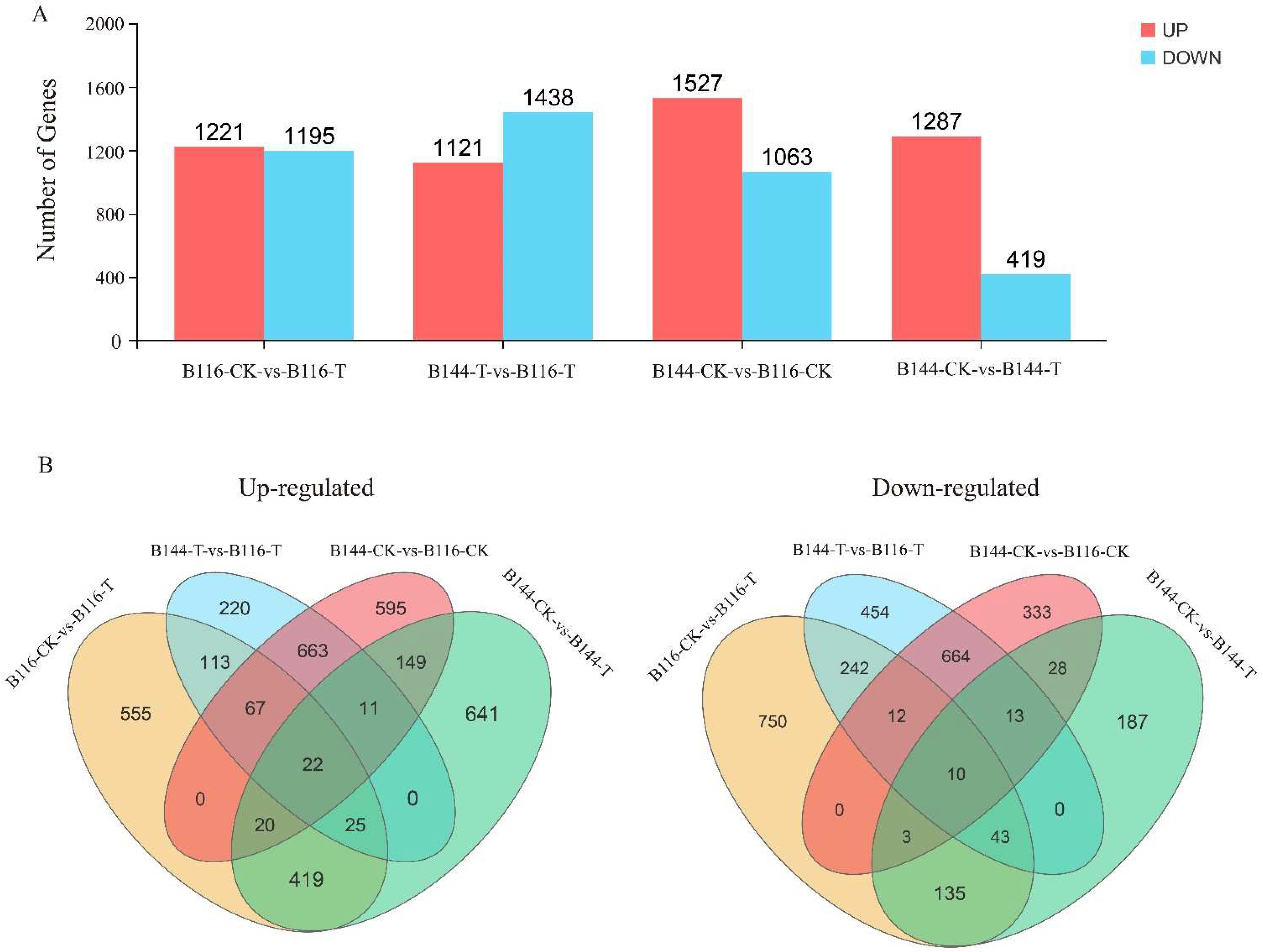
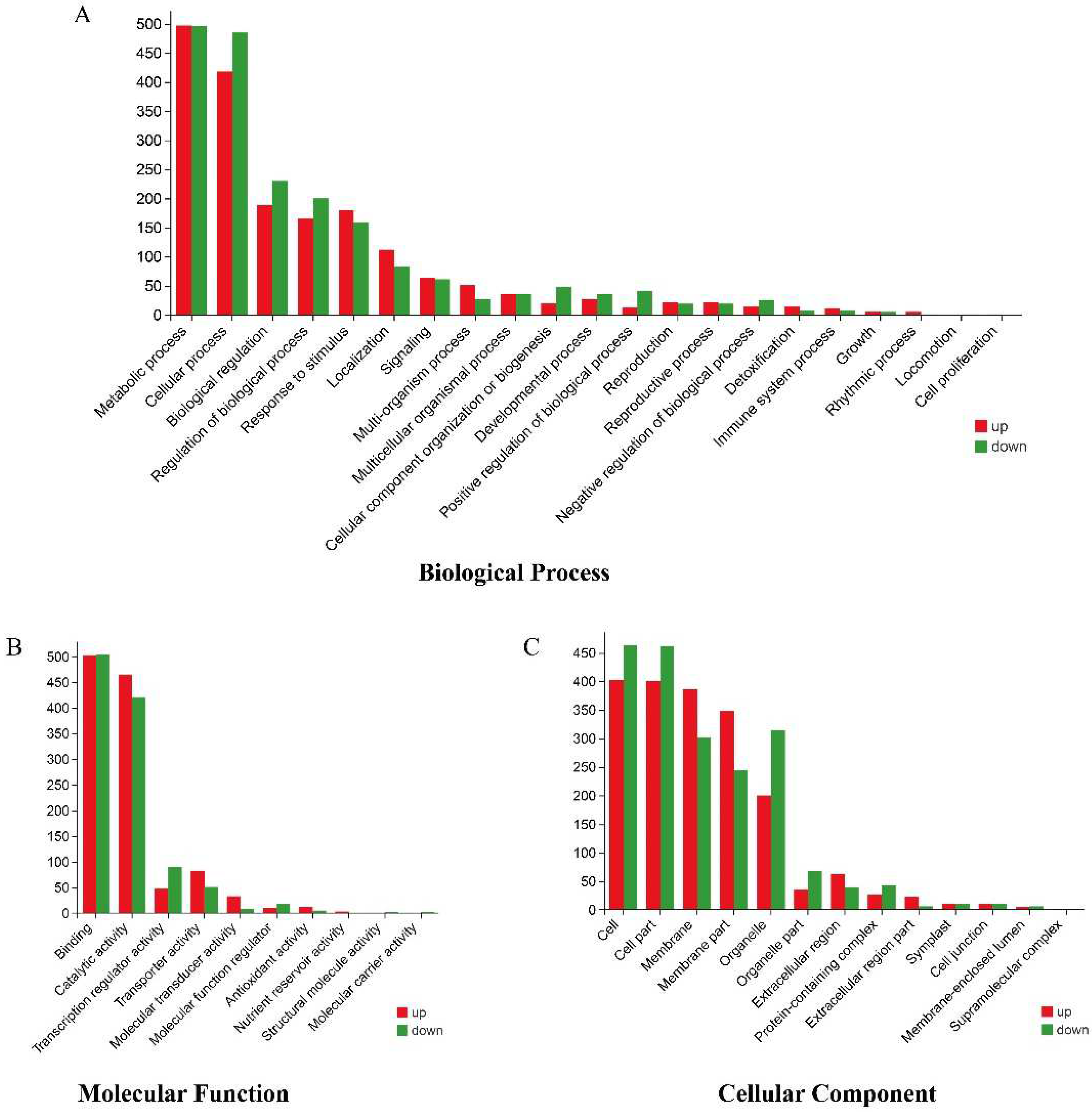
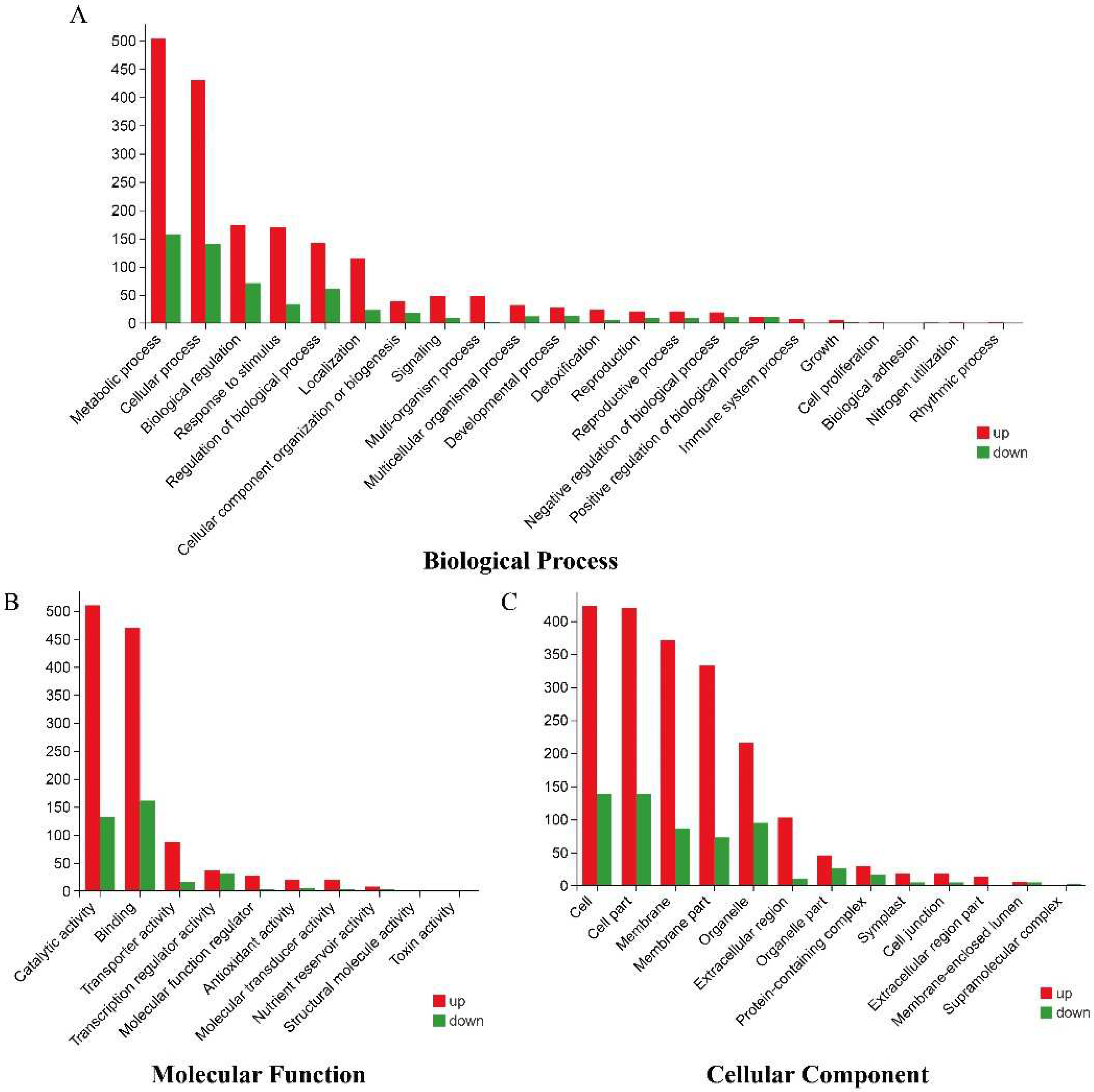
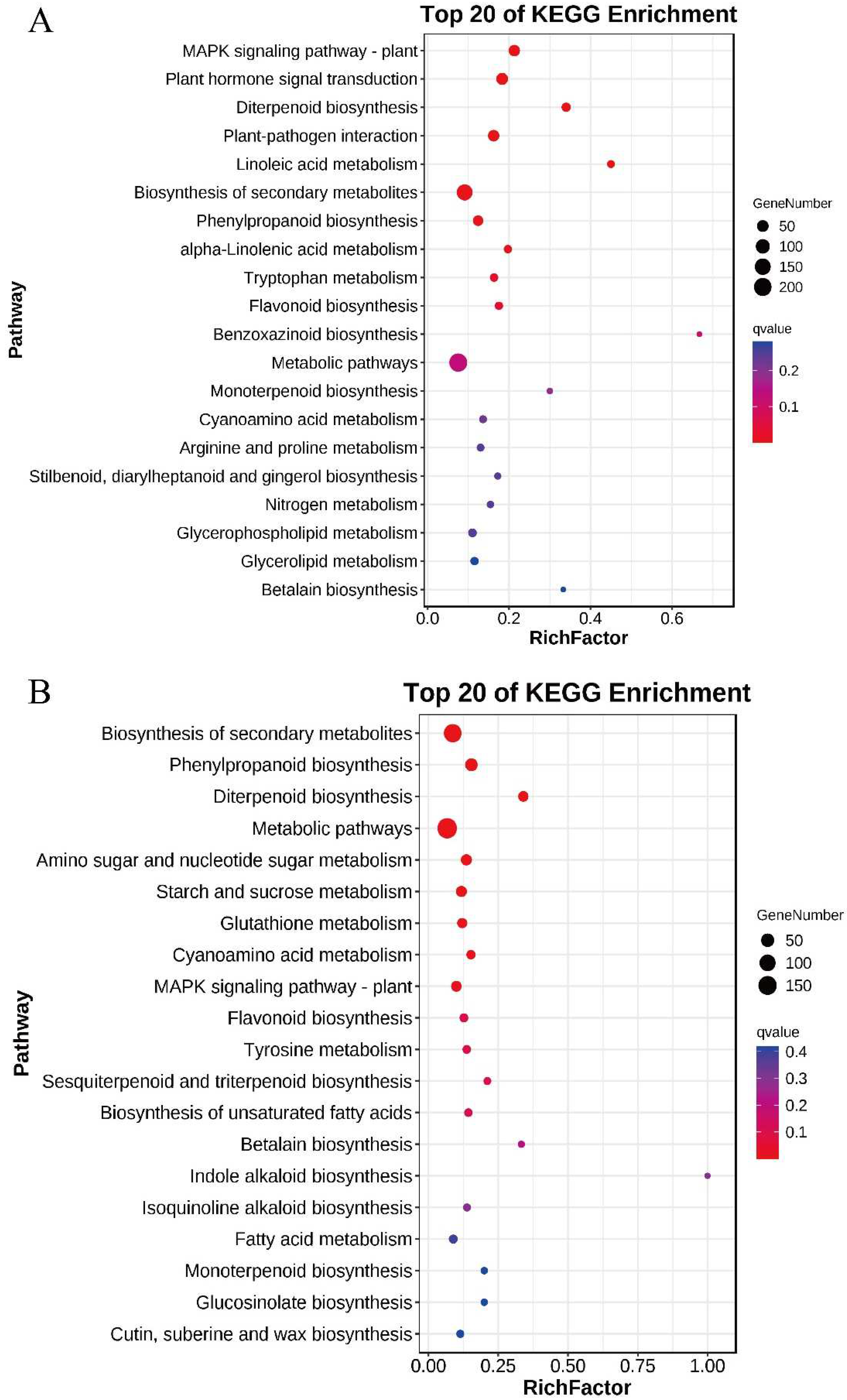
3.5. Transcriptome Sequencing and Functional Identification of B116 and B144
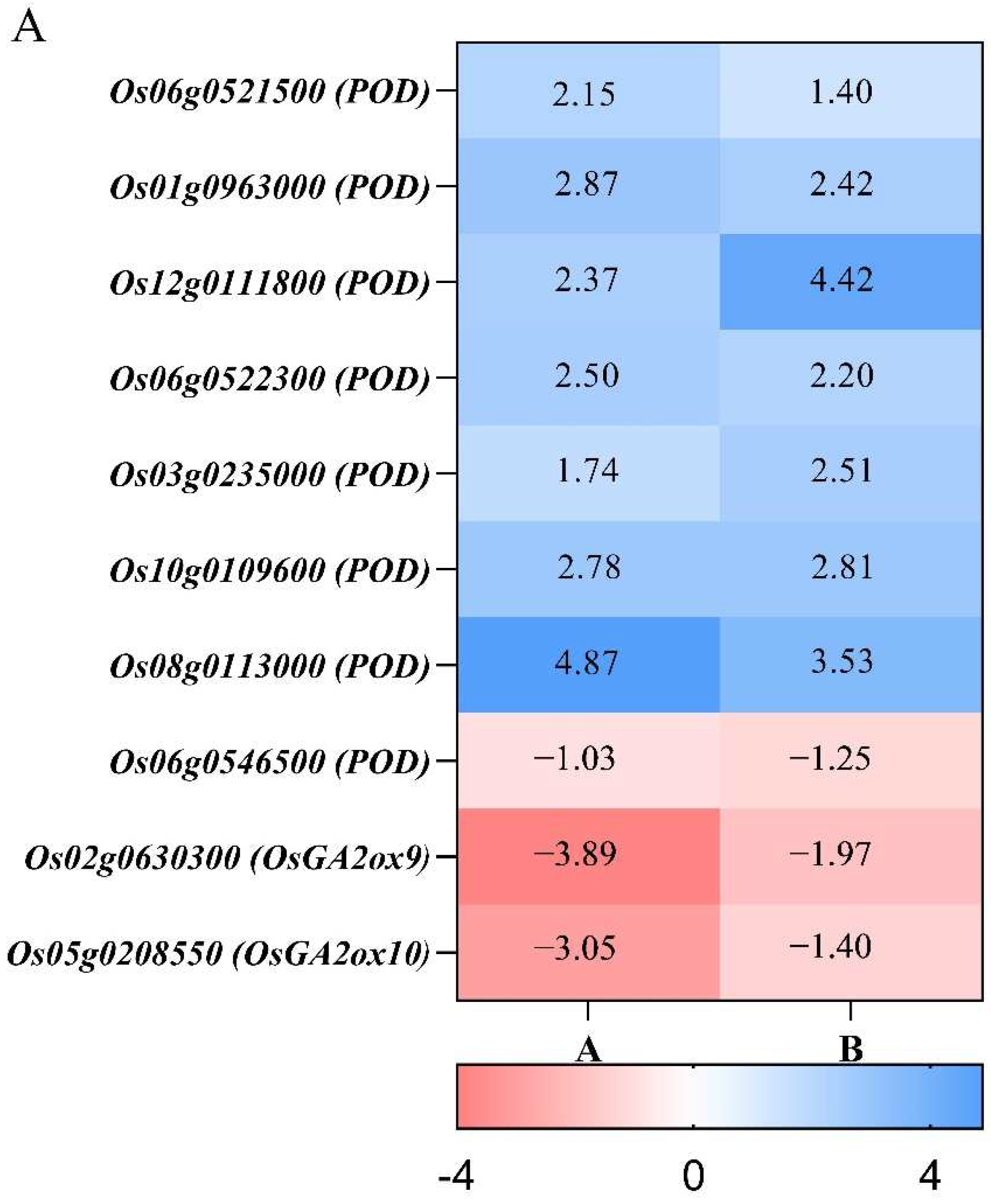
3.6. Differences in Antioxidant Enzyme and Gibberellin Metabolism during Compensatory Growth of Two Rice Genotypes
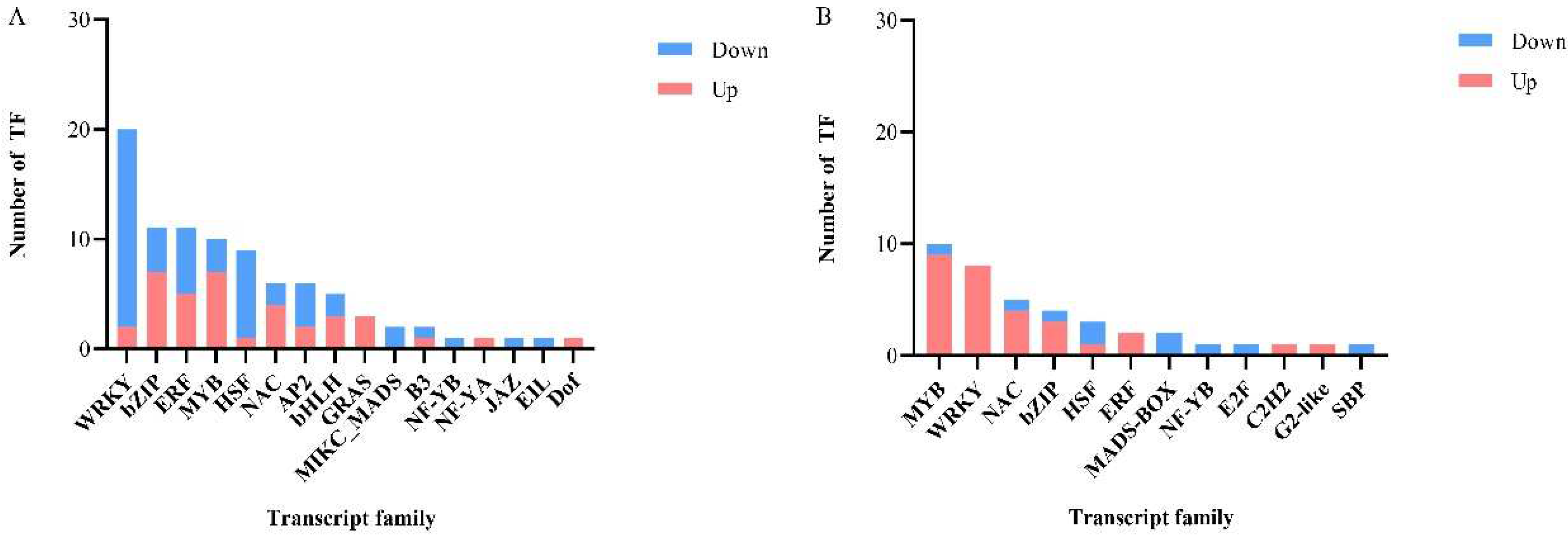
3.7. Regulation of Differentially Expressed Genes by Transcription Factors (TFs)
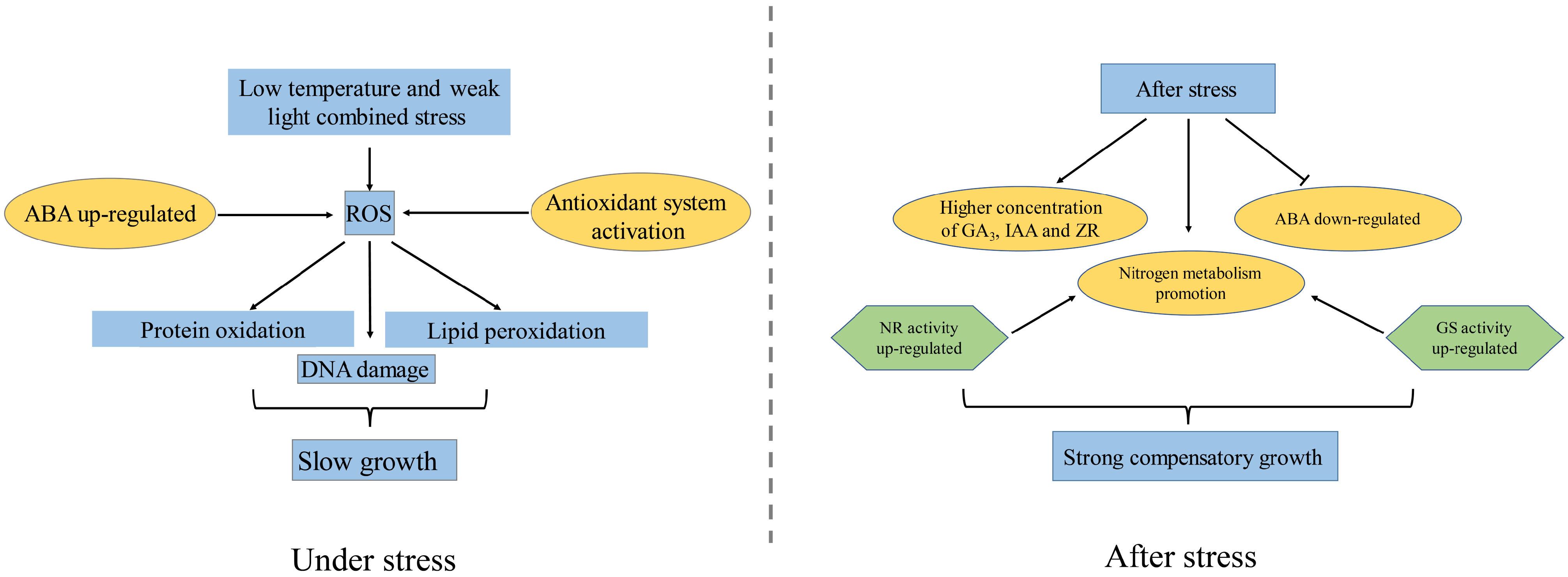
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Hallerman, E.M.; Liu, Q.; Wu, K.; Peng, Y. The development and status of Bt rice in China. Plant Biotechnol. J. 2016, 14, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, D.; Ladha, J.K.; Rana, D.S.; Jat, M.L.; Gathala, M.K.; Yadav, S.; Rao, A.N.; Ramesha, M.S.; Raman, A. A global analysis of alternative tillage and crop establishment practices for economically and environmentally efficient rice production. Sci. Rep. 2017, 7, 9342. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.-H.; Li, X.-Y.; Chen, H.-Z.; Wu, H.; Ouyang, Y.-Y.; Yu, J.-Q.; Huang, B.-M.; Luo, X. Effects of side deep placement of nitrogen fertilizer on yield and nitrogen utilization efficiency of mechanized direct-seeded rice. J. Nucl. Agric. Sci. 2020, 34, 2051–2058. [Google Scholar]
- Zhang, H.-C.; Xing, Z.-P.; Weng, W.-A.; Tian, J.-Y.; Tao, Y.; Cheng, S.; Hu, Q.; Hu, Y.-J.; Guo, B.-W.; Wei, H.-Y. Growth characteristics and key techniques for stable yield of growth constrained direct seeding rice. Sci. Agric. Sin. 2021, 54, 1322–1337. [Google Scholar]
- Luo, X.-W.; Wang, Z.-M.; Zeng, S.; Zang, Y.; Yang, W.-W.; Zhang, M.-H. Recent advances in mechanized direct seeding technology for rice. J. S. China Agric. Univ. 2019, 40, 1–13. [Google Scholar]
- Fang, J.-C.; Tsai, Y.C.; Chou, W.-L.; Liu, H.-Y.; Lu, C.-A. A CCR4-associated factor 1, OsCAF1B, confers tolerance of low-temperature stress to rice seedlings. Plant Mol. Biol. 2021, 105, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.-H.; Hu, B.-L.; Xiao, Y.-Q.; Li, X.; Peng, Z.-Q.; Wang, Y. Research advances on important stress tolerant traits and its molecular mechanisms in Dongxiang wild rice. China Rice 2017, 23, 12–18. [Google Scholar]
- Zhu, D.-F.; Zhang, Y.-P. Cause analysis and control measures of large-area rotten seedling caused by spring cold in early rice. China Rice 2020, 26, 10–12. [Google Scholar]
- Sharma, N.; Reinke, R.; Sacks, E.J. Comparison of methods to evaluate rice (Oryza sativa) germplasm for tolerance to low temperature at the seedling stage. Agronomy 2021, 11, 385. [Google Scholar] [CrossRef]
- Ahmed, G.G.; Habiba; Zheng, X.Z.; Miao, Y. Low Light/Darkness as Stressors of Multifactor-Induced Senescence in Rice Plants. Int. J. Mol. Sci. 2021, 22, 3936. [Google Scholar]
- Deng, F.; Li, Q.; Chen, H.; Zeng, Y.; Ren, W. Relationship between chalkiness and the structural and thermal properties of rice starch after shading during grain-filling stage. Carbohydr. Polym. 2021, 252, 117212. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-L.; Zhang, S.-H.; Yang, T.-F.; Zeng, Z.-C.; Huang, Z.-H.; Liu, Q.; Wang, X.-F.; Leach, J.; Leung, H.; Liu, B. Global transcriptional profiling of a cold-tolerant rice variety under moderate cold stress reveals different cold stress response mechanisms. Physiol. Plant 2015, 154, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-T.; Wang, W.; Mao, B.-G.; Chu, C.-C. Cold stress tolerance in rice: Physiological changes, molecular mechanism, and future prospects. Hereditas 2018, 40, 171–185. [Google Scholar] [PubMed]
- Wang, W.-Q.; Chen, Q.; Hussain, S.; Mei, J.-H.; Dong, H.-L.; Peng, S.-B.; Huang, J.-L.; Cui, K.-H.; Nie, L.-X. Pre-sowing seed treatments in direct-seeded early rice: Consequences for emergence, seedling growth and associated metabolic events under chilling stress. Sci. Rep. 2016, 6, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Zhang, W.-X.; Jiang, L.-G.; Zou, Y.-B. Impact of temperature changes on early-rice productivity in a subtropical environment of China. Field Crops Res. 2013, 146, 10–15. [Google Scholar] [CrossRef]
- Cui, X.-A.; Wang, Y.-W.; Wu, J.-X.; Han, X.; Gu, X.-F.; Lu, T.-G.; Zhang, Z.-G. The rna editing factor dua1 is crucial to chloroplast development at low temperature in rice. New Phytol. 2018, 221, 834–849. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.-G.; Yin, C.-Y.; Strasser, R.-J.; Govindjee; Yang, C.-L.; Qiang, S. Reactive oxygen species from chloroplasts contribute to 3-acetyl-5-isopropyltetramic acid-induced leaf necrosis of Arabidopsis thaliana. Plant Physiol. Biochem. 2012, 52, 38–51. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Shi, Y.; Huy, P.; Liu, Y.; Cao, S.-Y.; Zhang, Z.-H.; Chu, C.-C.; Michael, R.-S. Glycosyltransferase OsUGT90A1 helps protect the plasma membrane during chilling stress in rice. J. Exp. Bot. 2020, 9, 2723–2739. [Google Scholar] [CrossRef]
- Ruelland, E.; Zachowski, A. How plants sense temperature. Environ. Exp. Bot. 2010, 69, 225–232. [Google Scholar] [CrossRef]
- Adamski, J.M.; Rosa, L.M.G.; Menezes, P.C.R.; Pinheiro, C.L.; Fett, J.P.; Sperotto, R.A. Photosynthetic activity of indica rice sister lines with contrasting cold tolerance. Physiol. Mol. Biol. Plants 2020, 26, 955–964. [Google Scholar] [CrossRef]
- Wang, W.-X.; Chen, L.-M.; Liu, Y.-Q.; Zeng, Y.-J.; Wu, Z.-M.; Tan, X.-M.; Shi, Q.-H.; Pan, X.-H.; Zeng, Y.-H. Effects of humidity-cold combined stress at the seedling stage in direct-seeded indica rice. Environ. Exp. Bot. 2021, 191, 104617. [Google Scholar] [CrossRef]
- Yang, D.; Duan, L.-S.; Xie, H.-A.; Huang, T.-X. The response and correlation analysis of rice seedlings growth to low-light stress. China Agric. Sci. Bull. 2011, 27, 70–79. [Google Scholar]
- Wang, C.-Z.; Gao, L.-M.; Sun, Y.-M.; Wang, B.; Guo, S.-W. The effect of weak light stress on leaf photosynthetic characteristics in super hybrid rice and conventional rice at tillering stage. J. Nanjing Agric. Univ. 2019, 42, 111–117. [Google Scholar]
- Xu, G.-L.; Liu, J.; Liang, C.-G.; Wang, Y.; Ding, C.-B.; Li, T. Changes of nitrogen metabolism enzyme activities and protein content in response to low light during the seed filling stage in rice. J. Zhejiang Univ. 2016, 42, 53–62. [Google Scholar]
- Wang, W.-X.; Du, J.; Chen, L.-M.; Zeng, Y.-J.; Tan, X.-M.; Shi, Q.-H.; Pan, X.-H.; Wu, Z.-M.; Zeng, Y.-H. Transcriptomic, proteomic, and physiological comparative analyses of flooding mitigation of the damage induced by low-temperature stress in direct seeded early indica rice at the seedling stage. BMC Genom. 2021, 22, 176. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-W.; Chen, H.-C.; Liu, L.-Y.; Chang, M.-C. Comparative Transcriptome Analysis of Shoots and Roots of TNG67 and TCN1 Rice Seedlings under Cold Stress and Following Subsequent Recovery: Insights into Metabolic Pathways, Phytohormones, and Transcription Factors. PLoS ONE 2015, 10, e0131391. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.-G.; Duan, L.-C.; Wu, Z.-M.; Li, H.-J. Analysis of disaster characteristics caused by late spring cold and cold dew wind in double-season direct-seeding rice in Jiangxi province. Acta Agric. Jiangxi 2021, 33, 65–71. [Google Scholar]
- Zhou, T.; Chen, S.-Q.; Du, X.-D. Effect of different herbicides on japonica rice seedling. Heilongjiang Agric. Sci. 2015, 12, 55–59. [Google Scholar]
- Yi, Z.-H.; Zhu, D.-F.; Wang, Y.-L.; Hu, G.-H.; Zhang, Y.-P.; Xiang, J.; Zhang, Y.-K.; Chen, H.-Z. Advances of rice growth response to drought and its compensatory effects. China Rice 2020, 26, 1–9. [Google Scholar]
- Zhang, Y.-Y. Compensatory Effects of Rewatering after Drought on Growth and Physiology of Platycladus Orientalis. Master’s Thesis, Northwest A&F University, Yangling, China, 29 May 2022. [Google Scholar]
- Avramova, V.; Abdelgawad, H.; Zhang, Z.; Fotschki, B.; Casadevall, R.; Vergauwen, L.; Knapen, D.; Taleisnik, E.; Guisez, Y.; Asard, H.; et al. Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone. Plant Physiol. 2015, 169, 1382–1396. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, E.; Hsiao, T.C.; Henderson, D.W. Immediate and subsequent growth responses of maize leaves to changes in water status. Plant Physiol. 1971, 48, 631–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandran, A.K.; Priatama, R.A.; Kumar, V.; Xuan, Y.; Je, B.I.; Kim, C.M.; Jung, K.H.; Han, C.D. Genome-wide transcriptome analysis of expression in rice seedling roots in response to supplemental nitrogen. J. Plant Physiol. 2016, 200, 62–75. [Google Scholar] [CrossRef]
- Xiong, Q.-Q.; Tang, G.-P.; Zhong, L.; He, H.-H.; Chen, X.-R. Response to Nitrogen Deficiency and Compensation on Physiological Characteristics, Yield Formation, and Nitrogen Utilization of Rice. Front. Plant Sci. 2018, 9, 1075. [Google Scholar] [CrossRef] [PubMed]
- Standardization Administration of China: National Standard of the People’s Republic of China for Inversion Weather Indicators. 2017. Available online: http://www.gb688.cn/bzgk/gb/newGbInfo?hcno=A9917F534E4F8CC0C49BB62591E3C6E5 (accessed on 30 May 2022).
- Chen, J.-H.; Wu, X.; Song, J.-X.; Xing, G.-P.; Liang, L.; Yin, Q.-L.; Guo, A.-J.; Cui, J. Transcriptomic and physiological comparsion of the short-term responses of two Oryza sativa L. varieties to cadmium. Environ. Exp. Bot. 2021, 181, 104292. [Google Scholar] [CrossRef]
- Yu, Z.-P.; Duan, X.-B.; Luo, L.; Dai, S.-J.; Ding, Z.-J.; Xia, G.-M. How Plant Hormones Mediate Salt Stress Responses. Trends Plant Sci. 2020, 25, 1117–1130. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Meng, J.; Liang, X.; Yang, E.; Chen, W.-F. Biochar’s Leacheates Affect the Abscisic Acid Pathway in Rice Seedlings Under Low Temperature. Front. Plant Sci. 2021, 12, 646910. [Google Scholar] [CrossRef] [PubMed]
- Jurado, S.; Abraham, Z.; Manzano, C.; López-Torrejón, G.; Pacios, L.-F.; Del Pozo, J.-C. The Arabidopsis cell cycle F-box protein SKP2A binds to auxin. Plant Cell. 2010, 22, 3891–3904. [Google Scholar] [CrossRef]
- Kantharaj, V.; Ramasamy, N.-K.; Yoon, Y.-E.; Cheong, M.-S.; Kim, Y.-N.; Lee, K.-A.; Kumar, V.; Choe, H.; Kim, S.-Y.; Chohra, H.; et al. Auxin-Glucose Conjugation Protects the Rice (Oryza sativa L.) Seedlings Against Hydroxyurea-Induced Phytotoxicity by Activating UDP-Glucosyltransferase Enzyme. Front. Plant Sci. 2022, 12, 767004. [Google Scholar] [CrossRef]
- Wang, D.; Chu, G.; Chen, S.; Xu, C.-M.; Chen, L.-P.; Zhang, X.-F. Innovation and development of mechanized wet direct seeding technology for rice. China Rice 2020, 26, 19–21+25. [Google Scholar]
- Velitchkova, M.; Popova, A.-V.; Faik, A.; Gerganova, M.; Ivanov, A.-G. Low temperature and high light dependent dynamic photoprotective strategies in Arabidopsis thaliana. Physiol. Plant 2020, 170, 93–108. [Google Scholar] [CrossRef]
- Hashimoto, M.; Kisseleva, L.; Sawa, S.; Furukawa, T.; Komatsu, S.; Koshiba, T. A novel rice PR10 protein, RSOsPR10, specifically induced in roots by biotic and abiotic stresses, possibly via the jasmonic acid signaling pathway. Plant Cell Physiol. 2004, 45, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-L.; Tao, L.-Y.; Liu, T.-T.; Zhang, X.-H.; Wang, W.; Song, J.-M.; Yu, C.-L.; Peng, X.-L. Nitrogen application after low-temperature exposure alleviates tiller decrease in rice. Environ. Exp. Bot. 2019, 158, 205–214. [Google Scholar] [CrossRef]
- Liu, Z.-L.; Meng, J.-R.; Sun, Z.-F.; Su, J.-K.; Luo, X.-Y.; Song, J.-M.; Li, P.-F.; Sun, Y.-K.; Yu, C.-L.; Peng, X.-L. Zinc application after low temperature stress promoted rice tillers recovery: Aspects of nutrient absorption and plant hormone regulation. Plant Sci. 2022, 314, 111104. [Google Scholar] [CrossRef]
- Fanourakis, D.; Aliniaeifard, S.; Sellin, A.; Giday, H.; Tsaniklidis, G. Stomatal behavior following mid- or long-term exposure to high relative air humidity: A review. Plant Physiol. Biochem. 2020, 153, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-X.; Chen, L.-M.; Wang, H.-X.; Liu, Y.-Q.; Wu, Z.-M.; Zeng, Y.-J.; Tan, X.-M.; Pan, X.-H.; Shi, Q.-H.; Zeng, Y.-H. Study on physiological characteristics behind mitigative effects of flooding on low temperature-caused chilling damage to direct seeded early indica rice at the seedling stage. Chin. J. Rice Sci. 2021, 35, 166–176. [Google Scholar]
- Yang, B.; Ma, H.-Y.; Wang, X.-M. Improvement of nitrogen accumulation and metabolism in rice (Oryza sativa L.) by the endophyte Phomopsis liquidambari. Plant Physiol. Biochem. 2014, 82, 172–182. [Google Scholar] [CrossRef]
- Tabuchi, M.; Abikom, T.; Yamaya, T. Assimilation of ammonium ions and reutilization of nitrogen in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 2319–2327. [Google Scholar] [CrossRef]
- Xu, H.; Shang, J.; Liu, M.-Y.; Zhang, F.; Li, Y.-Y. Research progress of enzymes related to nitrogen metabolism. J. Anhui Agric. Sci. 2022, 50, 17–20. [Google Scholar]
- Jyotirmaya, M.; Anuradha, S.; Vikram, J.; Aashish, R. High photosynthesis rate in two wild rice species is driven by leaf anatomy mediating high Rubisco activity and electron transport rate. J. Exp. Bot. 2021, 72, 7119–7135. [Google Scholar]
- Mao, S.; Yuji, S.; Youshi, T.; Wataru, Y.; Amane, M. Co-overproducing Rubisco and Rubisco activase enhances photosynthesis in the optimal temperature range in rice. Plant Physiol. 2020, 185, 108–119. [Google Scholar]
- Muhammad, A.-K.; Muhammad, H.; Sajjad, A.; Murtaza, K.; Byung, W.-Y.; Sang, M.-K.; In, J.-L. Rescues plant growth under salinity stress via regulating gene expression, endogenous hormones, and antioxidant system of Oryza sativa L. Front. Plant Sci. 2021, 12, 665590. [Google Scholar]
- Waadt, R.; Seller, C.-A.; Hsu, P.-K.; Takahashi, Y.; Munemasa Schroeder, J.-I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Wuddineh, W.-A.; Mazarei, M.; Zhang, J.-Y.; Poovaiah, C.-R.; Mann, D.-G.; Ziebell, A.; Sykes, R.-W.; Davis, M.-F.; Udvardi, M.-K.; Stewart, C.-N., Jr. Identification and overexpression of gibberellin 2-oxidase (GA2ox) in switchgrass (Panicum virgatum L.) for improved plant architecture and reduced biomass recalcitrance. Plant Biotechnol. J. 2015, 13, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Zhang, Y.; Zhang, X.; Yao, D.-P.; He, R.-H.; Chen, Z.; Zhang, L.; Bo, B. Chang Characteristics of phytohormones contents in rice seedlings under salt stress. Hunan Agric. Sci. 2021, 12, 30–35. [Google Scholar]
- Liu, C.-F. Effects of Artificial Flooding on the Growth and Morphological Characteristics of 15 Tree Species Seedlings. Ph.D. Thesis, Nanjing Forestry University, Nanjing, China, 15 May 2022. [Google Scholar]
- Zhang, S.-N.; Jia, Y.; Qu, Z.-J.; Yang, L.; Li, X.; Zhang, Y.; Wang, Z.; Zhao, H.-W. Effect of cold water irrigation at booting stage on physiological indexes and chloroplast ultrastructure of japonica rice in cold region. Acta Agric. Boreali-Occident. Sin. 2020, 30, 886–897. [Google Scholar]
- Yu, X.-F.; Qiu, X.-L.; Zhang, S.-H.; Zhang, Y.-F.; Zeng, M.; Zeng, L.-Q.; Yang, S.-M. Effects of low temperature and nitrogen coupling on endogenous hormones of rice. J. Northwest A. F. Univ. 2020, 48, 73–80. [Google Scholar]
- Lo, S.-F.; Yang, S.-Y.; Chen, K.-T.; Hsing, Y.-L.; Zeevaart, J.-A.-D.; Chen, L.-J.; Yu, S.-M. A novel class of gibberellin 2-oxidases control semidwarfism, tillering, and root development in Rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Zhong, L.; Fu, X.; Huang, S.; Fu, H.; Shi, X.; Hu, L.; Cai, Y.; He, H.; Chen, X. Physiological and Transcriptomic Analyses Reveal the Mechanisms of Compensatory Growth Ability for Early Rice after Low Temperature and Weak Light Stress. Plants 2022, 11, 2523. https://doi.org/10.3390/plants11192523
Wang H, Zhong L, Fu X, Huang S, Fu H, Shi X, Hu L, Cai Y, He H, Chen X. Physiological and Transcriptomic Analyses Reveal the Mechanisms of Compensatory Growth Ability for Early Rice after Low Temperature and Weak Light Stress. Plants. 2022; 11(19):2523. https://doi.org/10.3390/plants11192523
Chicago/Turabian StyleWang, Hui, Lei Zhong, Xiaoquan Fu, Shiying Huang, Haihui Fu, Xiang Shi, Lifang Hu, Yicong Cai, Haohua He, and Xiaorong Chen. 2022. "Physiological and Transcriptomic Analyses Reveal the Mechanisms of Compensatory Growth Ability for Early Rice after Low Temperature and Weak Light Stress" Plants 11, no. 19: 2523. https://doi.org/10.3390/plants11192523