
Research Progress on the Effects of Selenium on the Growth and Quality of Tea Plants
 ,
,
Abstract
:1. Introduction
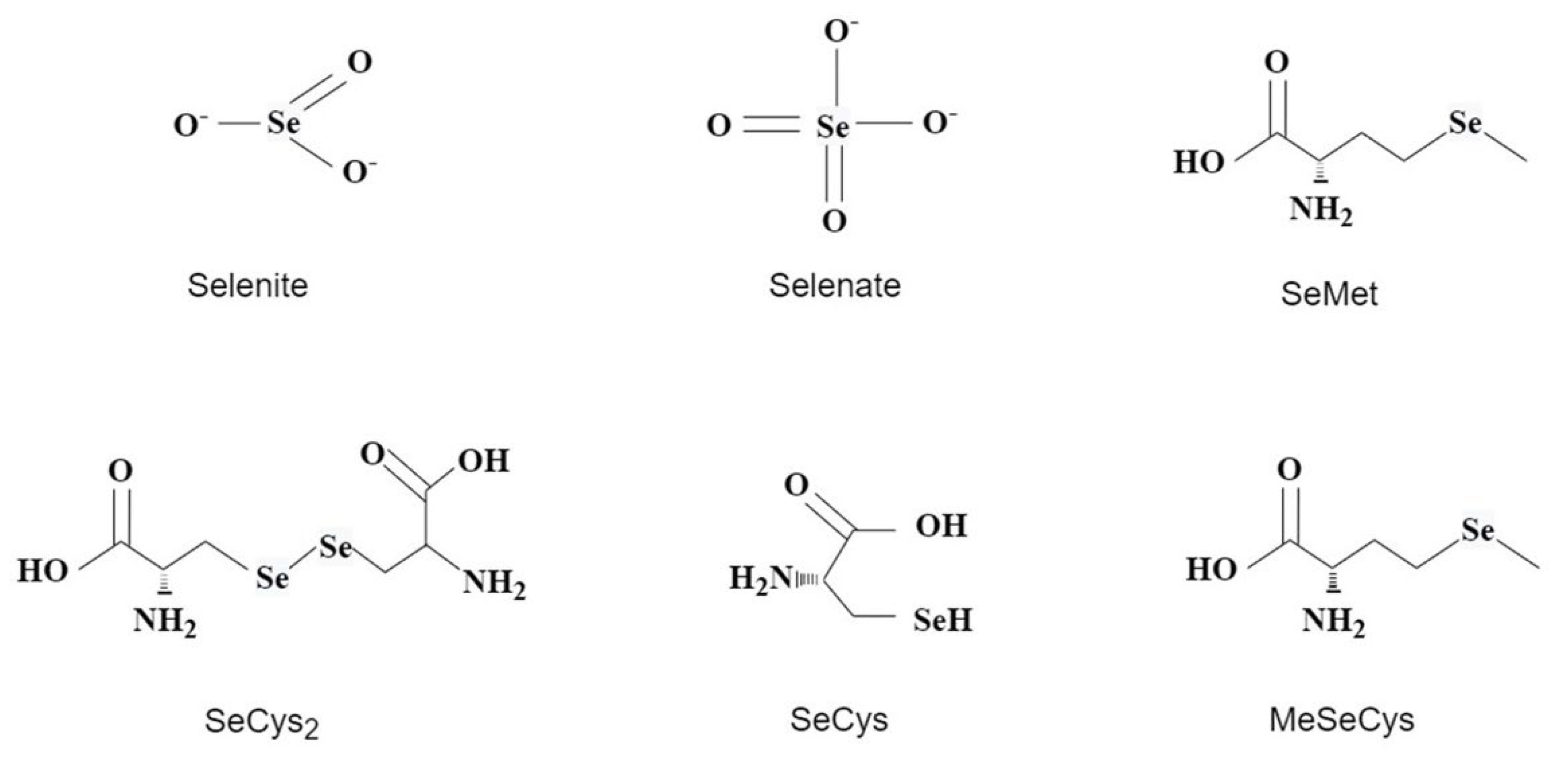
2. Selenium Uptake and Metabolism in Tea Plant
3. Effect of Selenium on Tea Plant Growth
3.1. Effect of Selenium on Tea Plant Biomass
3.2. Effect of Selenium on the Uptake of Mineral Elements by Tea Plant
4. Effects of Selenium on Tea Plant Quality
4.1. Effect of Exogenous Selenium on the Selenium Content of Tea Plants
4.2. Effect of Exogenous Selenium on Selenium Forms in Tea Plants
4.3. Effect of Exogenous Selenium on Tea Polyphenols in Tea Leaves
4.4. Effect of Exogenous Selenium on Flavonoids in Tea Leaves
4.5. Effect of Exogenous Selenium on Amino Acids in Tea Leaves
4.6. Effect of Exogenous Selenium on Other Nutrients in Tea Leaves
4.7. Effect of Exogenous Selenium on Aroma Composition in Tea Leaves
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Razaghi, A.; Poorebrahim, M.; Sarhan, D.; Björnstedt, M. Selenium stimulates the antitumour immunity: Insights to future research. Eur. J. Cancer 2021, 155, 256–267. [Google Scholar] [CrossRef]
- Alcolea, V.; Pérez, S. Selenium as an interesting option for the treatment of Chagas disease: A review. Eur. J. Med. Chem. 2020, 206, 112673. [Google Scholar] [CrossRef] [PubMed]
- Mendil, D.; Demirci, Z.; Uluozlu, O.D.; Tuzen, M.; Soylak, M. A new separation and preconcentration method for selenium in some foods using modified silica gel with 2,6-diamino-4-phenil-1,3,5-triazine. Food Chem. 2017, 221, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Liu, G.; Yousaf, B.; Ali, M.U.; Abbas, Q.; Munir, M.A.M.; Mian, M.M. Developmental selenium exposure and health risk in daily foodstuffs: A systematic review and meta-analysis. Ecotoxicol. Environ. Saf. 2018, 149, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ho, C.T.; Long, P.; Meng, Q.; Zhang, L.; Wan, X. Preventive efficiency of green tea and its components on nonalcoholic fatty liver disease. J. Agric. Food Chem. 2019, 67, 5306–5317. [Google Scholar] [CrossRef]
- Gao, Y.; Xu, Y.; Ruan, J.; Yin, J. Selenium affects the activity of black tea in preventing metabolic syndrome in high-fat diet-fed Sprague–Dawley rats. J. Sci. Food Agric. 2019, 100, 225–234. [Google Scholar] [CrossRef]
- Li, J.; Weng, G.; Pan, Y.; Li, C.; Wang, N. A scientometric review of tourism carrying capacity research: Cooperation, hotspots, and prospect. J. Clean. Prod. 2021, 325, 129278. [Google Scholar] [CrossRef]
- Gupta, U.C.; Winter, K.A.; McRae, K.B. Selenium enrichment of crops through foliar applications. Can. J. Soil Sci. 1988, 68, 519–526. [Google Scholar] [CrossRef]
- Zhang, L.H.; Song, Y.; Guo, B.; Fan, Y.; Huang, X.; Mao, K.; Liang, Z.; Hu, X.; Sun, Y.; Fang, X.; et al. Benefit–risk assessment of dietary selenium and its associated metals intake in China (2017–2019): Is current selenium-rich agro-food safe enough? J. Hazard. Mater. 2020, 398, 123224. [Google Scholar] [CrossRef]
- Cao, D.; Jin, X.; Gong, Z.; Ma, L.; Zheng, L. Advances in the mechanism of selenium uptake and tolerance in tea plants. Biotechnology. 2017, 7, 445–449. [Google Scholar]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.; Li, X.; Guo, L.; Wang, L.; Hao, X.; Zeng, J. Integrative transcriptome and proteome analysis reveals the absorption and metabolism of selenium in tea plants [Camellia sinensis (L.) o. Kuntze]. Front. Plant Sci. 2022, 13, 848349. [Google Scholar] [CrossRef] [PubMed]
- Eichert, T.; Goldbach, H.E. Equivalent pore radii of hydrophilic foliar uptake routes in stomatous and astomatous leaf surfaces--further evidence for a stomatal pathway. Physiol. Plant. 2008, 132, 491–502. [Google Scholar] [CrossRef]
- Kaiser, H. Stomatal uptake of mineral particles from a sprayed suspension containing an organosilicone surfactant. J. Plant Nutr. Soil Sc. 2014, 177, 869–874. [Google Scholar] [CrossRef]
- Guo, X.; Ji, Q.; Rizwan, M.; Li, H.; Li, D.; Chen, G. Effects of biochar and foliar application of selenium on the uptake and subcellular distribution of chromium in Ipomoea aquatica in chromium-polluted soils. Ecotox. Environ. Safe. 2020, 206, 111184. [Google Scholar] [CrossRef]
- Mazej, D.; Falnoga, I.; Veber, M.; Stibilj, V. Determination of selenium species in plant leaves by HPLC–UV–HG-AFS. Talanta 2006, 68, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Trippe, R.C., 3rd; Pilon Smits, E.A.H. Selenium transport and metabolism in plants: Phytoremediation and biofortification implications. J. Hazard Mater. 2021, 404, 124178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, B.; Li, W.; Che, R.; Deng, K.; Li, H.; Chu, C. OsPT2, a phosphate transporter, is involved in the active uptake of selenite in rice. New Phytol. 2014, 201, 1183–1191. [Google Scholar] [CrossRef]
- Hu, Q.; Xu, J.; Pang, G. Effect of selenium on the yield and quality of green tea leaves harvested in early spring. Agric. Food Chem. 2003, 51, 3379–3381. [Google Scholar] [CrossRef]
- Huang, S.; Kan, Y.; Tang, Y. Effect of bioorganic selenium on yield, quality and selenium content of green tea. J. Tea Commun. 2020, 47, 610–616. [Google Scholar]
- Wu, H.; Zhang, H.; Ren, Z.H. Effects of Foliar Application of Nano-Selenium on Photosynthetic Characteristics and Yield of Tea. Shandong Agric. Sci. 2021, 53, 64–68. [Google Scholar]
- Xu, J.; Zhu, F.S.; Yang, L.; Cheng, Y.; Hu, G.; Pan, Q. The influence of selenium on the antioxidant activity of green tea. J. Sci. Food Agric. 2003, 83, 451–455. [Google Scholar] [CrossRef]
- Liu, K.; Li, S.; Han, J.; Zeng, X.; Ling, M.; Mao, J.; Li, Y.; Jiang, J. Effect of selenium on tea (Camellia sinensis) under low temperature: Changes in physiological and biochemical responses and quality. Environ. Exp. Bot. 2021, 188, 104475. [Google Scholar] [CrossRef]
- Ruan, J. The Impact of pH and Calcium on the Uptake of Fluoride by Tea Plants (Camellia sinensis L.). Ann. Bot. 2004, 93, 97–105. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Comparative effects of selenite and selenate on growth and selenium accumulation in lettuce plants under hydroponic conditions. Plant Growth Regul. 2013, 70, 149–157. [Google Scholar] [CrossRef]
- Huang, Y.; Xu, J.; Hu, Q. Effect of Selenium on Preservation Quality of Green Tea during Autumn Tea-Processing Season. Food Chem. 2005, 53, 7444–7447. [Google Scholar] [CrossRef]
- Yang, T.; Li, H.; Hu, X.; Li, J.; Hu, J.; Liu, R.; Deng, Z. Effects of fertilizing with n, p, Se, and Zn on regulating the element and functional component contents and antioxidant activity of tea leaves planted in red soil. Food Chem. 2014, 62, 3823–3830. [Google Scholar] [CrossRef]
- Qin, Y.Y.; Wang, Y.R.; Shi, P.T. Effect of foliar selenium spray on the selenium and mineral element content of tea tree leaves. J. South. Agric. 2019, 50, 622–627. [Google Scholar]
- Cai, H.; Dong, Y.; Li, Y.; Li, D.; Peng, C.; Zhang, X.Z. Physiological and cellular responses to fluoride stress in tea (Camellia sinensis) leaves. Acta Physiol. Plant. 2016, 38, 371–374. [Google Scholar] [CrossRef]
- Niu, H.; Zhan, W.K.; Xu, C.; Peng, C.; Hou, Y.; Li, R.; Hou, X.; Wan, H. Selenium treatment modulates fluoride distribution and mitigates fluoride stress in tea plant (Camellia sinensis (L.) O. Kuntze). Environ. Pollut. 2020, 267, 115603. [Google Scholar] [CrossRef]
- Kim, D.; Chung, D.; Bai, S.; Kim, H.; Lee, Y. Effects of soil selenium supplementation level on selenium contents of green tea leaves and milk vetch. Prev. Nutr. Food Sci. 2007, 12, 35–39. [Google Scholar] [CrossRef]
- Jiang, F.Y.; Wu, Z.D.; Zhang, W.J. Effect of soil application of sodium selenite on the selenium content of oolong tea. Acta Tea Sinica. 2017, 58, 13–16. [Google Scholar]
- Winkel, L.; Vriens, B.; Jones, G.; Schneider, E.L.; Pilon-Smits, G.; Bañuelos, G.S. Selenium cycling across Soil-Plant-Atmosphere interfaces: A critical review. Nutrients 2015, 7, 4199–4239. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Liu, Y.; Ma, L.; Jin, X.; Guo, G.; Tan, R.; Liu, W. Transcriptome analysis of differentially expressed genes involved in selenium accumulation in tea plant (Camellia sinensis). PLoS ONE. 2018, 13, e197506. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Huang, Y.; Zhou, Z.Z. Effect of different concentrations of sodium selenate on the growth and physiological indicators of tea plants. Guihaia 2021, 41, 183–194. [Google Scholar]
- Chen, S.; Zhu, S.; Lu, D. Solidified floating organic drop microextraction for speciation of selenium and its distribution in selenium-rich tea leaves and tea infusion by electrothermal vapourisation inductively coupled plasma mass spectrometry. Food Chem. 2015, 169, 156–161. [Google Scholar] [CrossRef]
- Warburton, E.; Goenaga-Infante, H. Methane mixed plasma—improved sensitivity of inductively coupled plasma mass spectrometry detection for selenium speciation analysis of wheat-based food. Anal. Anal. Spectrom. 2007, 22, 370–376. [Google Scholar] [CrossRef]
- Dumont, E.; Vanhaecke, F.; Cornelis, R. Selenium speciation from food source to metabolites: A critical review. Anal. Bioanal. 2006, 385, 1304–1323. [Google Scholar] [CrossRef]
- Xing, L.; Zhang, H.; Qi, R.; Tsao, R.; Mine, Y. Recent advances in the understanding of the health benefits and molecular mechanisms associated with green tea polyphenols. Food Chem. 2019, 67, 1029–1043. [Google Scholar] [CrossRef]
- Hayat, K.; Iqbal, H.; Malik, U.; Bilal, U.; Mushtaq, S. Tea and its consumption: Benefits and risks. Crit. Rev. Food Sci. 2015, 55, 939–954. [Google Scholar] [CrossRef]
- Chowdhury, P.; Barooah, A.K. Tea bioactive modulate innate immunity: In perception to COVID-19 pandemic. Front. Immunol. 2020, 11, 590716. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhou, C.; Zou, N.; Wu, Y.; Zhang, J.; An, Q.; Li, J.; Pan, C. Nano selenium foliar application enhances biosynthesis of tea leaves in metabolic cycles and associated responsive pathways. Environ. Pollut. 2021, 273, 116503. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, T.; Yang, C. Effect of foliar selenium spray on the selenium content and quality of black and green teas produced by Taicha 12. Southwest China J. Agric. Sci. 2016, 29, 2578–2582. [Google Scholar]
- Wang, Q.; Guan, X.; Lai, C.; Gao, H.; Zheng, Y.; Huang, J.; Lin, B. Selenium enrichment improves anti-proliferative effect of oolong tea extract on human hepatoma HuH-7 cells. Food Chem. Toxicol. 2021, 147, 111873. [Google Scholar] [CrossRef] [PubMed]
- Molan, A.L.; Flanagan, J.; Wei, W.; Moughan, P.J. Selenium-containing green tea has higher antioxidant and prebiotic activities than regular green tea. Food Chem. 2009, 114, 829–835. [Google Scholar] [CrossRef]
- Hodgson, J.M.; Croft, K.D. Tea flavonoids and cardiovascular health. Aspects Med. 2010, 31, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.; Cheigh, C.; Chung, M. Relationship analysis between flavonoids structure and subcritical water extraction (SWE). Food Chem. 2014, 143, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Barreca, D.; Trombetta, D.; Smeriglio, A.; Mandalari, G.; Romeo, O.; Felice, M.R.; Gattuso, G.; Nabavi, S.M. Food flavonols: Nutraceuticals with complex health benefits and functionalities. Trends Food Sci. 2021, 117, 194–204. [Google Scholar] [CrossRef]
- Fan, Z.; Jia, W.; Du, A.; Xia, Z.; Kang, J.; Xue, L.; Sun, Y. Discovery of Se-containing flavone in Se-enriched green tea and the potential application value in the immune regulation. Food Chem. 2022, 94, 3468. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yue, C.; Zhou, C.; Huang, Y.; Cao, H.; Hao, X.; Wang, X.; Zeng, J. Cloning and expression analysis of the tea tree sulfate transporter protein gene CsSUL3.5. Acta Hortic. Sinica. 2015, 42, 2306–2314. [Google Scholar]
- Wang, J.; Cappa, J.J.; Harris, J.; Edger, P.; Zhou, W.; Pires, J.; Adair, M.; Unruh, S.; Simmons, M.; Schiavon, M. Transcriptome—wide comparison of selenium hyperaccumulator and nonaccumulator Stanleya species provides new insight into key processes mediating the hyperaccumulation syndrome. Plant Biotechnol. 2018, 16, 1582–1594. [Google Scholar] [CrossRef]
- Yang, X.; Liao, X.; Yu, L. Combined metabolome and transcriptome analysis reveal the mechanism of selenate influence on the growth and quality of cabbage (Brassica oleracea var. capitata L.). Food Res. Int. 2022, 156, 111135. [Google Scholar] [CrossRef]
- Ye, Y.; Yan, J.; Cui, J.; Mao, S.; Li, M.; Liao, X.; Tong, H. Dynamic changes in amino acids, catechins, caffeine and gallic acid in green tea during withering. J. Food Compos. Anal. 2018, 66, 98–108. [Google Scholar] [CrossRef]
- Li, H.; Li, F.; Yang, F.; Fang, Y.; Xin, Z.; Zhao, L.; Hu, Q. Size effect of Se-Enriched green tea particles on in vitro antioxidant and antitumor activities. Food Chem. 2008, 56, 4529–4533. [Google Scholar] [CrossRef]
- Liu, M.; Tang, D.; Shi, Y.; Ma, L.; Li, Y.; Zhang, Q.; Ruan, J. Short-term inhibition of glutamine synthetase leads to reprogramming of amino acid and lipid metabolism in roots and leaves of tea plant (Camellia sinensis L.). BMC Plant Biol. 2019, 19, 425. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, Y.; Liu, J.; Chen, Y.; Zhang, X. Exploring the effects of selenium treatment on the nutritional quality of tomato fruit. Food Chem. 2018, 252, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Oñatibia-Astibia, A.; Martínez-Pinilla, E.; Franco, R. The potential of methylxanthine-based therapies in pediatric respiratory tract diseases. Resp. Med. 2016, 112, 1–9. [Google Scholar] [CrossRef]
- Lu, L.; Chai, L.; Wang, W.; Yuan, X.; Li, S.; Cao, C. A Selenium-Enriched ziyang green tea polysaccharide induces Bax-Dependent mitochondrial apoptosis and inhibits TGF-β1-Stimulated collagen expression in human keloid fibroblasts via NG2 inactivation. Trace Elem. Res. 2017, 176, 270–277. [Google Scholar] [CrossRef]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef]
- Tang, C.; Zheng, H.X.; Li, S. Tea aroma formation. Food Sci. Hum. Wellness 2015, 4, 9–27. [Google Scholar]
- Yang, Z.Y.; Baldermann, S.; Watanabe, N. Recent studies of the volatile compounds in tea. Food Res. 2013, 53, 585–599. [Google Scholar] [CrossRef]
- Zhu, Y.; Lv, H.; Shao, C.; Kang, S.; Zhang, Y.; Guo, L.; Dai, D.; Tan, F.; Peng, Q.; Lin, Z. Identification of key odorants responsible for chestnut-like aroma quality of green teas. Food Res. 2018, 108, 74–82. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Tea Species | Se Source | Dose | Type of Treatment | Se Content (DW) | Increased Nutrient Content | References |
|---|---|---|---|---|---|---|
| Early Spring Green Tea | selenite and selenate fertilizer | 60 mg /L | Field foliar spraying | 7.5–10.6 mg/kg | Amino acid; vitamin C FW | [20] |
| Wu Niuzao | organic Se | 100 mg/kg | Field foliar spraying | 4.72 mg/kg | Organic selenium; polyphenol; caffeine DW | [21] |
| Baiye No.1 | Nano-Se | 13.5 g/hm2 | Field foliar spraying | NA | Significant increase in chlorophyll content FW | [22] |
| Early Summer Green Tea | Se-enriched fertilizer | 100 mg/kg | Field foliar spraying | 5.895 mg/kg | Vitamin C FW; tea polyphenol DW | [23] |
| Guilv No.1 | sodium selenite | 100 mg/L | Field foliar spraying | 15.88 mg/kg | Organic selenium; Zn, K, Fe, Ca, and Mg DW | [24] |
| Qiancha 601 | sodium selenate | 0.3 mg/L | hydroponics | ≥0.25 mg/kg | Chlorophyll FW; tea polyphenol DW | [25] |
| Zhongcha 108 | Nano-Se | 10 mg/L | Field foliar spraying | 1–1.5 mg/kg | Tea polyphenol; flavonoids; caffeine DW; amino acid chlorophyll FW | [26] |
| Tea No. 12 | organic Se | 750–2100 g/hm2 | Field foliar spraying | 0.344–1.111 mg/kg | Tea polyphenol; caffeine DW | [27] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, J.; Rao, S.; Chen, Q.; Zhang, W.; Cheng, S.; Cong, X.; Zhang, Y.; Yang, X.; Xu, F. Research Progress on the Effects of Selenium on the Growth and Quality of Tea Plants. Plants 2022, 11, 2491. https://doi.org/10.3390/plants11192491
Xiang J, Rao S, Chen Q, Zhang W, Cheng S, Cong X, Zhang Y, Yang X, Xu F. Research Progress on the Effects of Selenium on the Growth and Quality of Tea Plants. Plants. 2022; 11(19):2491. https://doi.org/10.3390/plants11192491
Chicago/Turabian StyleXiang, Juan, Shen Rao, Qiangwen Chen, Weiwei Zhang, Shuiyuan Cheng, Xin Cong, Yue Zhang, Xiaoyan Yang, and Feng Xu. 2022. "Research Progress on the Effects of Selenium on the Growth and Quality of Tea Plants" Plants 11, no. 19: 2491. https://doi.org/10.3390/plants11192491