
Development of a Highly Efficient Shoot Organogenesis System for an Ornamental Aeschynanthus pulcher (Blume) G. Don Using Leaves as Explants

Abstract
:1. Introduction
2. Results
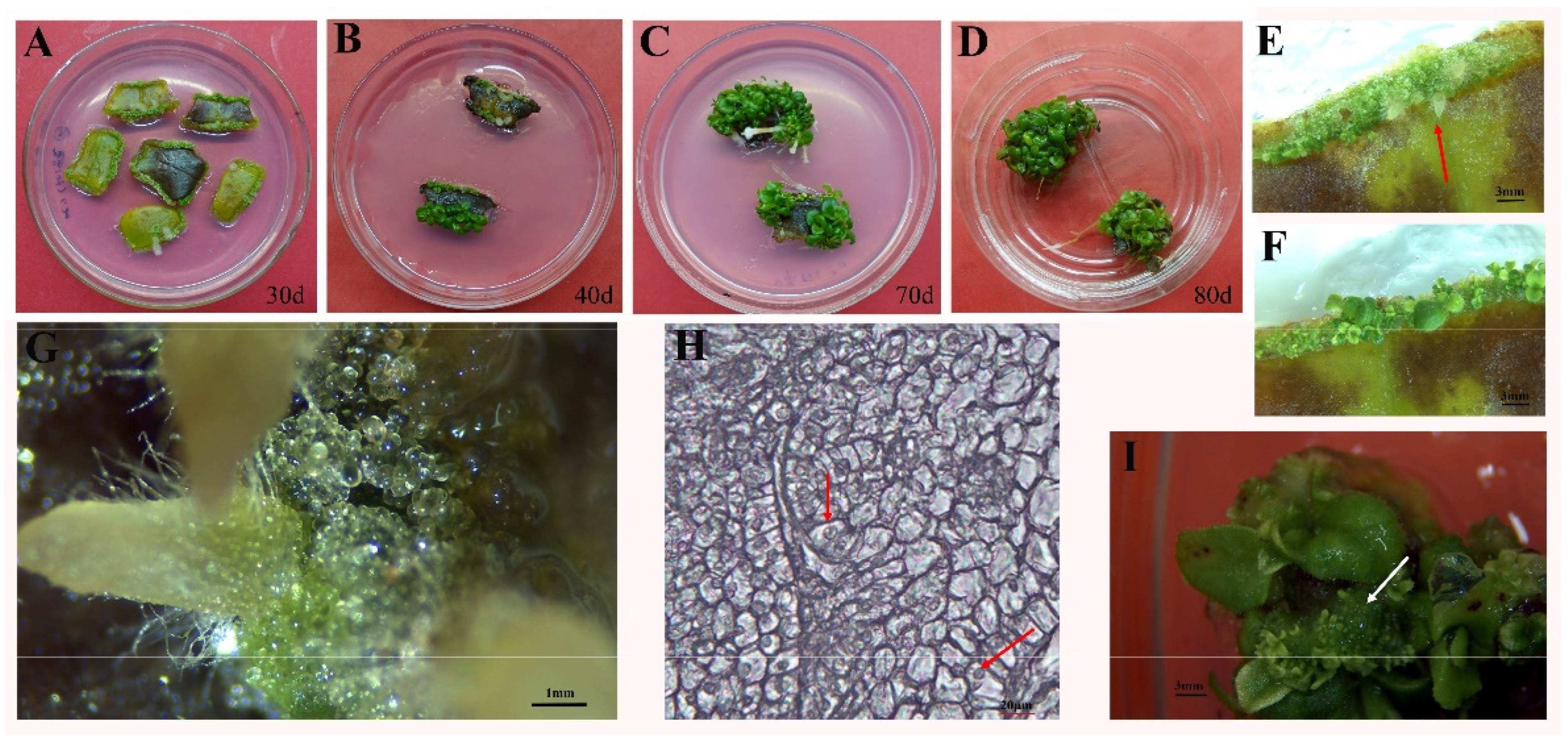
2.1. The Effect of PGR Application on the Induction of Shoot Regeneration from Leaf Explants
2.2. The Synergistic Effect of Cytokinin and Auxin on Organogenesis and Embryogenic Callus Formation from Leaf Explants
2.3. Shoot Proliferation, Rooting, and Acclimatization
3. Discussion
3.1. The Effects of Various PGRs on Organogenesis
3.2. The Synergistic Effect of Cytokinins and Auxins on Callus Formation
3.3. The Synergistic Effect of Cytokinins and Auxins on Callus Formation on Shoot Proliferation
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.2. Screening PGRs for Their Shoot Regeneration Potential from Leaf Explants
4.3. Screening PGRs for Their Embryogenic Callus Formation Potential from Leaf Explants
4.4. Shoot Proliferation, Root Formation, and Acclimatization
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Denduangboripant, J.; Cronk, Q.C. High intraindividual variation in internal transcribed spacer sequences in Aeschynanthus (Gesneriaceae): Implications for phylogenetics. Proc. R. Soc. Lond. B 2000, 267, 1407–1415. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Chen, J.; Henny, R.J. Regeneration of Aeschynanthus radicans via direct somatic embryogenesis and analysis of regenerants with flow cytometry. In Vitro Cell. Dev. Biol. Plant 2009, 45, 34–43. [Google Scholar] [CrossRef]
- Mendum, M.; Scott, S.M.; Galloway, L.E.R. The Gesneriaceae of Sulawesi IV. Two new species of Aeschynanthus. Edinb. J. Bot. 2006, 63, 67–72. [Google Scholar] [CrossRef]
- Chhetri, D.R.; Basnet, D.; Chiu, P.F.; Kalikotay, S.; Chhetrim, G.; Parajuli, S. Current status of ethnomedicinal plants in the Darjeeling Himalaya. Curr. Sci. 2005, 89, 264–268. [Google Scholar]
- Huai, H.Y.; Pei, S.J. Plants used medicinally by folk healers of the Lahu people from the autonomous county of Jinping Miao, Yao, and Dai in Southwest China. Econ. Bot. 2004, 58, 265–273. [Google Scholar] [CrossRef]
- Li, S.; Long, C.; Liu, F.; Lee, S.; Guo, Q.; Li, R.; Liu, Y. Herbs for medicinal baths among the traditional Yao communities of China. J. Ethnopharmcol. 2006, 108, 59–67. [Google Scholar] [CrossRef]
- Middleton, D.J. A Revision of Aeschynanthus (Gesneriaceae) in Thailand. Edinb. J. Bot. 2007, 64, 363–429. [Google Scholar] [CrossRef]
- Halevy, A.H. Aeschynanthus Pulcher. In CRC Handbook of Flowering; CRC Press: Boca Raton, FL, USA, 1985; p. 393. [Google Scholar]
- Huxley, A. The New Royal Horticultural Society Dictionary of Gardening; Macmillan: New York, NY, USA, 1994. [Google Scholar]
- Wu, S.T.; Chen, Y.X.; Li, L. Potted production and management technology of Aeschynanthus pulcher. J. Green Sci. Technol. 2020, 3, 110–111. (In Chinese) [Google Scholar]
- Zheng, Z.B.; Cheng, D.M.; Liu, Y.X.; Ye, Z.D.; Cheng, S.C.; Luo, S. Studies on cultivation and florescence regulation techniques of Aeschynanthus pulchira. Guangdong Agric. Sci. 2014, 41, 112–114. (In Chinese) [Google Scholar]
- Zheng, P.; Xu, H.G.; Lin, D.F.; Zheng, K. Cutting propagation and cultivation technology of Aeschynanthus radicans. J. China South. Agric. Mach. 2020, 51, 12–16. (In Chinese) [Google Scholar]
- Parrella, G.; Gognalons, P.; Gebre-Selassie, K.; Vovlas, C.; Marchoux, G. An update of the host range of tomato spotted wilt virus. J. Plant Path. 2003, 85, 227–264. [Google Scholar]
- Mei, B.J.; Ai, H. Shoot tip culture and plantlet regeneration of Aeschynanthus marmoratus. Plant Physiol. J. 1989, 6, 48. (In Chinese) [Google Scholar]
- Zhang, J.J.; Zhang, L.F.; Li, L.H.; Xu, H. Callus induction and plantlet regeneration of Aeschynanthus lobbianus. Plant Physiol. J. 2004, 2, 196. (In Chinese) [Google Scholar]
- Zeng, Q.Z.; Han, Z.Q.; Kang, X.Y. Adventitious shoot regeneration from leaf, petiole and root explants in triploid (Populus alba × P. glandulosa) × P. tomentosa. Plant Cell Tiss. Org. Cult. 2019, 138, 121–130. [Google Scholar] [CrossRef]
- Yang, H.L.; Yang, Y.H.; Wang, Q.; He, J.Y.; Liang, L.Y.; Qiu, H.; Wang, Y.; Zou, L.J. Adventitious shoot regeneration from leaf explants in Sinningia Hybrida ‘Isa’s Murmur’. Plants 2022, 11, 1232. [Google Scholar] [CrossRef]
- Kumari, P.; Singh, S.; Yadav, S.; Tran, L.S.P. Pretreatment of seeds with thidiazuron delimits its negative effects on explants and promotes regeneration in chickpea (Cicer arietinum L.). Plant Cell Tiss. Org. Cult. 2018, 133, 103–114. [Google Scholar] [CrossRef]
- Yu, R.P.; Zhang, G.F.; Li, H.; Cao, H.; Mo, X.J.; Gui, M.; Zhou, X.H.; Jiang, Y.L.; Li, S.C.; Wang, J.H. In vitro propagation of the endangered tree fern Cibotium barometz through formation of green globular bodies. Plant Cell Tiss. Org. Cult. 2017, 128, 369–379. [Google Scholar] [CrossRef]
- Wu, Q.G.; Yang, H.L.; Sun, Y.X.; Hu, J.Y.; Zou, L.J. Organogenesis and high-frequency plant regeneration in Caryopteris terniflora Maxim. using thidiazuron. In Vitro Cell. Dev. Biol. Plant 2020, 57, 39–47. [Google Scholar] [CrossRef]
- Chen, S.Y.; Xiong, Y.P.; Yu, X.C.; Pang, J.H.; Zhang, T.; Wu, K.L.; Ren, H.; Jian, S.G. Adventitious shoot organogenesis from leaf explants of Portulaca pilosa L. Sci. Rep. 2020, 10, 3675. [Google Scholar] [CrossRef]
- Wu, Q.G.; Zhang, C.; Yang, H.L.; Hu, J.Y.; Zou, L.J. In vitro propagation via organogenesis and formation of globular bodies of Salvia plebeia: A valuable medicinal plant. In Vitro Cell. Dev. Biol. Plant 2021, 58, 51–60. [Google Scholar] [CrossRef]
- Trifunovi‘c, M.; Subotić, A.; Petrić, M.; Jevremović, S. The Role of Arabinogalactan Proteins in Morphogenesis of Centaurium erythraea Rafn In Vitro; Springer: Berlin, Germany, 2015; pp. 113–138. [Google Scholar]
- Frick, E.M.; Strader, L.C. Roles for IBA-derived auxin in plant development. J. Exp. Bot. 2018, 69, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Suraya, A.A.; Misran, A.; Hakiman, M. The efficient and easy micropropagation protocol of Phyllanthus niruri. Plants 2021, 10, 2141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Qin, S.; Cao, H.; Zhang, Q.; Luo, K. Plant regeneration in Vernicia fordii from apical bud. Heliyon 2020, 6, e03884. [Google Scholar] [CrossRef] [PubMed]
- Gladfelter, H.J.; Wilde, H.D. Development of embryogenic dogwood cultures and the regeneration of plants. Plant Biotechnol. 2019, 36, 53–56. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Sugimoto, K.; Iwase, A. Plant callus: Mechanisms of induction and repression. Plant Cell 2013, 25, 3159–3173. [Google Scholar] [CrossRef]
- Chirumamilla, P.; Gopu, C.; Jogam, P.; Taduri, S. Highly efficient rapid micropropagation and assessment of genetic fidelity of regenerants by ISSR and SCoT markers of Solanum khasianum Clarke. Plant Cell Tiss. Org. Cult. 2021, 144, 397–407. [Google Scholar] [CrossRef]
- Kumar, P.P.; Chandler, S.F.; Harry, I.S.; Lu, C.Y.; Stasolla, C.; Yeung, E.C. Trevor alleyne thorpe: His academic life and scientific legacy. In Vitro Cell. Dev. Biol. Plant 2020, 56, 728–737. [Google Scholar] [CrossRef]
- Shukla, P.S.; Das, A.K.; Jha, B.; Agarwal, P.K. High-frequency in vitro shoot regeneration in Cucumis sativus by inhibition of endogenous auxin. In Vitro Cell. Dev. Biol. Plant 2014, 50, 729–737. [Google Scholar] [CrossRef]
- Uno, Y.; Yagi, M. Anther-Based Regeneration of African Violet (Saintpaulia ionantha). Methods Mol. Biol. 2021, 2289, 249–261. [Google Scholar]
- Li, Q.; Deng, M.; Zhang, J.; Zhao, W.; Song, Y.; Li, Q.; Huang, Q. Shoot organogenesis and plant regeneration from leaf explants of Lysionotus serratus D. Don. Sci. World J. 2013, 5, 280384. [Google Scholar]
- Li, J.; Zhang, D.; Ouyang, K.; Chen, X. High frequency plant regeneration from leaf culture of Neolamarckia cadamba. Plant Biotechnol. 2019, 36, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bozorov, T.A.; Li, D.X.; Zhou, P.; Li, D.X.; Wen, X.J.; Ding, Y.; Zhang, D.Y. An efficient in vitro regeneration system from different wild apple (Malus sieversii) explants. Plant Methods 2020, 16, 56. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; An, D.J.; Moon, K.B.; Cho, H.S.; Min, S.R.; Sohn, J.H.; Jeon, J.H.; Kim, H.S. Highly efficient plant regeneration and Agrobacterium–mediated transformation of Helianthus tuberosus L. Ind. Crops Prod. 2016, 83, 670–679. [Google Scholar] [CrossRef]
- Zhang, J.J.; Yang, Y.S.; Lin, M.F.; Li, S.Q.; Tang, Y.; Chen, H.B.; Chen, X.Y. An efficient micropropagation protocol for direct organogenesis from leaf explants of an economically valuable plant, drumstick (Moringa oleifera Lam.). Ind. Crops Prod. 2017, 103, 59–63. [Google Scholar]
- Xu, L.; Cheng, F.; Zhong, Y. Efficient plant regeneration via meristematic nodule culture in Paeonia ostii ‘Feng Dan’. Plant Cell Tiss. Org. Cult. 2022, 149, 599–608. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Zhu, S.S.; Gao, X.Q.; Zhang, X.S. Cytokinin and auxin regulates WUS induction and inflorescence regeneration in vitro in Arabidopsis. Plant Cell Rep. 2010, 29, 927–933. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Zhou, J.H.; Wu, T.; Cao, J.S. Shoot regeneration and the relationship between organogenic capacity and endogenous hormonal contents in pumpkin. Plant Cell Tiss. Org. Cult. 2008, 93, 323–331. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassay with tobacco tissue culture. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Medium (MS) | Browned (%) | Response (%) | Rooting (%) | Shoots/Explant | Visible Appearance |
|---|---|---|---|---|---|
| BA 3 mg L−1 + PVP 1 g L−1 | 54.3 ± 1.7 b | 50.5 ± 1.9 c | 0.0 ± 0.0 b | 3.5 ± 0.4 c | No callus, direct induction of shoots regeneration |
| TDZ 3 mg L−1 + PVP 1 g L−1 | 21.5c ± 1.1c | 82.4 ± 2.5 a | 0.0 ± 0.0 b | 38.6 ± 3.5 a | Callus around the leaves, abundant adventitious shoots |
| ZT 3 mg L−1 + PVP 1 g L−1 | 100.0 ± 0.0 a | 0.0 ± 0.0 d | 0.0 ± 0.0 b | 0.0 ± 0.0 d | No callus, no shoots, leaves brown and withered |
| IBA 3 mg L−1 + PVP 1 g L−1 | 40.6 ± 2.0 b | 60.1b ± 2.4 b | 95.4 ± 3.2 a | 14.5 ± 2.4 b | No callus, leaves first differentiate into adventitious root then adventitious shoots |
| 2,4-D 3 mg L−1 + PVP 1 g L−1 | 100.0 ± 0.0 a | 0.0 ± 0.0 d | 0.0 ± 0.0 b | 0.0 ± 0.0 d | No callus, no shoots, leaves brown and withered |
| PIC 3 mg L−1 + PVP 1 g L−1 | 0.0 ± 0.0 d | 0.0 ± 0.0 d | 0.0 ± 0.0 b | 0.0 ± 0.0 d | No callus, no shoots, leaves fade to green |
| Treatment | Callogenesis (%) | Shoot Proliferation Coefficient | Root Number | Observed Results |
|---|---|---|---|---|
| NAA 0.1 mg L−1 + BA 4 mg L−1 + Casein 5 mg L−1 + PVP 1 g L−1 | 72.6 ± 2.8 c | 10.0 ± 1.5 b | 12.3 ± 1.0 b | Embryogenic callus; adventitious shoot differentiation; adventitious root formation, dwarf shoot |
| NAA 0.1 mg L−1 + BA 3 mg L−1 + Casein 5 mg L−1 + PVP 1 g L−1 | 95.2 ± 2.4 a | 19.4 ± 2.2 a | 18.8 ± 0.9 a | Same result as above |
| NAA 0.1 mg L−1 + BA 2 mg L−1 + Casein 5 mg L−1 + PVP 1 g L−1 | 80.3 ± 4.6 b | 13.6 ± 1.8 b | 14.2 ± 2.7 a | Same result as above |
| NAA 0.1 mg L−1 + TDZ 4 mg L−1 + Casein 5 mg L−1 + PVP 1 g L−1 | 82.4 ± 3.1 b | 11.6 ± 1.3 b | 9.2 ± 1.4 b | Same result as above |
| NAA 0.1 mg L−1 + TDZ 3 mg L−1 + Casein 5 mg L−1 + PVP 1 g L−1 | 96.6 ± 1.8 a | 21.5 ± 3.7 a | 14.4 ± 0.8 a | Same result as above |
| NAA 0.1 mg L−1 + TDZ 2 mg L−1 + Casein 5 mg L−1 + PVP 1 g L−1 | 84.0 ± 1.7 b | 15.6 ± 1.3 b | 10.6 ± 0.7 b | Embryogenic callus; adventitious shoot differentiation; adventitious root formation; robust shoot |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Yuan, H.; Du, C.; Liang, L.; Chen, M.; Zou, L. Development of a Highly Efficient Shoot Organogenesis System for an Ornamental Aeschynanthus pulcher (Blume) G. Don Using Leaves as Explants. Plants 2022, 11, 2456. https://doi.org/10.3390/plants11192456
Yang H, Yuan H, Du C, Liang L, Chen M, Zou L. Development of a Highly Efficient Shoot Organogenesis System for an Ornamental Aeschynanthus pulcher (Blume) G. Don Using Leaves as Explants. Plants. 2022; 11(19):2456. https://doi.org/10.3390/plants11192456
Chicago/Turabian StyleYang, Honglin, Honglin Yuan, Cunmei Du, Liyun Liang, Meiling Chen, and Lijuan Zou. 2022. "Development of a Highly Efficient Shoot Organogenesis System for an Ornamental Aeschynanthus pulcher (Blume) G. Don Using Leaves as Explants" Plants 11, no. 19: 2456. https://doi.org/10.3390/plants11192456