
Protein Hydrolysates from Animal or Vegetal Sources Affect Morpho-Physiological Traits, Ornamental Quality, Mineral Composition, and Shelf-Life of Chrysanthemum in a Distinctive Manner

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Results
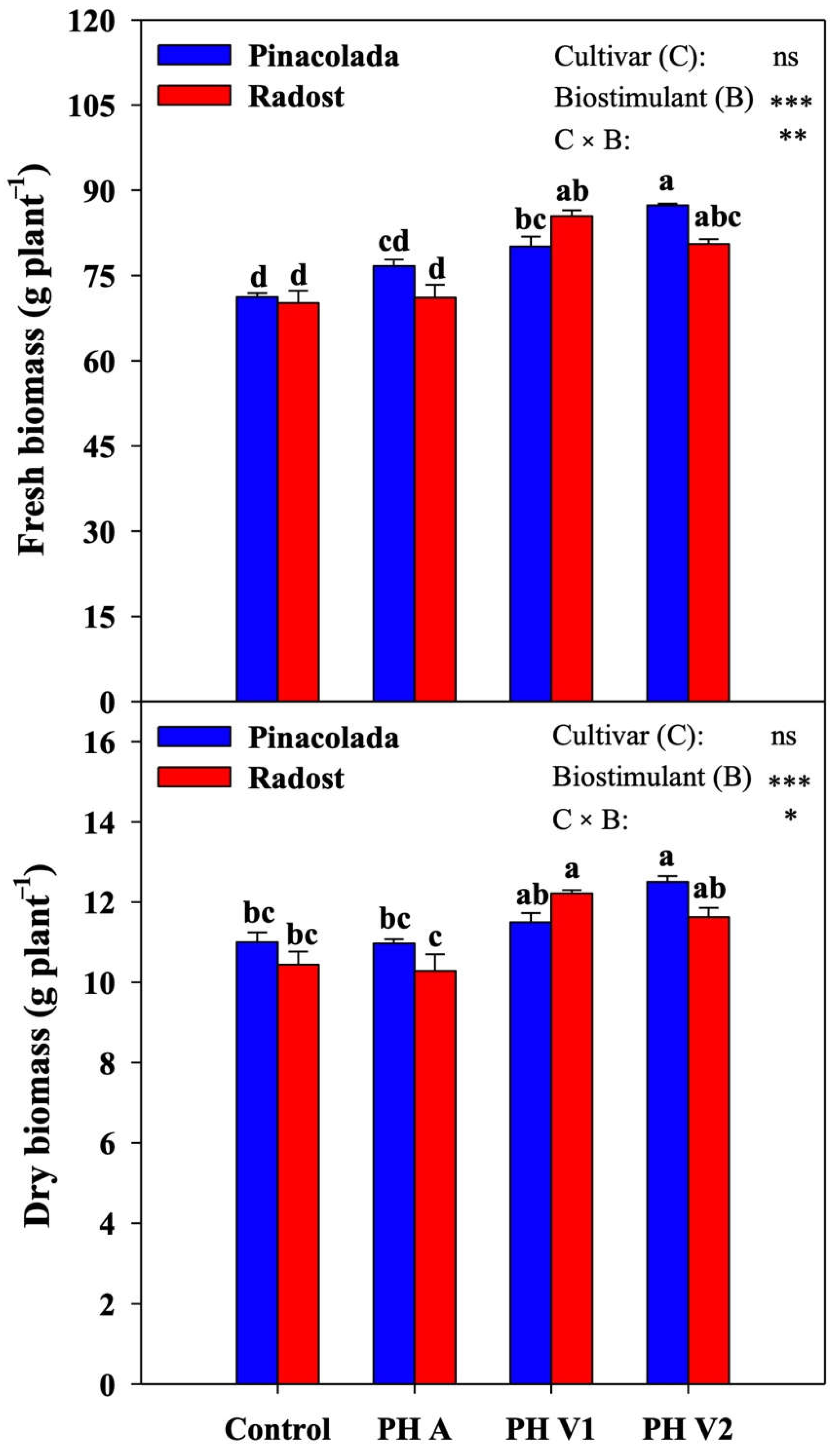
2.1. Biomass Production and Quality Parameters
2.2. Physiological Measurements
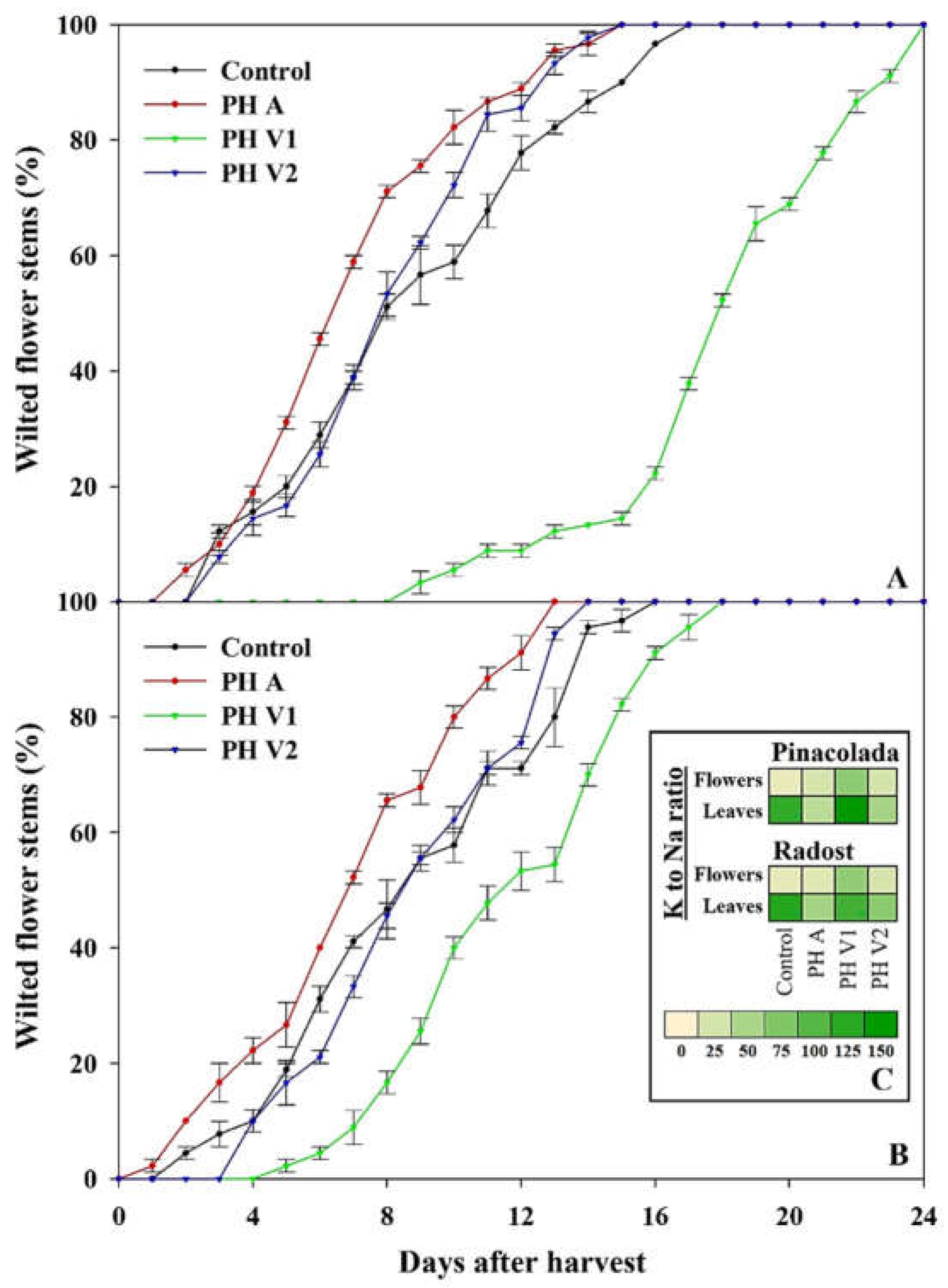
2.3. Mineral Composition
2.4. Vase Life of Flower Stems
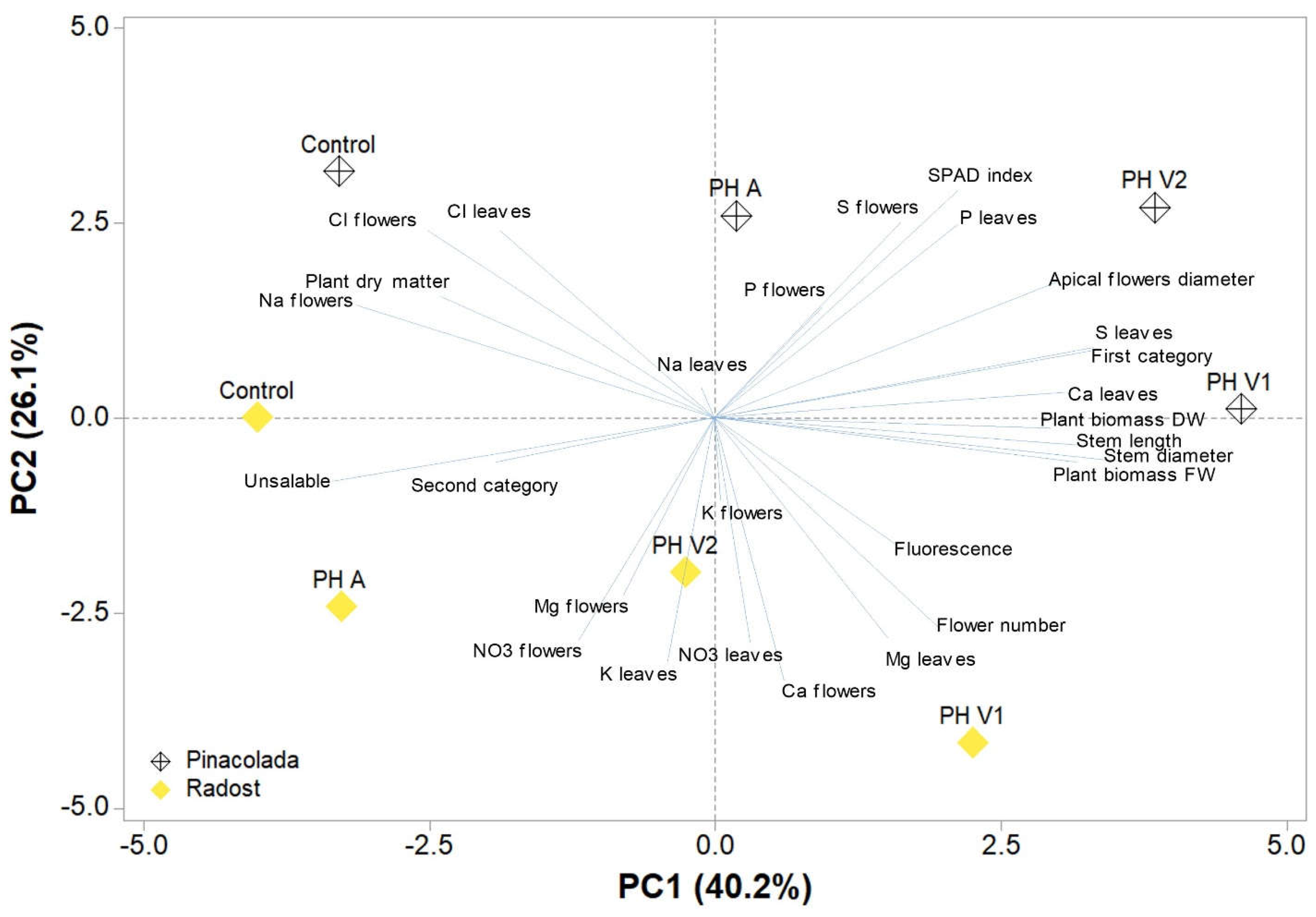
2.5. Principal Component Analysis (PCA)
3. Discussion
4. Materials and Methods
4.1. Experimental Site, Plant Material, and Growth Conditions
4.2. Biostimulant Treatments and Experimental Design
4.3. Biomass Production and Quality Parameters
4.4. Soil Plant Analysis Development (SPAD) Index and Maximum Quantum Efficiency of Photosystem II Measurements (Fv/Fm)
4.5. Mineral Content Analysis
4.6. Post-Harvest Measurements
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gabellini, S.; Scaramuzzi, S. Evolving Consumption Trends, Marketing Strategies, and Governance Settings in Ornamental Horticulture: A Grey Literature Review. Horticulturae 2022, 8, 234. [Google Scholar] [CrossRef]
- Sun, C.-Q.; Huang, Z.-Z.; Wang, Y.-L.; Chen, F.-D.; Teng, N.-J.; Fang, W.-M.; Liu, Z.-L. Overcoming pre-fertilization barriers in the wide cross between Chrysanthemumgrandiflorum (Ramat.) Kitamura and C. nankingense (Nakai) Tzvel. by using special pollination techniques. Euphytica 2011, 178, 195–202. [Google Scholar] [CrossRef]
- Amarin, R.; Kafawin, O.; Al-Zyoud, F.; Haddad, N.; Ayad, J.; Amarin, A. Performance of chrysanthemum Chrysanthemum morifolium Ramat. Jordan J. Agric. Sci. 2021, 17, 15. [Google Scholar]
- CBI. Exporting Chrysanthemums to Europe. A Study of CBI Ministry of Foreign Affairs. Available online: https://www.cbi.eu/market-information/cut-flowers-foliage/chrysanthemums/europe (accessed on 5 August 2022).
- Wainwright, H.; Jordan, C.; Day, H. Environmental Impact of Production Horticulture. In Horticulture: Plants for People and Places; Springer: Dordrecht, The Netherlands, 2014; Volume 1, pp. 503–522. [Google Scholar]
- Moreno, C.A.P.; Hernández, R.E.R.; Arredondo, H.I.V.; Castro, S.H.L.; Hasenstab, C. An Environmental Evaluation of the Cut-Flower Supply Chain (Dendranthema Grandiflora) Through a Life Cycle Assessment. Reveia 2019, 16, 27–42. [Google Scholar] [CrossRef]
- Cavalcante da Costa, L.; Ferreira de Araujo, F.; Souto Ribeiro, W.; de Sousa Santos, M.N.; Finger, F.L. Postharvest physiology of cut flowers. Ornam. Hortic. 2021, 27, 374–385. [Google Scholar] [CrossRef]
- Nguyen, T.K.; Lim, J.H. Do Eco-Friendly Floral Preservative Solutions Prolong Vase Life Better than Chemical Solutions? Horticulturae 2021, 7, 415. [Google Scholar] [CrossRef]
- Toumi, K.; Joly, L.; Vleminckx, C.; Schiffers, B. Risk Assessment of Florists Exposed to Pesticide Residues through Handling of Flowers and Preparing Bouquets. Int. J. Environ. Res. Public Health 2017, 14, 526. [Google Scholar] [CrossRef]
- Cristiano, G.; De Lucia, B. Petunia Performance Under Application of Animal-Based Protein Hydrolysates: Effects on Visual Quality, Biomass, Nutrient Content, Root Morphology, and Gas Exchange. Front. Plant Sci. 2021, 12, 890. [Google Scholar] [CrossRef]
- Cristiano, G.; Pallozzi, E.; Conversa, G.; Tufarelli, V.; De Lucia, B. Effects of an Animal-Derived Biostimulant on the Growth and Physiological Parameters of Potted Snapdragon (Antirrhinum majus L.). Front. Plant Sci. 2018, 9, 861. [Google Scholar] [CrossRef]
- Kim, H.-J.; Ku, K.-M.; Choi, S.; Cardarelli, M. Vegetal-Derived Biostimulant Enhances Adventitious Rooting in Cuttings of Basil, Tomato, and Chrysanthemum via Brassinosteroid-Mediated Processes. Agronomy 2019, 9, 74. [Google Scholar] [CrossRef]
- Toscano, S.; Romano, D.; Massa, D.; Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulant applications in low input horticultural cultivation systems. Italus Hortus 2018, 25, 27–36. [Google Scholar] [CrossRef]
- Benedetto, A.D.; Pittaluga, D.; Benedicto, D. Protein hydrolysate formulations on vase life for cut roses. Científica 2006, 34, 269–272. [Google Scholar]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- da Silva, C.P.; Laschi, D.; Ono, E.O.; Rodrigues, J.D.; Mogor, Á.F. Aplicação Foliar do Extrato de Alga Ascophyllum nodosum e do Ácido Glutâmico no Desenvolvimento Inicial de Crisântemos (Dendranthema morifolium Ramat. Kitam.) em Vasos. Rev. Bras. Hortic. Ornam. 2010, 16, 179–181. [Google Scholar]
- Taranjit, S.; Madhu, B. Effect of putrescine and benzyl adenine on growth, flowering and post-harvest keeping quality parameters in chrysanthemum (Chrysanthemum morifolium Ramat.). J. Hortic. Sci. 2020, 15, 191–196. [Google Scholar] [CrossRef]
- Sedaghathoor, S.; Narouei, Z.; Sajjadi, S.A.; Piri, S. The effect of chemical treatments (silver thiosulfate and putrescine) on vase life and quality of cut Chrysanthemum morifolium (Ram.) flowers. Cogent Biol. 2020, 6, 1754320. [Google Scholar] [CrossRef]
- Wise, K.; Gill, H.; Selby-Pham, J. Willow bark extract and the biostimulant complex Root Nectar® increase propagation efficiency in chrysanthemum and lavender cuttings. Sci. Hortic. 2020, 263, 109108. [Google Scholar] [CrossRef]
- Gawade, N.; Varu, D.; Devdhara, U. Effect of biostimulants and biofertilizers on growth and flowering of Chrysanthemum cv. Ratlam selection. Int. J. Chem. Stud. 2019, 7, 3423–3428. [Google Scholar]
- Fan, H.-M.; Wang, X.-E.; Sun, X.; Li, Y.-Y.; Sun, X.-Z.; Zheng, C.-S. Effects of humic acid derived from sediments on growth, photosynthesis and chloroplast ultrastructure in chrysanthemum. Sci. Hortic. 2014, 177, 118–123. [Google Scholar] [CrossRef]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtílek, M.; Panzarová, K.; et al. Understanding the Biostimulant Action of Vegetal-Derived Protein Hydrolysates by High-Throughput Plant Phenotyping and Metabolomics: A Case Study on Tomato. Front. Plant Sci. 2019, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef] [PubMed]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Ertani, A.; Pizzeghello, D.; Francioso, O.; Sambo, P.; Sanchez-Cortes, S.; Nardi, S. Capsicum chinensis L. growth and nutraceutical properties are enhanced by biostimulants in a long-term period: Chemical and metabolomic approaches. Front. Plant Sci. 2014, 5, 375. [Google Scholar] [CrossRef]
- Ertani, A.; Cavani, L.; Pizzeghello, D.; Brandellero, E.; Altissimo, A.; Ciavatta, C.; Nardi, S. Biostimulant activity of two protein hydrolyzates in the growth and nitrogen metabolism of maize seedlings. J. Plant Nutr. Soil Sci. 2009, 172, 237–244. [Google Scholar] [CrossRef]
- Ertani, A.; Schiavon, M.; Muscolo, A.; Nardi, S. Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays L. plants. Plant Soil 2013, 364, 145–158. [Google Scholar] [CrossRef]
- Liu, X.-Q.; Lee, K.-S. Effect of Mixed Amino Acids on Crop Growth, Agricultural Science; Aflakpui, G., Ed.; InTech Europe: Rijeka, Croatia, 2012; Available online: http://www.intechopen.com/books/agricultural-science/effect-of-mixed-amino-acids-on-crop-growth (accessed on 27 August 2022).
- Carillo, P.; Colla, G.; Fusco, G.M.; Dell’Aversana, E.; El-Nakhel, C.; Giordano, M.; Pannico, A.; Cozzolino, E.; Mori, M.; Reynaud, H.; et al. Morphological and Physiological Responses Induced by Protein Hydrolysate-Based Biostimulant and Nitrogen Rates in Greenhouse Spinach. Agronomy 2019, 9, 450. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant Sci. 2014, 5, 448. [Google Scholar] [CrossRef]
- Colla, G.; Cardarelli, M.; Bonini, P.; Rouphael, Y. Foliar Applications of Protein Hydrolysate, Plant and Seaweed Extracts Increase Yield but Differentially Modulate Fruit Quality of Greenhouse Tomato. HortScience 2017, 52, 1214–1220. [Google Scholar] [CrossRef]
- Rouphael, Y.; Giordano, M.; Cardarelli, M.; Cozzolino, E.; Mori, M.; Kyriacou, M.C.; Bonini, P.; Colla, G. Plant- and Seaweed-Based Extracts Increase Yield but Differentially Modulate Nutritional Quality of Greenhouse Spinach through Biostimulant Action. Agronomy 2018, 8, 126. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dosedependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Baffi, C.; Colla, G. A Vegetal Biopolymer-Based Biostimulant Promoted Root Growth in Melon While Triggering Brassinosteroids and Stress-Related Compounds. Front. Plant Sci. 2018, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- Lucini, L.; Rouphael, Y.; Cardarelli, M.; Canaguier, R.; Kumar, P.; Colla, G. The effect of a plant-derived biostimulant on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 2015, 182, 124–133. [Google Scholar] [CrossRef]
- Li, X.; Zeng, R.; Liao, H. Improving crop nutrient efficiency through root architecture modifications. J. Integr. Plant Biol. 2016, 58, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Cerdán, M.; Sánchez-Sánchez, A.; Oliver, M.; Juárez, M.; Sánchez-Andreu, J.J. Effect of foliar and root applications of amino acids on iron uptake by tomato plants. Acta Hortic. 2009, 830, 481–488. [Google Scholar]
- Cerdán, M.; Sánchez-Sánchez, A.; Jordá, J.D.; Juárez, M.; Sánchez-Andreu, J. Effect of commercial amino acids on iron nutrition of tomato plants grown under lime-induced iron deficiency. J. Plant Nutr. Soil Sci. 2013, 176, 859–866. [Google Scholar] [CrossRef]
- Rouphael, Y.; Carillo, P.; Cristofano, F.; Cardarelli, M.; Colla, G. Effects of vegetal- versus animal-derived protein hydrolysate on sweet basil morpho-physiological and metabolic traits. Sci. Hortic. 2021, 284, 110123. [Google Scholar] [CrossRef]
- Darras, A. Overview of the Dynamic Role of Specialty Cut Flowers in the International Cut Flower Market. Horticulturae 2021, 7, 51. [Google Scholar] [CrossRef]
- Hasler, K. Environmental impact of mineral fertilizers: Possible Improvements through the Adoption of Eco-Innovations. Ph.D. Thesis, Wageningen University and Research, Wageningen, The Netherlands, 2017. [Google Scholar]
- Statista. Revenue of Cut Flowers Sold by Royal FloraHolland in 2020, by Type (in million Euros). Available online: https://www.statista.com/statistics/829413/sales-value-of-cut-flowers-sold-by-royal-floraholland-by-type/ (accessed on 7 August 2022).
- Carillo, P.; De Micco, V.; Ciriello, M.; Formisano, L.; El-Nakhel, C.; Giordano, M.; Colla, G.; Rouphael, Y. Morpho-Anatomical, Physiological, and Mineral Composition Responses Induced by a Vegetal-Based Biostimulant at Three Rates of Foliar Application in Greenhouse Lettuce. Plants 2022, 11, 2030. [Google Scholar] [CrossRef]
- Carillo, P.; Woo, S.L.; Comite, E.; El-Nakhel, C.; Rouphael, Y.; Fusco, G.M.; Borzacchiello, A.; Lanzuise, S.; Vinale, F. Application of Trichoderma harzianum, 6-Pentyl-α-pyrone and Plant Biopolymer Formulations Modulate Plant Metabolism and Fruit Quality of Plum Tomatoes. Plants 2020, 9, 771. [Google Scholar] [CrossRef] [PubMed]
- Carillo, P.; Colla, G.; El-Nakhel, C.; Bonini, P.; D’Amelia, L.; Dell’Aversana, E.; Pannico, A.; Giordano, M.; Sifola, M.I.; Kyriacou, M.C.; et al. Biostimulant Application with a Tropical Plant Extract Enhances Corchorus olitorius Adaptation to Sub-Optimal Nutrient Regimens by Improving Physiological Parameters. Agronomy 2019, 9, 249. [Google Scholar] [CrossRef]
- Niyokuri, A.; Nyalala, S.; Mwangi, M. Effects of bioslurry and plant biostimulant Hicure® on yield, flower quality and vase life of carnation (Dianthus caryophyllus L.). J. Appl. Hortic. 2017, 19, 29–34. [Google Scholar] [CrossRef]
- Niyokuri, A.; Nyalala, S.; Mwangi, M. Residual effects of bioslurry and amino acids plant biostimulant on carnation (Dianthus caryophyllus L.) flower quality. Adv. Hortic. Sci. 2018, 32, 137–142. [Google Scholar] [CrossRef]
- Han, R.; Khalid, M.; Juan, J.; Huang, D. Exogenous glycine inhibits root elongation and reduces nitrate-N uptake in pak choi (Brassica campestris ssp. Chinensis L.). PLoS ONE 2018, 13, e0204488. [Google Scholar] [CrossRef] [Green Version]
- Sorrentino, M.; Panzarová, K.; Spyroglou, I.; Spíchal, L.; Buffagni, V.; Ganugi, P.; Rouphael, Y.; Colla, G.; Lucini, L.; De Diego, N. Integration of Phenomics and Metabolomics Datasets Reveals Different Mode of Action of Biostimulants Based on Protein Hydrolysates in Lactuca sativa L. and Solanum lycopersicum L. Under Salinity. Front. Plant Sci. 2022, 12, 808711. [Google Scholar] [CrossRef]
- Cirillo, C.; De Micco, V.; Arena, C.; Carillo, P.; Pannico, A.; De Pascale, S.; Rouphael, Y. Biochemical, Physiological and Anatomical Mechanisms of Adaptation of Callistemon citrinus and Viburnum lucidum to NaCl and CaCl2 Salinization. Front. Plant Sci. 2019, 10, 742. [Google Scholar] [CrossRef]
- Carillo, P.; Cirillo, C.; De Micco, V.; Arena, C.; De Pascale, S.; Rouphael, Y. Morpho-anatomical, physiological and biochemical adaptive responses to saline water of Bougainvillea spectabilis Willd. trained to different canopy shapes. Agric. Water Manag. 2019, 212, 12–22. [Google Scholar] [CrossRef]
- Giordano, M.; El-Nakhel, C.; Carillo, P.; Colla, G.; Graziani, G.; Di Mola, I.; Mori, M.; Kyriacou, M.C.; Rouphael, Y.; Soteriou, G.A.; et al. Plant-Derived Biostimulants Differentially Modulate Primary and Secondary Metabolites and Improve the Yield Potential of Red and Green Lettuce Cultivars. Agronomy 2022, 12, 1361. [Google Scholar] [CrossRef]
- Druege, U. Influence of pre-harvest nitrogen supply on post-harvest behaviour of ornamentals: Importance of carbohydrate status, photosynthesis and plant hormones. Gartenbauwissenschaft 2000, 65, 53–64. [Google Scholar]
- Khan, M.I.; Trivellini, A.; Fatma, M.; Masood, A.; Francini, A.; Iqbal, N.; Ferrante, A.; Khan, N.A. Role of ethylene in responses of plants to nitrogen availability. Front. Plant. Sci. 2015, 6, 927. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene Role in Plant Growth, Development and Senescence: Interaction with Other Phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef]
- Fu, Y.; Yang, Y.; Chen, S.; Ning, N.; Hu, H. Arabidopsis IAR4 Modulates Primary Root Growth Under Salt Stress Through ROS-Mediated Modulation of Auxin Distribution. Front. Plant Sci. 2019, 10, 522. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Blumwald, E. Na+ transport in plants. FEBS Lett. 2007, 581, 2247–2254. [Google Scholar] [CrossRef] [PubMed]
- Trivellini, A.; Cocetta, G.; Hunter, D.A.; Vernieri, P.; Ferrante, A. Spatial and temporal transcriptome changes occurring during flower opening and senescence of the ephemeral hibiscus flower, Hibiscus rosa-sinensis. J. Exp. Bot. 2016, 67, 5919–5931. [Google Scholar] [CrossRef]
- Sorrentino, M.; De Diego, N.; Ugena, L.; Spíchal, L.; Lucini, L.; Miras-Moreno, B.; Zhang, L.; Rouphael, Y.; Colla, G.; Panzarová, K. Seed Priming With Protein Hydrolysates Improves Arabidopsis Growth and Stress Tolerance to Abiotic Stresses. Front. Plant Sci. 2021, 12, 837. [Google Scholar] [CrossRef]
- Pannico, A.; El-Nakhel, C.; Kyriacou, M.C.; Giordano, M.; Stazi, S.R.; De Pascale, S.; Rouphael, Y. Combating Micronutrient Deficiency and Enhancing Food Functional Quality Through Selenium Fortification of Select Lettuce Genotypes Grown in a Closed Soilless System. Front. Plant Sci. 2019, 10, 1495. [Google Scholar] [CrossRef]
- Dell’Aversana, E.; Hessini, K.; Ferchichi, S.; Fusco, G.M.; Woodrow, P.; Ciarmiello, L.F.; Abdelly, C.; Carillo, P. Salinity Duration Differently Modulates Physiological Parameters and Metabolites Profile in Roots of Two Contrasting Barley Genotypes. Plants 2021, 10, 307. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source of Variance | Flower Number | Stem Diameter | Stem Length | Apical Flowers Diameter | Plant Dry Matter |
|---|---|---|---|---|---|
| (no. plant−1) | (cm) | (cm) | (cm) | (%) | |
| Cultivar (C) | |||||
| Pinacolada | 10.52 ± 0.32 | 0.64 ± 0.02 | 92.90 ± 0.88 | 6.73 ± 0.19 | 14.61 ± 0.16 |
| Radost | 11.46 ± 0.48 | 0.61 ± 0.01 | 91.36 ± 0.91 | 5.57 ± 0.16 | 14.52 ± 0.08 |
| t−test | * | ns | ** | *** | ns |
| Biostimulant (B) | |||||
| Control | 9.15 ± 0.19 b | 0.57 ± 0.01 c | 90.42 ± 0.14 c | 5.78 ± 0.11 b | 15.16 ± 0.15 a |
| PH A | 11.00 ± 0.59 a | 0.61 ± 0.01 b | 89.26 ± 0.87 bc | 5.68 ± 0.44 b | 14.38 ± 0.10 b |
| PH V1 | 12.00 ± 0.40 a | 0.66 ± 0.01 a | 95.71 ± 1.04 a | 6.76 ± 0.30 a | 14.33 ± 0.08 b |
| PH V2 | 11.80 ± 0.26 a | 0.65 ± 0.02 ab | 93.14 ± 0.79 ab | 6.37 ± 0.30 a | 14.37 ± 0.12 b |
| *** | *** | *** | *** | *** | |
| C × B | |||||
| Pinacolada × Control | 9.07 ± 0.19 | 0.56 ± 0.03 | 90.43 ± 0.23 | 5.90 ± 0.07 bc | 15.45 ± 0.18 |
| Pinacolada × PH A | 10.13 ± 0.19 | 0.62 ± 0.02 | 90.96 ± 0.69 | 6.60 ± 0.32 ab | 14.31 ± 0.17 |
| Pinacolada × PH V1 | 11.33 ± 0.37 | 0.68 ± 0.01 | 96.42 ± 1.60 | 7.41 ± 0.14 a | 14.35 ± 0.09 |
| Pinacolada × PH V2 | 11.53 ± 0.30 | 0.68 ± 0.02 | 93.80 ± 1.50 | 7.00 ± 0.17 a | 14.32 ± 0.21 |
| Radost × Control | 9.23 ± 0.37 | 0.57 ± 0.01 | 90.41 ± 0.23 | 5.67 ± 0.22 c | 14.88 ± 0.01 |
| Radost × PH A | 11.87 ± 0.98 | 0.61 ± 0.01 | 87.56 ± 0.64 | 4.76 ± 0.11 d | 14.45 ± 0.13 |
| Radost × PH V1 | 12.67 ± 0.47 | 0.65 ± 0.02 | 95.00 ± 1.52 | 6.10 ± 0.09 bc | 14.30 ± 0.14 |
| Radost × PH V2 | 12.07 ± 0.44 | 0.62 ± 0.01 | 92.47 ± 0.68 | 5.73 ± 0.08 c | 14.43 ± 0.16 |
| ns | ns | ns | ** | ns |
| Source of Variance | First Category | Second Category | Unmarketable |
|---|---|---|---|
| (%) | (%) | (%) | |
| Cultivar © | |||
| Pinacolada | 72.76 ± 3.34 | 18.83 ± 1.79 | 8.42 ± 2.01 |
| Radost | 60.84 ± 2.58 | 19.86 ± 1.40 | 19.30 ± 2.07 |
| t-test | *** | ns | *** |
| Biostimulant (B) | |||
| Control | 54.55 ± 1.58 d | 21.58 ± 1.30 ab | 23.87 ± 2.45 a |
| PH A | 61.58 ± 4.22 c | 24.11 ± 1.23 a | 14.30 ± 3.26 b |
| PH V1 | 73.16 ± 1.79 b | 20.18 ± 0.74 b | 6.66 ± 1.68 d |
| PH V2 | 77.89 ± 3.89 a | 11.50 ± 1.10 c | 10.61 ± 2.88 c |
| *** | *** | *** | |
| C × B | |||
| Pinacolada × Control | 56.64 ± 2.67 d | 24.36 ± 0.82 ab | 19.00 ± 2.22 |
| Pinacolada × PH A | 70.99 ± 0.45 bc | 21.72 ± 1.10 bc | 7.29 ± 1.49 |
| Pinacolada × PH V1 | 76.90 ± 1.32 b | 20.09 ± 1.20 c | 3.01 ± 0.12 |
| Pinacolada × PH V2 | 86.51 ± 1.11 a | 9.13 ± 0.37 e | 4.36 ± 1.48 |
| Radost × Control | 52.47 ± 0.97 d | 18.81 ± 0.31 c | 28.73 ± 1.16 |
| Radost × PH A | 52.18 ± 0.52 d | 26.51 ± 0.83 a | 21.31 ± 1.33 |
| Radost × PH V1 | 69.43 ± 0.63 c | 20.27 ± 1.13 bc | 10.31 ± 0.89 |
| Radost × PH V2 | 69.27 ± 0.43 c | 13.87 ± 0.55 d | 16.87 ± 0.37 |
| *** | *** | ns |
| Source of Variance | SPAD Index | Chlorophyll Fluorescence |
|---|---|---|
| (Fv/Fm) | ||
| Cultivar (C) | ||
| Pinacolada | 54.99 ± 0.47 | 0.825 ± 0.002 |
| Radost | 43.00 ± 0.33 | 0.828 ± 0.003 |
| t-test | *** | ns |
| Biostimulant (B) | ||
| Control | 47.63 ± 2.48 b | 0.822 ± 0.003 b |
| PH A | 48.18 ± 2.79 b | 0.826 ± 0.004 ab |
| PH V1 | 50.39 ± 2.62 a | 0.834 ± 0.002 a |
| PH V2 | 49.80 ± 2.89 a | 0.824 ± 0.004 b |
| *** | * | |
| C × B | ||
| Pinacolada × Control | 53.09 ± 0.86 b | 0.817 ± 0.001 bc |
| Pinacolada × PH A | 54.41 ± 0.40 ab | 0.820 ± 0.004 bc |
| Pinacolada × PH V1 | 56.24 ± 0.42 a | 0.831 ± 0.003 abc |
| Pinacolada × PH V2 | 56.24 ± 0.51 a | 0.832 ± 0.002 ab |
| Radost × Control | 42.16 ± 0.07 d | 0.827 ± 0.003 abc |
| Radost × PH A | 41.95 ± 0.19 d | 0.831 ± 0.004 abc |
| Radost × PH V1 | 44.55 ± 0.28 c | 0.836 ± 0.003 a |
| Radost × PH V2 | 43.36 ± 0.35 cd | 0.816 ± 0.003 c |
| ns | ** |
| Source of Variance | Nitrate | P | K | Ca | Mg | S | Na | Cl | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | (g kg−1 DW) | |||||||||
| Leaves | Flowers | Leaves | Flowers | Leaves | Flowers | Leaves | Flowers | Leaves | Flowers | Leaves | Flowers | Leaves | Flowers | Leaves | Flowers | |
| Cultivar (C) | ||||||||||||||||
| Pinacolada | 35.37 ± 1.75 | 1.12 ± 0.06 | 1.35 ± 0.04 | 2.08 ± 0.11 | 60.68 ± 1.67 | 17.25 ± 0.89 | 11.00 ± 0.44 | 0.42 ± 0.02 | 4.51 ± 0.13 | 0.77 ± 0.04 | 0.35 ± 0.02 | 0.41 ± 0.01 | 0.80 ± 0.19 | 0.62 ± 0.12 | 18.58 ± 1.34 | 4.82 ± 0.19 |
| Radost | 45.42 ± 1.85 | 2.70 ± 0.23 | 1.01 ± 0.04 | 1.76 ± 0.12 | 69.52 ± 1.09 | 18.18 ± 1.12 | 9.63 ± 0.38 | 0.53 ± 0.03 | 4.78 ± 0.13 | 0.87 ± 0.04 | 0.28 ± 0.02 | 0.28 ± 0.01 | 0.77 ± 0.10 | 0.68 ± 0.11 | 17.16 ± 1.04 | 4.63 ± 0.19 |
| t-test | *** | *** | *** | * | *** | ns | ** | *** | ns | ns | * | *** | ns | ns | ns | ns |
| Biostimulant (B) | ||||||||||||||||
| Control | 40.42 ± 4.23 b | 2.27 ± 0.49 a | 1.13 ± 0.05 | 2.09 ± 0.13 | 60.47 ± 2.48 b | 19.65 ± 0.63 a | 9.57 ± 0.34 b | 0.42 ± 0.02 b | 4.19 ± 0.07 b | 0.85 ± 0.06 ab | 0.26 ± 0.01 b | 0.35 ± 0.03 ab | 0.42 ± 0.02 b | 1.16 ± 0.14 a | 20.75 ± 0.77 a | 5.44 ± 0.23 a |
| PH A | 35.47 ± 1.29 c | 1.63 ± 0.34 b | 1.20 ± 0.11 | 1.75 ± 0.25 | 68.59 ± 1.59 a | 15.55 ± 1.15 b | 9.13 ± 0.54 b | 0.45 ± 0.02 b | 4.71 ± 0.20 ab | 0.80 ± 0.04 ab | 0.27 ± 0.03 b | 0.33 ± 0.04 b | 1.32 ± 0.27 a | 0.63 ± 0.03 b | 20.22 ± 1.60 a | 4.83 ± 0.22 ab |
| PH V1 | 48.57 ± 2.05 a | 2.36 ± 0.44 a | 1.27 ± 0.10 | 2.16 ± 0.14 | 66.69 ± 2.53 ab | 21.04 ± 0.34 a | 12.10 ± 0.60 a | 0.59 ± 0.04 a | 4.98 ± 0.08 a | 0.94 ± 0.04 a | 0.37 ± 0.02 a | 0.39 ± 0.03 a | 0.47 ± 0.05 b | 0.28 ± 0.03 b | 12.45 ± 0.75 b | 4.07 ± 0.01 c |
| PH V2 | 37.13 ± 2.14 bc | 1.38 ± 0.23 b | 1.13 ± 0.11 | 1.67 ± 0.10 | 64.64 ± 3.28 ab | 14.63 ± 1.24 b | 10.45 ± 0.30 b | 0.43 ± 0.04 b | 4.71 ± 0.22 ab | 0.69 ± 0.06 b | 0.34 ± 0.03 a | 0.30 ± 0.03 b | 0.94 ± 0.10 ab | 0.52 ± 0.09 b | 18.07 ± 0.85 a | 4.56 ± 0.20 bc |
| *** | *** | ns | ns | * | *** | *** | *** | * | * | *** | *** | ** | *** | *** | *** | |
| C × B | ||||||||||||||||
| Pinacolada × Control | 31.15 ± 1.73 f | 1.18 ± 0.02 cd | 1.15 ± 0.09 bcd | 2.20 ± 0.14 | 56.36 ± 3.47 | 18.26 ± 0.10 | 9.99 ± 0.25 | 0.40 ± 0.02 cd | 4.13 ± 0.14 | 0.90 ± 0.06 | 0.24 ± 0.01 bc | 0.41 ± 0.03 | 0.41 ± 0.04 | 1.17 ± 0.26 | 19.53 ± 1.15 ab | 5.44 ± 0.27 |
| Pinacolada × PH A | 33.39 ± 1.91 ef | 1.02 ± 0.11 cd | 1.43 ± 0.02 ab | 2.15 ± 0.28 | 67.45 ± 2.77 | 16.66 ± 0.23 | 9.94 ± 0.45 | 0.42 ± 0.03 bcd | 4.54 ± 0.35 | 0.75 ± 0.03 | 0.33 ± 0.02 ab | 0.42 ± 0.01 | 1.48 ± 0.54 | 0.59 ± 0.06 | 23.66 ± 0.44 a | 5.23 ± 0.26 |
| Pinacolada × PH V1 | 44.51 ± 1.88 bc | 1.38 ± 0.05 bcd | 1.46 ± 0.05 a | 2.17 ± 0.31 | 61.29 ± 1.11 | 20.68 ± 0.51 | 13.17 ± 0.74 | 0.51 ± 0.01 b | 5.02 ± 0.15 | 0.87 ± 0.03 | 0.41 ± 0.02 a | 0.46 ± 0.00 | 0.36 ± 0.02 | 0.27 ± 0.05 | 11.77 ± 1.08 d | 4.07 ± 0.01 |
| Pinacolada × PH V2 | 32.45 ± 0.54 ef | 0.92 ± 0.03 d | 1.37 ± 0.05 abc | 1.78 ± 0.17 | 57.61 ± 1.74 | 13.38 ± 1.72 | 10.88 ± 0.28 | 0.34 ± 0.02 d | 4.36 ± 0.11 | 0.57 ± 0.06 | 0.40 ± 0.01 a | 0.36 ± 0.02 | 0.96 ± 0.20 | 0.45 ± 0.02 | 19.38 ± 0.53 ab | 4.53 ± 0.27 |
| Radost × Control | 49.70 ± 0.76 ab | 3.37 ± 0.03 a | 1.10 ± 0.06 cd | 1.98 ± 0.22 | 64.58 ± 1.39 | 21.03 ± 0.27 | 9.15 ± 0.60 | 0.45 ± 0.03 bcd | 4.24 ± 0.07 | 0.81 ± 0.11 | 0.28 ± 0.01 bc | 0.30 ± 0.03 | 0.44 ± 0.03 | 1.16 ± 0.17 | 21.98 ± 0.40 a | 5.43 ± 0.44 |
| Radost × PH A | 37.55 ± 0.57 de | 2.25 ± 0.44 b | 0.98 ± 0.07 d | 1.35 ± 0.25 | 69.73 ± 1.91 | 14.43 ± 2.31 | 8.32 ± 0.76 | 0.48 ± 0.04 bc | 4.88 ± 0.23 | 0.86 ± 0.06 | 0.22 ± 0.05 c | 0.25 ± 0.01 | 1.16 ± 0.21 | 0.67 ± 0.02 | 16.79 ± 0.93 bc | 4.42 ± 0.10 |
| Radost × PH V1 | 52.63 ± 1.00 a | 3.35 ± 0.10 a | 1.07 ± 0.08 d | 2.15 ± 0.09 | 72.09 ± 1.28 | 21.39 ± 0.44 | 11.02 ± 0.24 | 0.67 ± 0.01 a | 4.93 ± 0.10 | 1.01 ± 0.04 | 0.32 ± 0.00 ab | 0.32 ± 0.02 | 0.57 ± 0.02 | 0.29 ± 0.03 | 13.13 ± 1.08 cd | 4.07 ± 0.01 |
| Radost × PH V2 | 41.81 ± 0.86 cd | 1.83 ± 0.23 bc | 0.89 ± 0.03 d | 1.56 ± 0.11 | 71.67 ± 1.12 | 15.89 ± 1.79 | 10.03 ± 0.44 | 0.51 ± 0.03 b | 5.07 ± 0.33 | 0.80 ± 0.06 | 0.29 ± 0.01 bc | 0.25 ± 0.01 | 0.92 ± 0.10 | 0.58 ± 0.18 | 16.76 ± 1.28 bc | 4.58 ± 0.35 |
| *** | * | ** | ns | ns | ns | ns | * | ns | ns | * | ns | ns | ns | *** | ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carillo, P.; Pannico, A.; Cirillo, C.; Ciriello, M.; Colla, G.; Cardarelli, M.; De Pascale, S.; Rouphael, Y. Protein Hydrolysates from Animal or Vegetal Sources Affect Morpho-Physiological Traits, Ornamental Quality, Mineral Composition, and Shelf-Life of Chrysanthemum in a Distinctive Manner. Plants 2022, 11, 2321. https://doi.org/10.3390/plants11172321
Carillo P, Pannico A, Cirillo C, Ciriello M, Colla G, Cardarelli M, De Pascale S, Rouphael Y. Protein Hydrolysates from Animal or Vegetal Sources Affect Morpho-Physiological Traits, Ornamental Quality, Mineral Composition, and Shelf-Life of Chrysanthemum in a Distinctive Manner. Plants. 2022; 11(17):2321. https://doi.org/10.3390/plants11172321
Chicago/Turabian StyleCarillo, Petronia, Antonio Pannico, Chiara Cirillo, Michele Ciriello, Giuseppe Colla, Mariateresa Cardarelli, Stefania De Pascale, and Youssef Rouphael. 2022. "Protein Hydrolysates from Animal or Vegetal Sources Affect Morpho-Physiological Traits, Ornamental Quality, Mineral Composition, and Shelf-Life of Chrysanthemum in a Distinctive Manner" Plants 11, no. 17: 2321. https://doi.org/10.3390/plants11172321