1. Introduction
Nutrient return to soil through plant litter is a defining aspect of nutrient cycling and soil food web dynamics in forested ecosystems. Plant litter plays an important role in sustaining soil fertility, the global carbon (C) cycle, and soil biodiversity [
1]. Most research on this topic has focused on litter and soil characteristics that predict decomposition speed, but Killingbeck [
2] noted that litter nutrient concentrations may differ over time and attributed these temporal changes to variations of resorption efficiency and proficiency. Although limited in scope, several studies have discussed how the time of year influences litter nutrient deposition from plants to soils [
3,
4,
5,
6,
7,
8,
9,
10,
11,
12,
13,
14].
The prescribed planting of native trees in urban and peri-urban systems and in restoration plantings is a promising way to manage litter deposition and nutrient cycling and foster recovery of degraded areas. Recent initiatives are coordinating efforts among many agencies to improve Guam forest health and resilience with increased plantings of native plant species [
15].
Serianthes nelsonii is a critically endangered Fabaceae tree [
16,
17], and is among the native plant species that are actively being planted to address the goals. However, to orient the selection of which trees to use, there is a need expand knowledge about the potential of native trees in producing litter and contributing nutrients to the ecosystem. Several recent reports have included leaf nutrient traits for
S. nelsonii [
18,
19,
20], but nothing has been published concerning the seasonal variations of fresh or senesced leaves and other organs. Therefore, a better understanding of how nutrient content changes throughout the year is needed to appreciate the ecosystem services provided by
S. nelsonii and factor this information into habitat management plans.
This study aimed to evaluate monthly production and nutrient return to soils of leaf, fruit, and stem litter from a mature S. nelsonii tree in northern Guam. The objectives were to determine the influence of months on S. nelsonii litter quantity and quality to better understand the contributions of this species to nutrient flux in the tropical karst ecosystem.
2. Materials and Methods
The tropical wet climate of the study site in northern Guam is Af under the Köppen-Geiger classification. The karst soils that support the forests in the coastal zones of northern Guam developed in slope alluvium, loess, and residuum overlying limestone (Clayey-skeletal, gibbsitic, nonacid, isohyperthermic Lithic Ustorthents) [
21]. Litterfall dynamics were studied beneath the only known mature
S. nelsonii tree in Guam, and the tree exhibited a diameter at breast height of ca. 56 cm and maximum canopy radius of 4.8 m.
Litterfall beneath the tree was collected using six litter traps that were 50 cm in diameter and installed about 20 cm above the forest floor. The collections began in late 2012 and continued through early 2014. The details of the site and collection methods were described previously [
22]. For the purpose of this study, we pooled all litter collected during each month throughout 2013. The collection dates occurred in the first and third week of each month, so the pooled litter reported for each month included the second half of the previous month and the first half of the current month. Processing the samples began with removal of all litter components that were not from the
S. nelsonii tree. The remaining
S. nelsonii litter was separated into leaf, fruit, and stem litter. Litter from flowers was mostly abscised stamens and the volume was negligible and rarely recovered in the traps. When found, we added the flower litter to the fruit litter samples. Seeds within the fruits were extracted and either used in recovery plantings or returned to the seed bank, therefore there was no removal of propagules or any living materials during the course of this study.
For each month, the tissue from three organ types in each of six traps was dried in a forced draft oven for 48 h at 75 °C. The weight of each sample was measured. Some of the individual trap samples did not contain enough tissue volume for the elemental analysis of the litter. Therefore, the tissue of each organ type from all six traps was combined into one sample for each month. The tissue was milled to pass through 20-mesh screen. Total nitrogen (N) and total C were determined by dry combustion [
23] (LECO CN Analyzer, LECO Corporation, St. Joseph, MI, USA). Litter minerals and metals were digested with diethylene triamine pentaacetic acid and were quantified by inductively coupled plasma optical emission spectrometry [
24] (Spectro Genesis; SPECTRO Analytical Instruments, Kleve, Germany). Elements included in the analysis were phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), manganese (Mn), iron (Fe), zinc (Zn), boron (B), and copper (Cu).
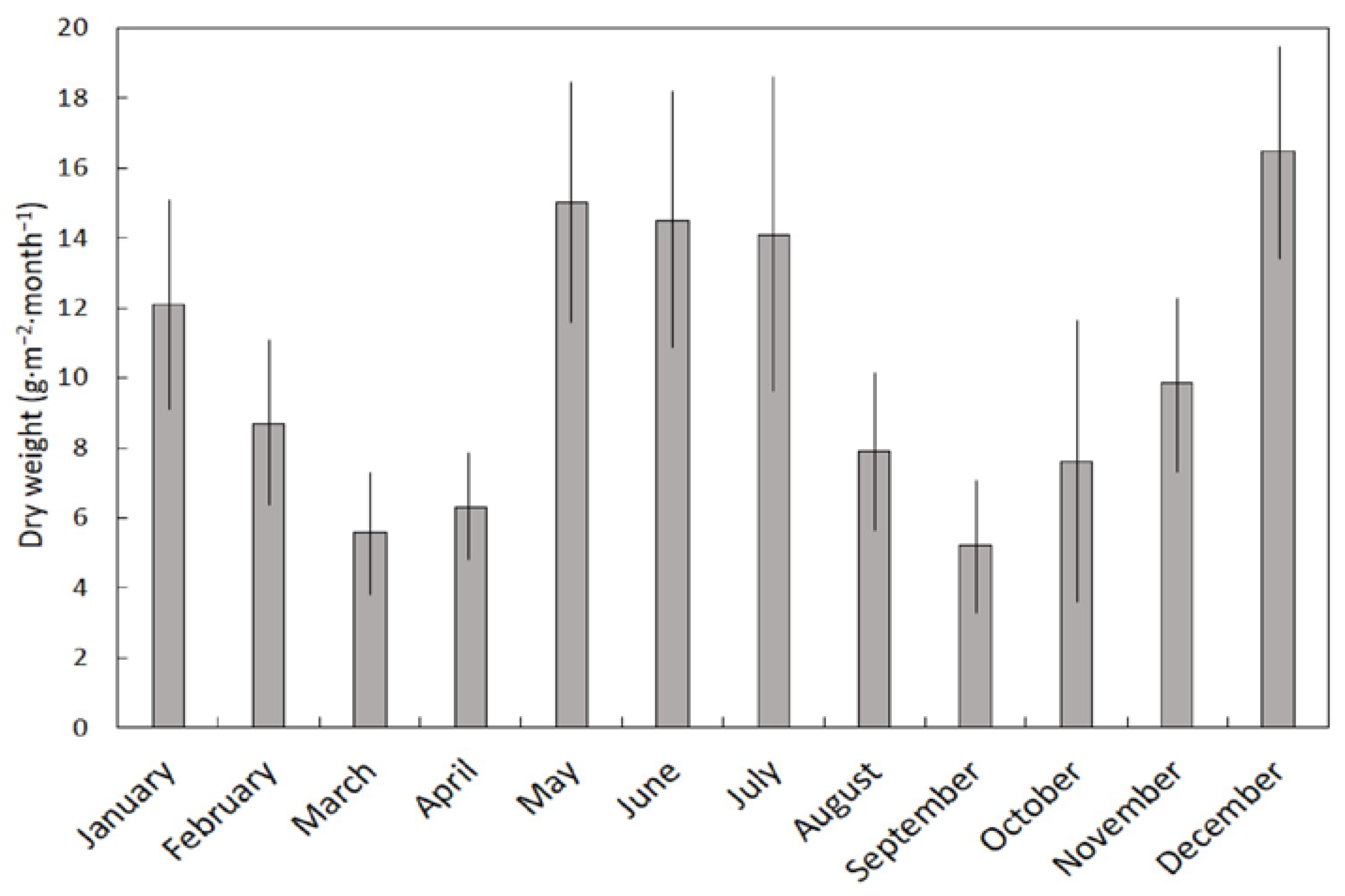
The concentrations of the minerals and metals were used to calculate the estimated pool of each element in each litter trap to provide six replications per month for the total tissue dry weight and the total weight of each mineral and metal. For this purpose, the weights per trap were converted to g, mg, or µg per square meter. The data did not conform to prerequisites for use of parametric analytical methods. Therefore, the data were subjected to the Kruskal–Wallis H test (SAS Institute, Cary, NC, USA) to determine differences among the 12 months for each of the response variables. A variability index (VI) was calculated for each response variable as (monthly maximum − monthly minimum)/monthly maximum.
4. Discussion
Nutrient return to soil through plant litter plays an important role in sustaining soil fertility and many aspects of ecosystem health [
1]. One inadequately studied aspect of litter production is the temporal dynamics of litterfall variation [
2,
3,
4,
5,
6,
7,
8,
9,
10,
11,
12,
13,
14]. Highly contrasting monthly litterfall is often correlated to growing versus non-growing seasons based on temperatures [
4,
9,
11,
12] or rainy versus dry seasons [
5,
6,
7,
13,
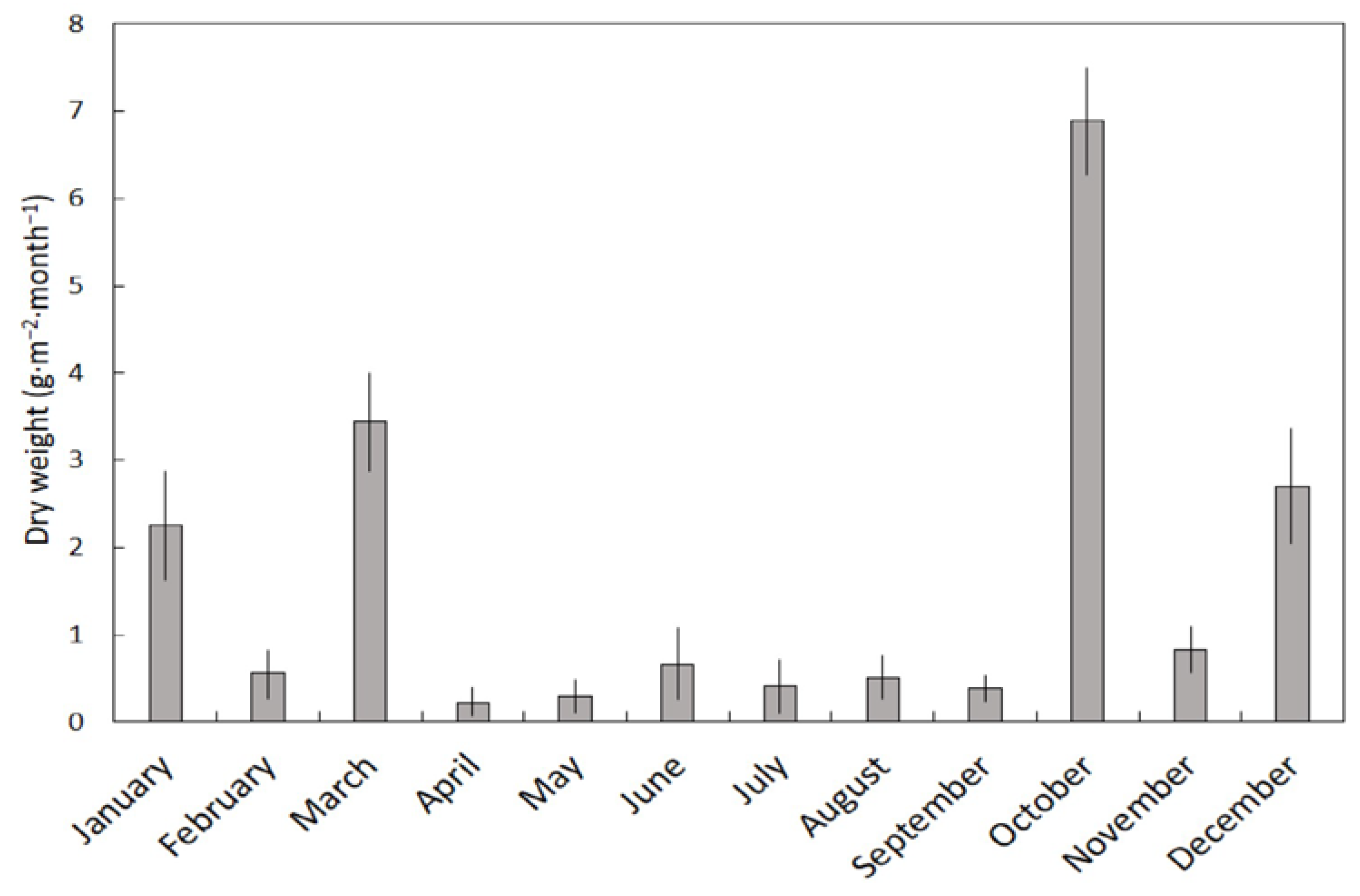
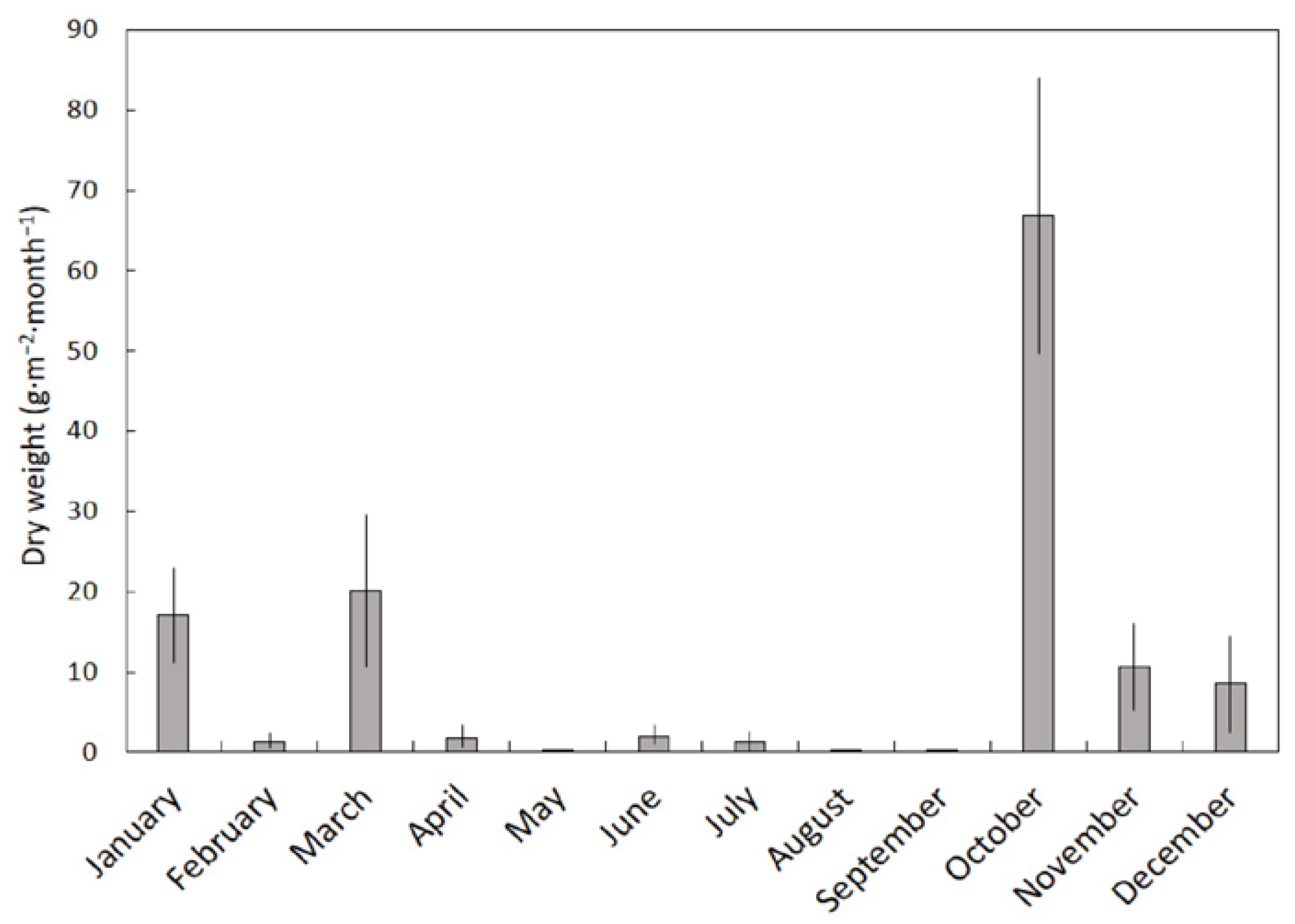
14]. The benign climate of the Mariana Islands is not characterized by strong seasonal patterns. January–May comprises the dry season, and July–November comprises the rainy season. The leaf litterfall we report herein did not differ between the two seasons, but the stem and fruit litterfall was greater in the rainy season. As a result, 949 kg·ha
−1 of
S. nelsonii litterfall occurred in the dry season and 1329 kg·ha
−1 occurred in the rainy season. However, these results were caused by stochastic high wind events rather than rainfall patterns per se. The strongest winds during the study period occurred on 20 September 2013 with maximum winds of 84 km·h
−1, and the litterfall that resulted from this wind event was collected during the first of our October collection dates. This one wind event caused the October collection dates to account for 51% of the annual stem litter and 36% of the annual fruit litter. The seasonal patterns of litterfall in Pohnpei, a second island in Micronesia, exhibited a similar increase in litterfall during the rainy season, but the causes were not discussed [
14].
Our total litter inputs for the year accumulated to 2728 kg·ha
−1, an amount that was less than reported for other tropical regions e.g., [
14,
25,
26,
27,
28]. However, our methods were unique, in that we focused on litterfall from the single species
S. nelsonii, and therefore removed the litter that originated from other sympatric tree species. The dry weight of the non-
Serianthes litter that was collected from our traps was 4435 kg·ha
−1, indicating total annual litterfall accumulated to 7163 kg·ha
−1. This total litterfall was more in congruence with the earlier reports from tropical forests [
14,
25,
26,
27,
28]. We did not separate the non-
Serianthes litter of the other tree species. However, common tree species in the habitat included
Aglaia mariannensis Merr.,
Eugenia reinwardtiana (Blume) DC.,
Ficus prolixa G.Forst.,
Macaranga thompsonii Merr.,
Meiogyne cylindrocarpa (Burck) Heusden,
Ochrosia mariannensis A.DC., and
Ochrosia oppositifolia (Lam.) K.Schum.
The past and ongoing research into the agenda of forest ecosystem services has included C and N as high priority topics [
29]. The relative proportion of these two elements in litter is one of the more important traits used to predict litter decomposition speed [
30,
31]. In turn, decomposition speed operates in concert with litterfall quantity and timing to define the volume of standing litter on the forest floor. The spatial heterogeneity of standing litter volume has direct impact on many aspects of forest ecology [
32] and creates different regeneration niches, which contribute to species diversity [
33,
34]. Litter layer integrity may also influence localized soil nutrition by reducing the risk of erosion [
35]. In this study, the predicted speed of decomposition of
S. nelsonii litter based on C/N was in the order wood < fruit < leaf. The relatively even timing of labile leaf litterfall throughout the year contrasted sharply with the pulsed timing of recalcitrant wood litter inputs, and the interplay of these behaviors may be part of the controlling factors that define the spatiotemporal characteristics of the litter layer in this tropical karst forest.
Environmental factors interact with litter quality to control the speed of plant litter decomposition. Among these factors are the amount and timing of rainfall. Indeed, decomposition speed is more rapid during high rainfall seasons than low rainfall seasons [
36,
37,
38,
39,
40]. Developing a full understanding of how
S. nelsonii participates in the nutrient flux of this tropical karst forest will require more research on seasonal aspects of actual litter decomposition to augment our results which focused on litter inputs. The unique characteristics of
S. nelsonii litter decomposition may help to better understand the means by which the tree creates unique biogeochemical spatial niches [
41].
The forests in the Mariana Islands experience more tropical cyclones than in any other state or territory of the United States [
42]. These extreme wind events occur on Guam with such frequency [
43] that they must be factored into ecosystem management decisions. Climate change models predict a greater intensity of tropical cyclones in the future [
44]. Our findings that ephemeral high wind events are a dominant factor controlling the annual litterfall cycle in this tropical karst forest indicate that the predicted changes in tropical cyclone intensity may impose extreme changes in nutrient flux in the region.
The litter component that was dominant in our study was stem tissue, causing leaf litter to account for only 45% of the annual litterfall. Leaf tissue accounts for about 70% of litterfall in most forests globally [
45]. We suggest that the frequency of tropical cyclones and extreme wind events in the Mariana Islands is responsible for the greater representation of stem tissue in this Guam forest litter. These observations illuminate the need for more litterfall studies to determine if all forest habitats in the Mariana Islands exhibit increased stem tissue and decreased leaf tissue in litterfall relative to global patterns.
Extinction risk for more than half of the world’s threatened species may not be addressed adequately unless species-specific recovery plans are developed by knowledgeable conservationists [
46]. Against this backdrop, efforts to safeguard tree diversity from continued threats has become a global challenge [
47,
48]. Addressing this challenge will demand the use of adaptive management approaches, whereby non-destructive experimental and monitoring approaches that generate new evidence are integrated into programming [
49]. In support of these global efforts, federal agencies are attempting to coordinate efforts to improve Guam’s forest health and resilience by increasing native plant coverage [
15].
Serianthes nelsonii is among the list of native plant species that are actively being planted to address this initiative. The 28-year-old national recovery plan for this tree species [
50] has been inadequately implemented, partly due to historical choices to ignore the value of funding adaptive management research conducted by competent scientists to generate new knowledge [
51]. The failures of meeting the goals of the recovery plan reveal how past decision makers can create a level of distrust that impedes continued recovery efforts, particularly where militarization is part of the conservation model, such as the case in Guam [
52]. Our methods provide an example of how non-destructive methods may be used by practitioners to generate new evidence that can be factored into conservation programming to inform species and habitat recovery plans. This snapshot of
S. nelsonii litterfall behaviors in 2013 also provides a benchmark for future studies to use repeatable methods to quantify alterations in litterfall that accompany future climate change [
53].







{kind=link}
{kind=link}
{kind=link}