
Effects of Nitrogen and Phosphorus Regulation on Plant Type, Population Ecology and Sheath Blight of Hybrid Rice
Abstract
:1. Introduction
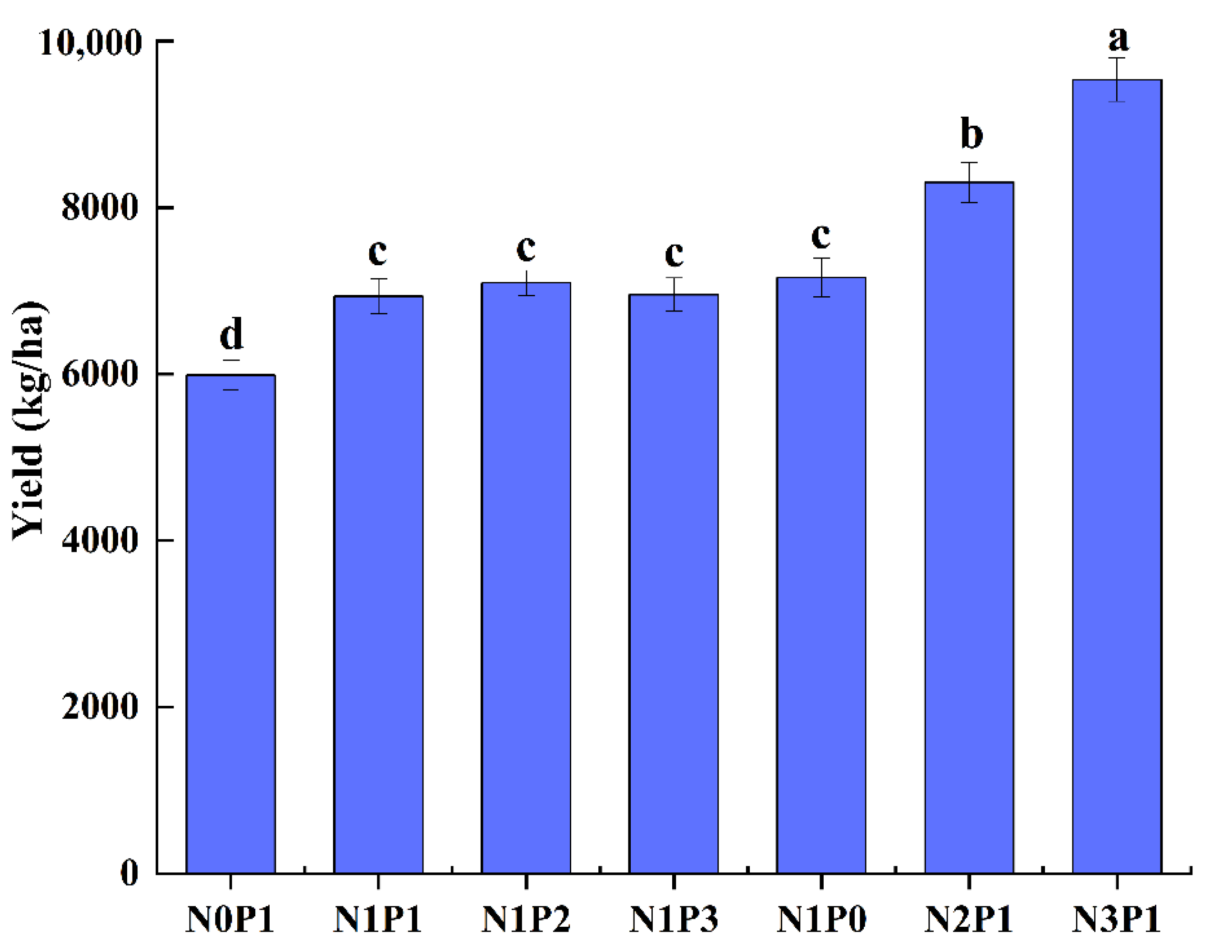
2. Results
2.1. Effects of N and P Regulation on Rice Plant Morphology
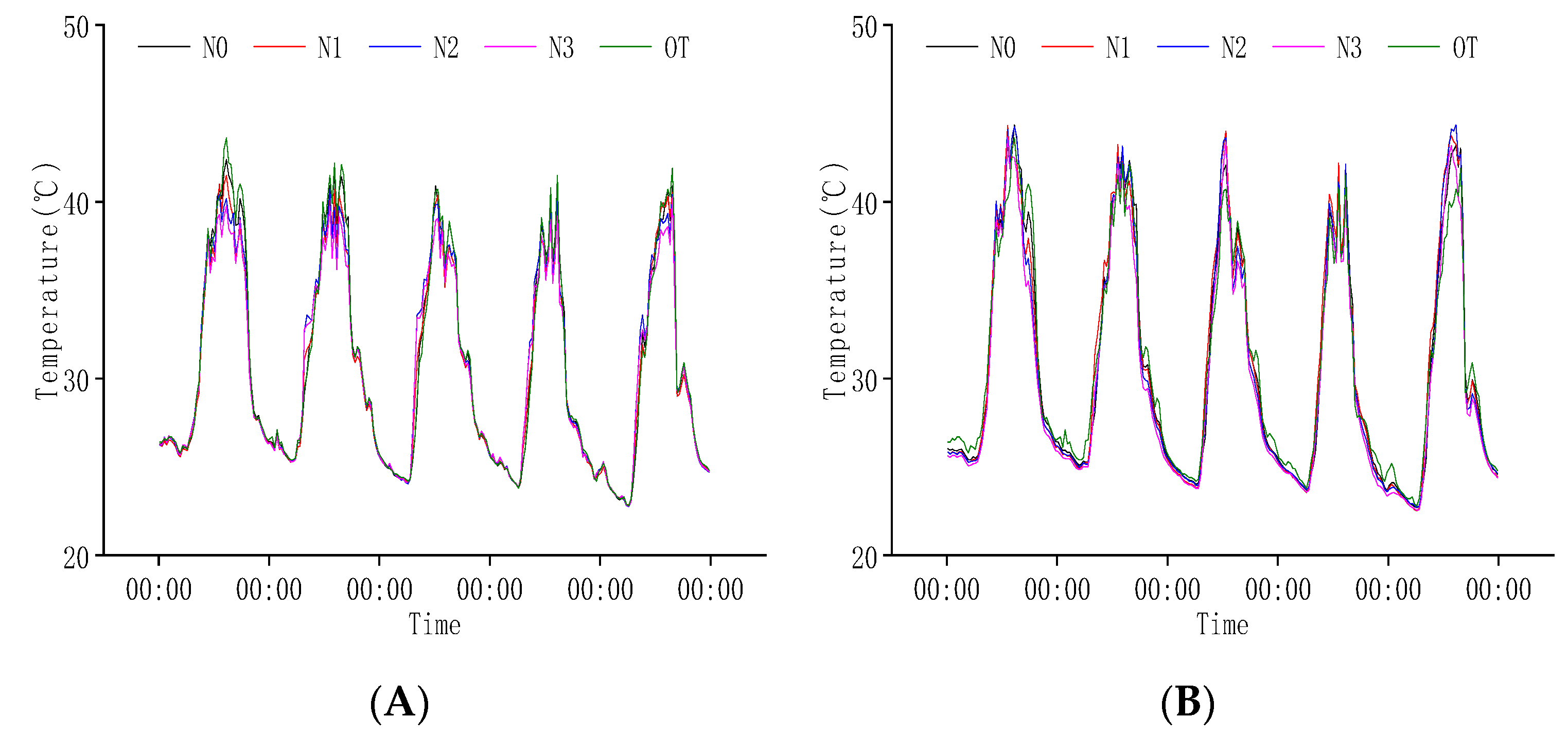
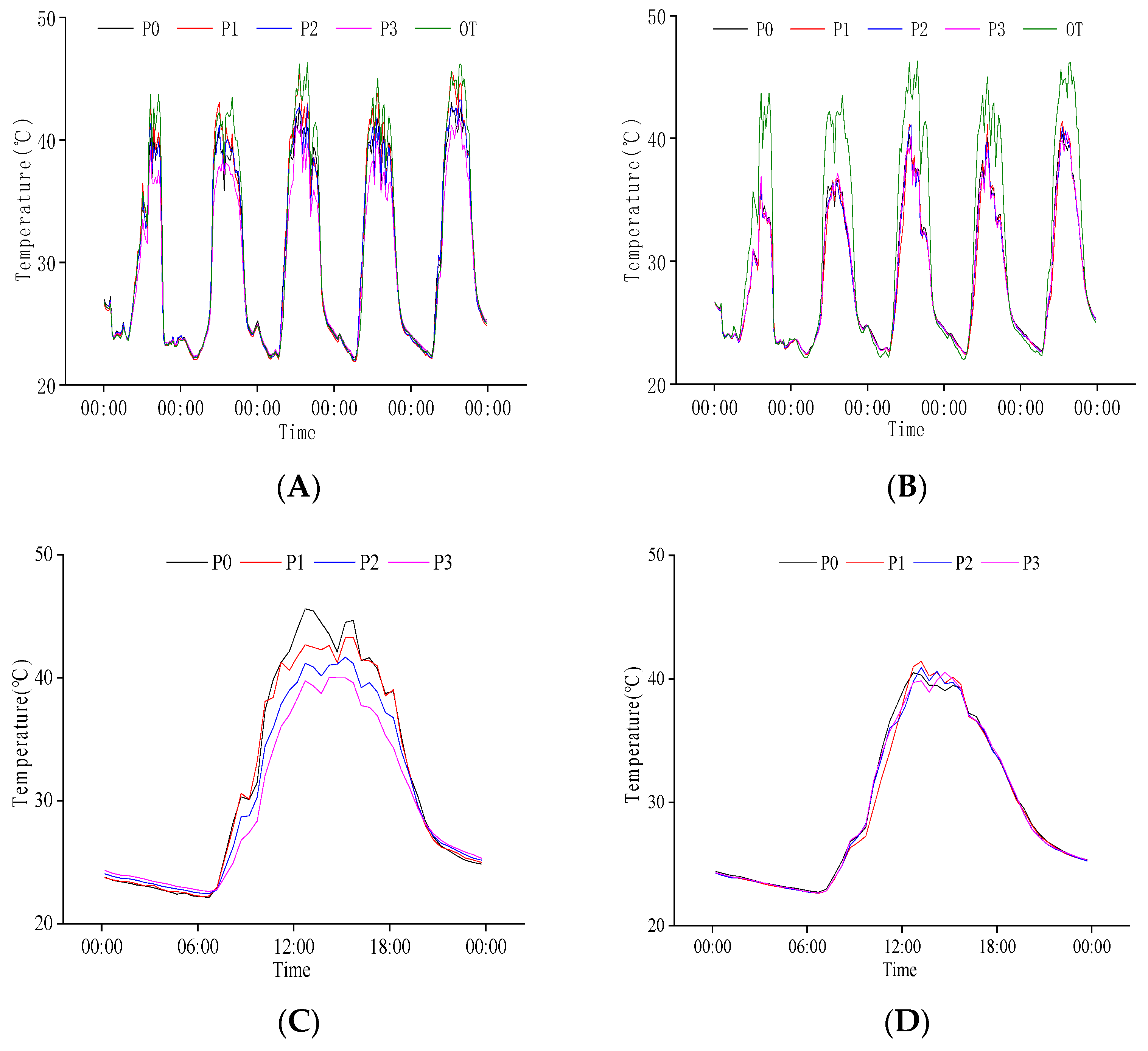
2.2. Response of Temperature and Humidity to Rice Population and N Regulation
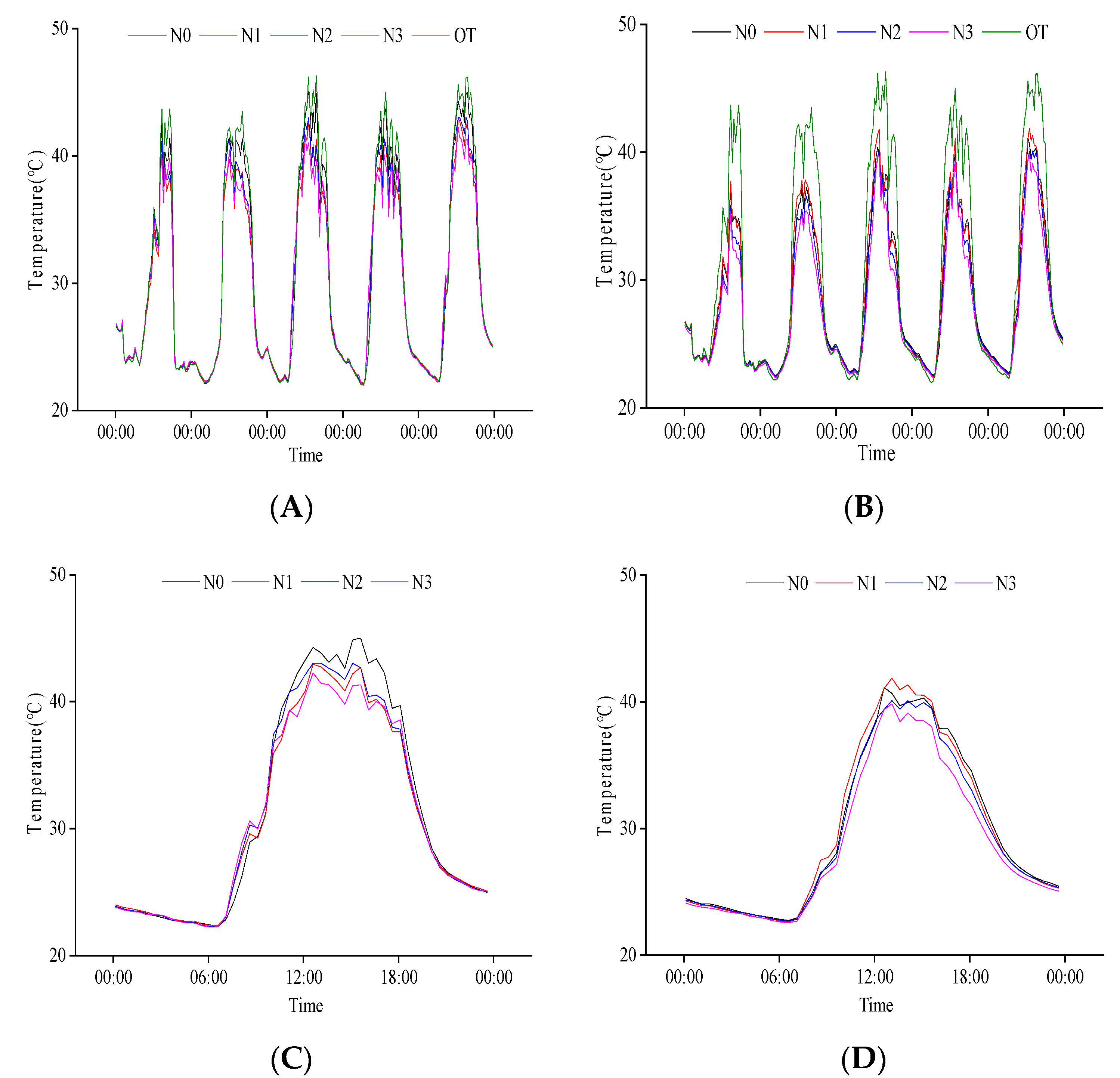
2.2.1. Response of Population Temperature at Heading Stage to N Regulation
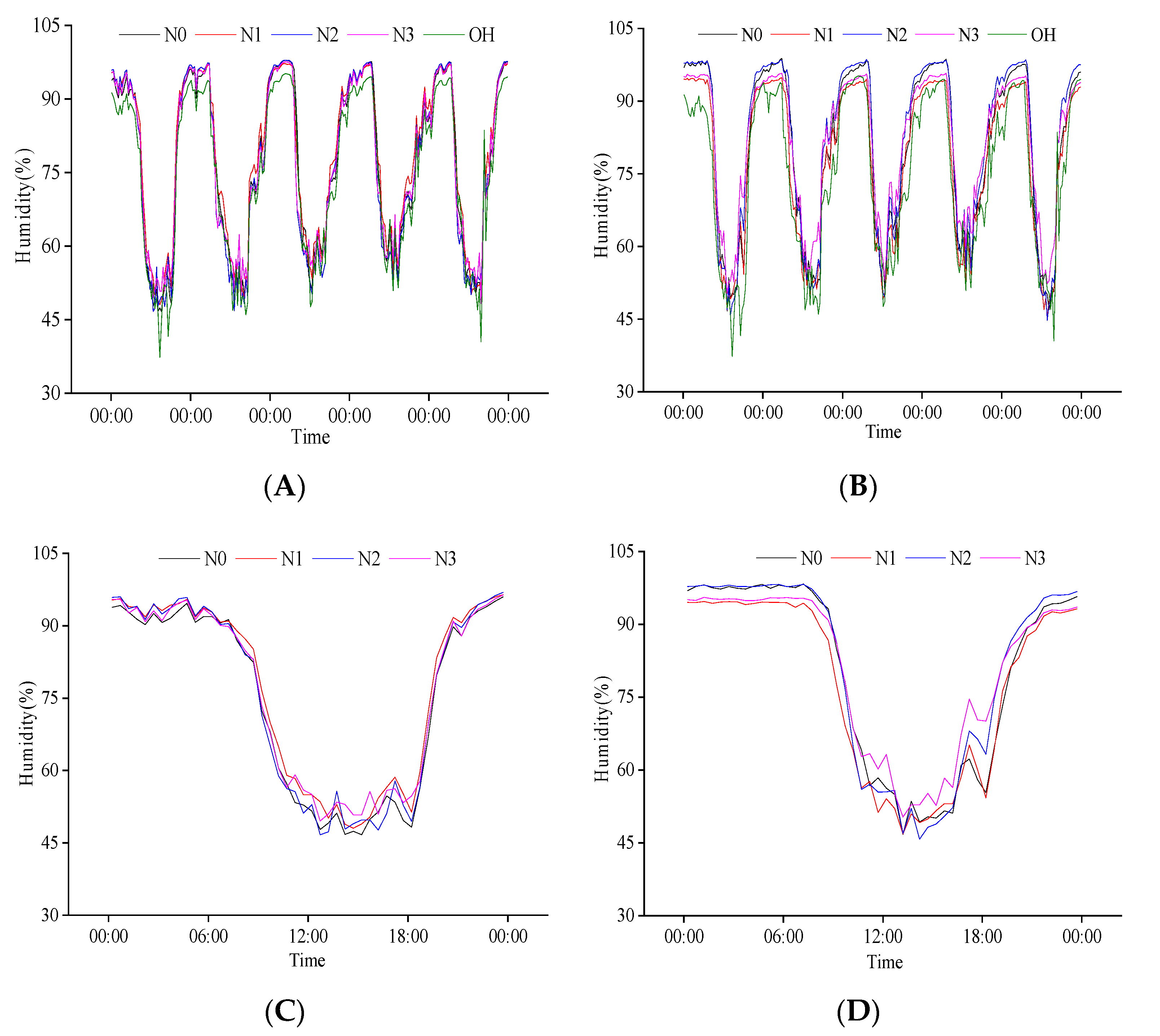
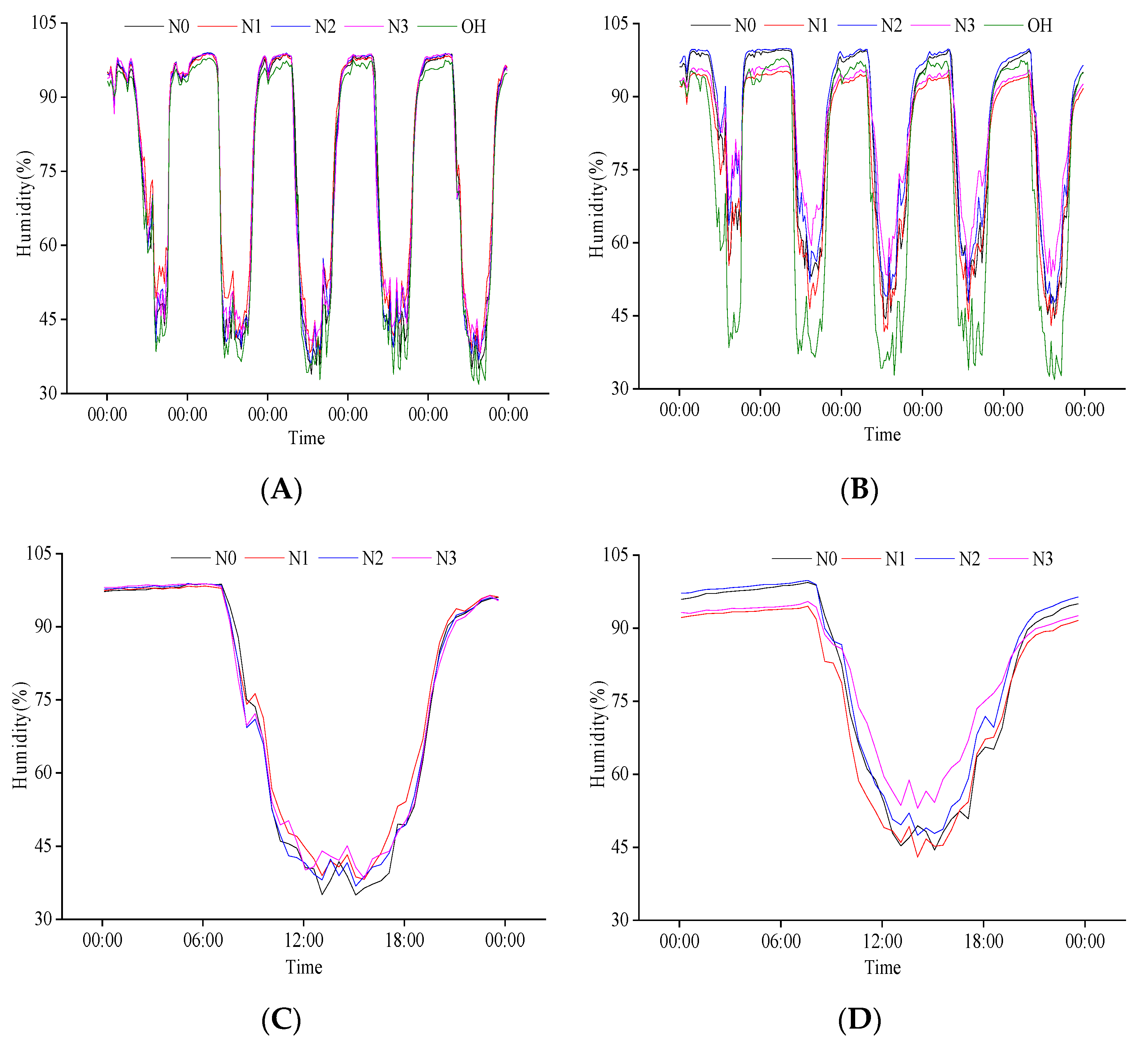
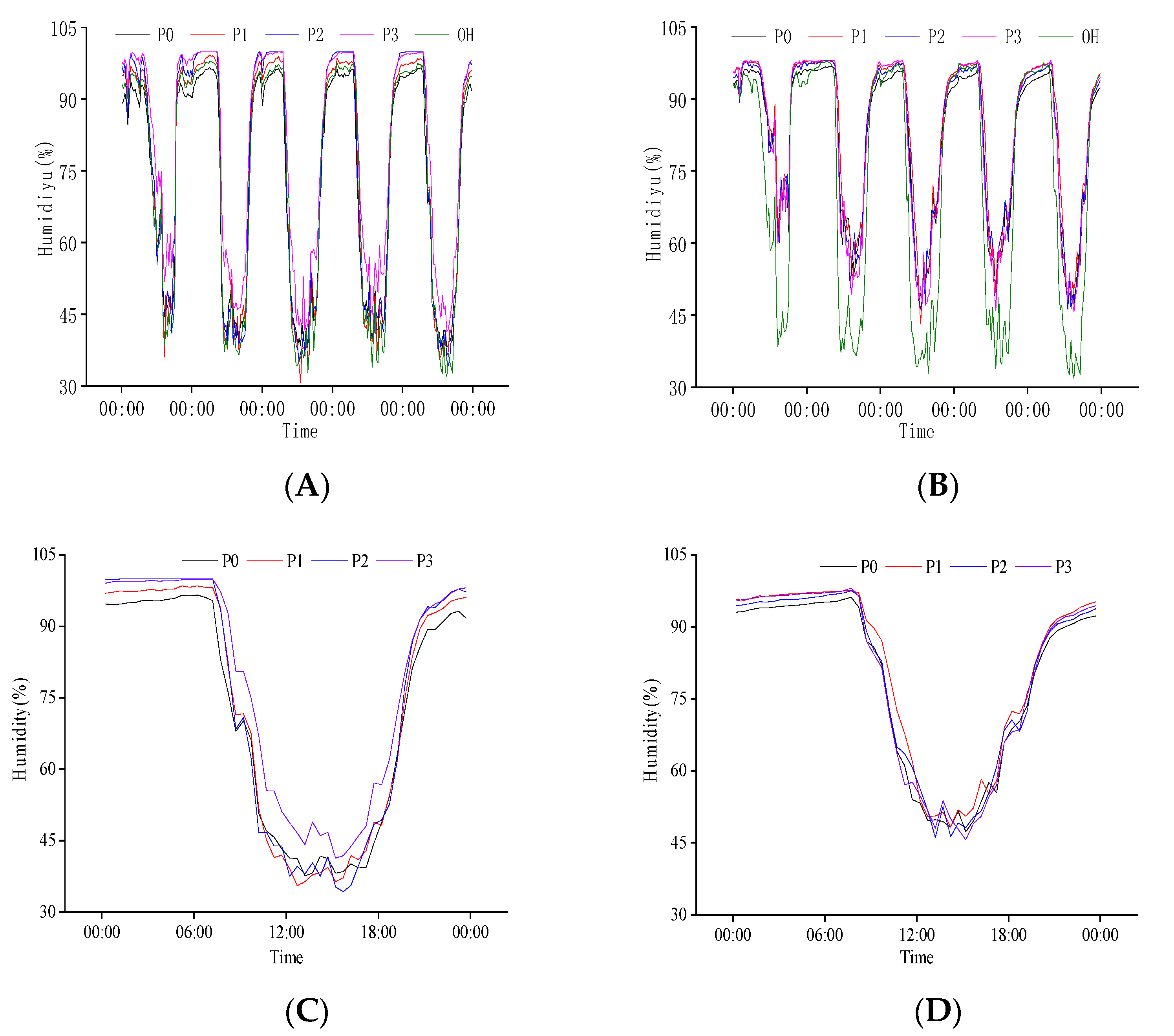
2.2.2. Response of Population Humidity to N Regulation at the Heading Stage
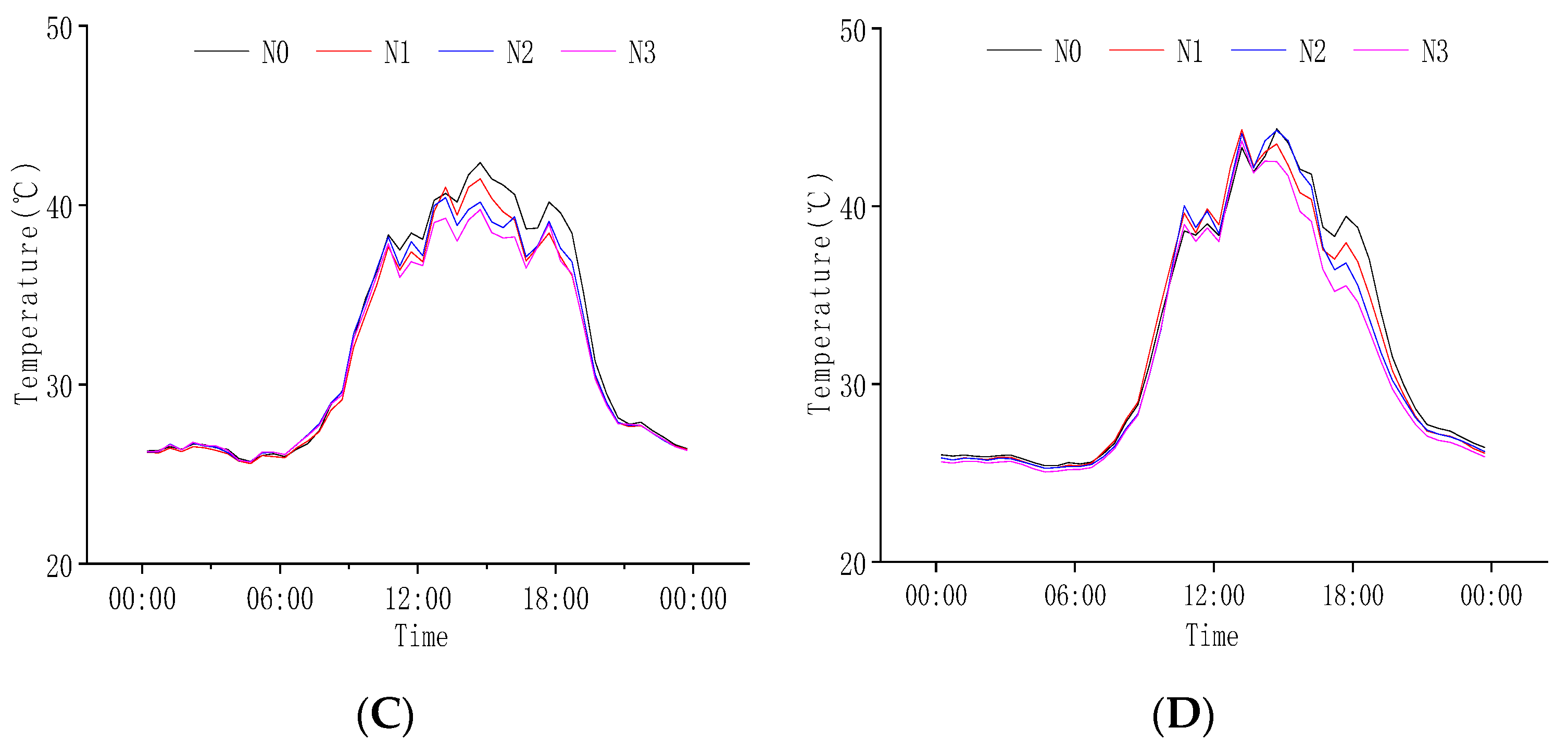
2.2.3. Response of Population Temperature to N Regulation at Grain Filling Stage
2.2.4. Response of Population Humidity to N Regulation at Grain Filling Stage
2.3. Response of Temperature and Humidity of Rice Population to P Regulation
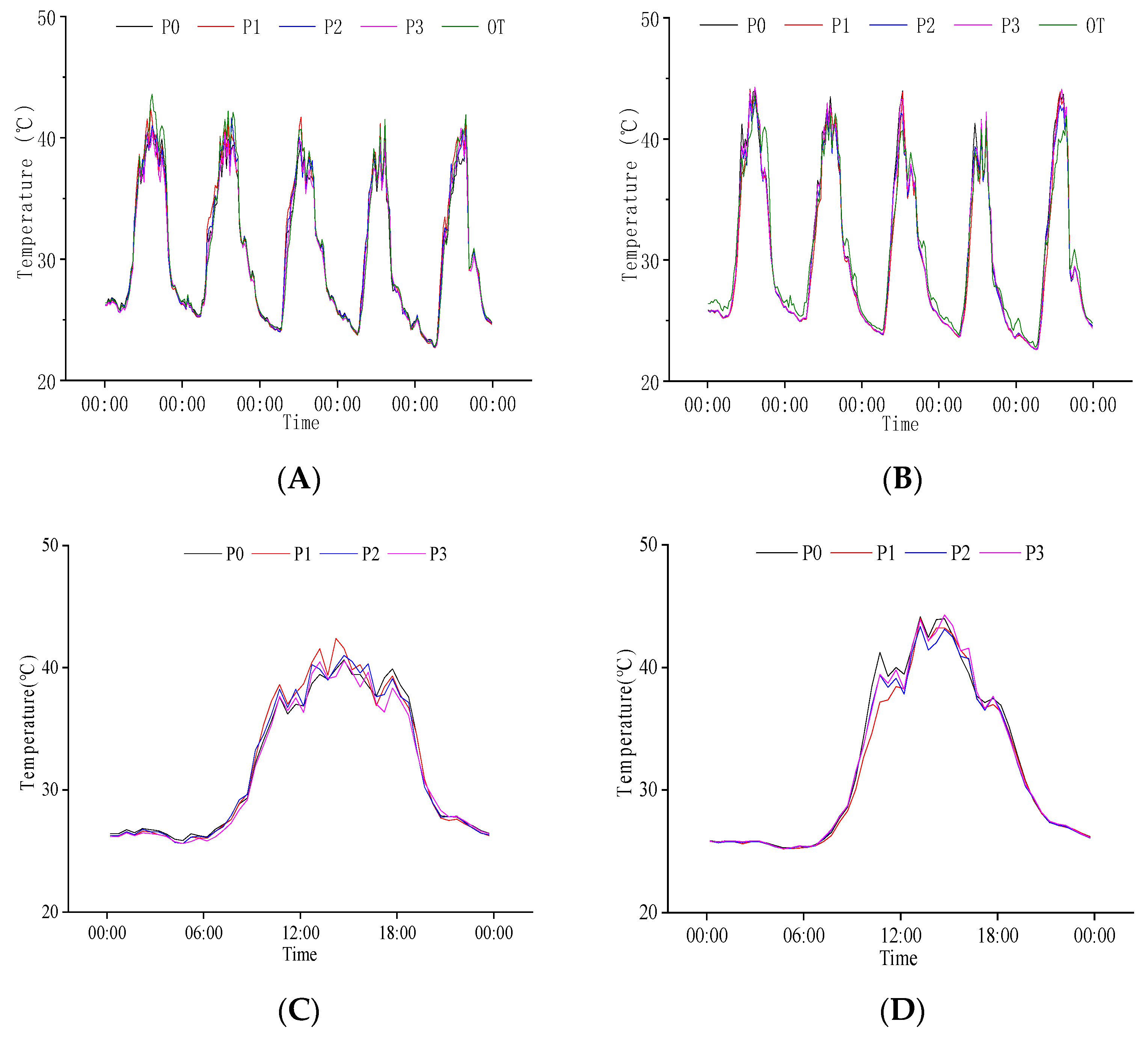
2.3.1. Response of Population Temperature at Heading Stage to P Regulation
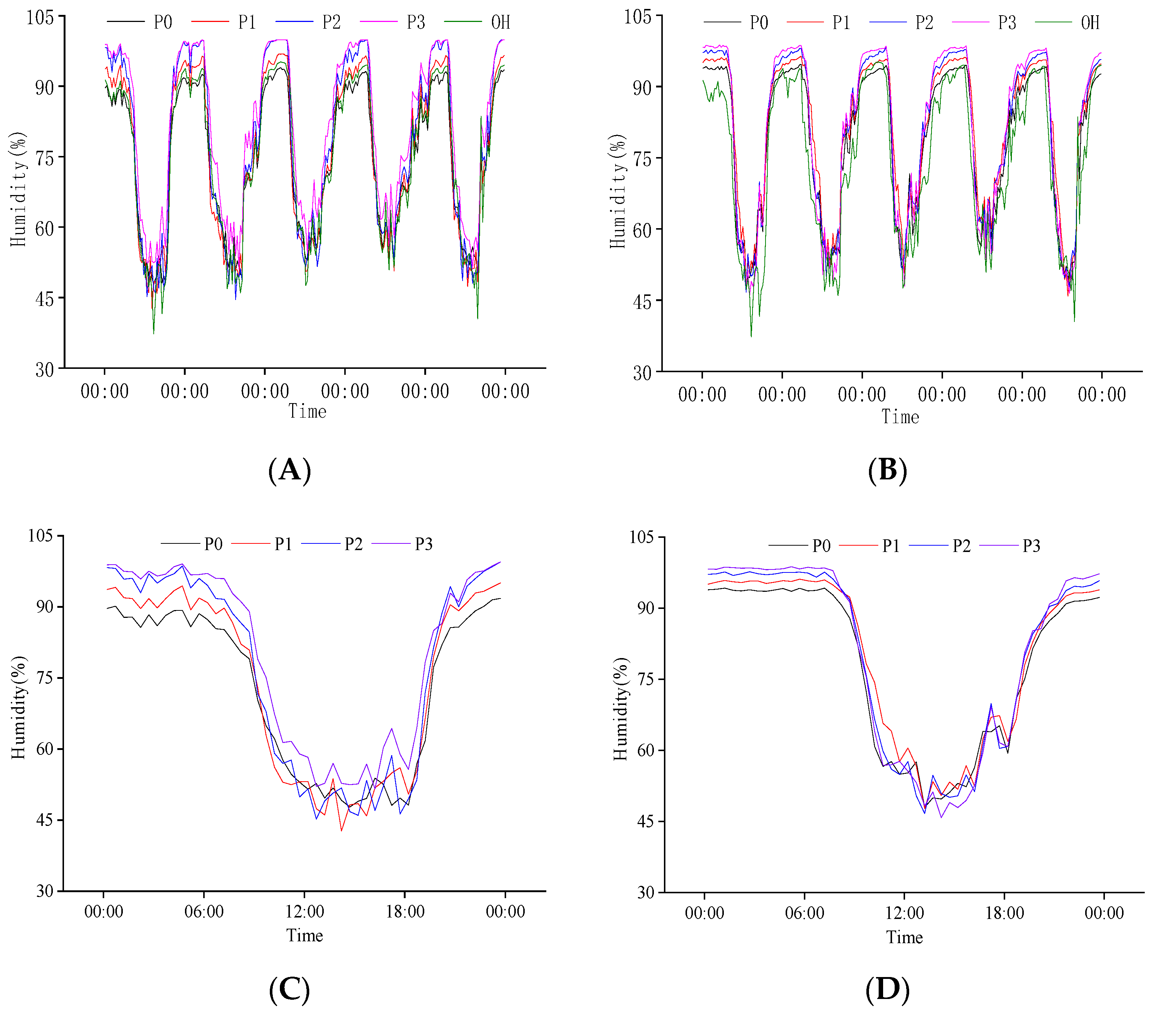
2.3.2. Response of Population Humidity to P Regulation at the Heading Stage
2.3.3. Response of Population Temperature to P Regulation at Grain Filling Stage
2.3.4. Response of Population Humidity to P Regulation at Grain Filling Stage
2.4. Effects of N and P Regulation on the Severity of Rice Sheath Blight
2.5. Correlation Analysis of N and P Regulation, Population Temperature, Humidity Change, and Sheath Blight
2.5.1. Correlation Analysis of N Regulation, Population Temperature, Humidity Variation, and Sheath Blight
2.5.2. Correlation Analysis of P Regulation, Population Temperature, Humidity Change, and Sheath Blight
3. Discussion
3.1. N and P Regulation Significantly Affected Hybrid Rice Plant Morphology
3.2. Rice Plant Type Variation Caused Population Microclimate Difference
3.3. Population Temperature and Humidity Variation Was an Important Factor Affecting the Incidence of Rice Sheath Blight
4. Materials and Methods
4.1. Experimental Design
4.2. Sampled and Determination
4.2.1. Determination of Plant Type Characteristics
4.2.2. Measurement of the Hybrid Rice Population Temperature and Humidity
4.2.3. Investigation of Sheath Blight in the Field
4.2.4. Data Analysis and Plotting
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, W.; Sakai, H.; Yagi, K.; Hasegawa, T. Interactions of elevated [CO2] and night temperature on rice growth and yield. Agric. For. Meteorol. 2009, 149, 51–58. [Google Scholar] [CrossRef]
- Guo, Z.; Liu, H.; Yuan, H.; Yang, G.; Zheng, J.; Chen, L. Insect-proof nets affect paddy field microclimate parameters and grain quality of different japonica rice varieties. J. Crop Sci. Biotechnol. 2015, 18, 73–81. [Google Scholar] [CrossRef]
- Pal, N.; Kumar, A.; Malannavar, A. Effect of temperature and pH levels on the growth and sporulation of Fusarium oxysporum f. sp. lini causing linseed wilt. Int. J. Chem. Stud. 2019, 7, 4494–4497. [Google Scholar]
- Pan, S.; Liu, H.; Mo, Z.; Patterson, B.; Duan, M.; Tian, H.; Hu, S.; Tang, X. Effects of nitrogen and shading on root morphologies, nutrient accumulation, and photosynthetic parameters in different rice genotypes. Sci. Rep. 2016, 6, 32148. [Google Scholar] [CrossRef]
- Van Kiet, H.; Nose, A.; Zheng, S.H. Effect of cold stress on root growth, accumulation of soluble proteins and free amino acids of sheath blight-resistant rice genotype 32R. Trop. Agric. Dev. 2016, 60, 191–194. [Google Scholar]
- Bhukal, N.; Singh, R.; Mehta, N. Progression and development of sheath blight of rice in relation to weather variables. J. Mycol. Plant Pathol. 2015, 45, 166–172. [Google Scholar]
- Milati, L.N.; Nuryanto, B.; Sumarlin, U. The Relationship between Sheath Blight Disease Incidence, Disease Severity, and Rice Yield. J. Fitopatol. Indones. 2021, 17, 113–120. [Google Scholar] [CrossRef]
- Richa, K.; Tiwari, I.M.; Kumari, M.; Devanna, B.; Sonah, H.; Kumari, A.; Nagar, R.; Sharma, V.; Botella, J.R.; Sharma, T.R. Functional characterization of novel chitinase genes present in the sheath blight resistance QTL: qSBR11-1 in rice line tetep. Front. Plant Sci. 2016, 7, 244. [Google Scholar] [CrossRef]
- Yadav, S.; Anuradha, G.; Kumar, R.R.; Vemireddy, L.; Sudhakar, R.; Balram, M.; Siddiq, E. Bulked Segregant Analysis to Detect Main Effect of QTL Associated with Sheath Blight Resistance in BPT-5204/ARC10531 Rice (Oryza sativa L). Rice Res. 2015, 3, 149. [Google Scholar] [CrossRef]
- Wen, Z.; Zeng, Y.; Ji, Z.; Yang, C. Mapping quantitative trait loci for sheath blight disease resistance in Yangdao 4 rice. Genet. Mol. Res. 2015, 14, 1636–1649. [Google Scholar] [CrossRef]
- Han, Y.P.; Xing, Y.Z.; Gu, S.L.; Chen, Z.X.; Pan, X.B.; Chen, X.L. Effect of morphological traits on sheath blight resistance in rice. Acta Bot. Sin. 2003, 45, 825–831. [Google Scholar]
- Slaton, N.A.; Cartwright, R.D.; Meng, J.; Gbur, E.E.; Norman, R.J. Sheath blight severity and rice yield as affected by nitrogen fertilizer rate, application method, and fungicide. Agron. J. 2003, 95, 1489–1496. [Google Scholar] [CrossRef]
- Mo, R.; Jiang, L.; Guo, L.; Hu, J.; Liu, K.; Zhou, J.; Liang, T.; Zeng, K.; Ding, C. Effect of nitrogen application on contents of different forms of nitrogen in rice plants. J. Agric. Sci. 2011, 12, 1484–1489. [Google Scholar]
- Rebitanim, N.A.; Rebitanim, N.Z.; Tajudin, N.S. Impact of silicon in managing important rice diseases: Blast, sheath blight, brown spot and grain discoloration. Int. J. Agron. Agric. Res. 2015, 6, 71–85. [Google Scholar]
- Schurt, D.A.; Cruz, M.F.; Nascimento, K.J.; Filippi, M.C.; Rodrigues, F.A. Silicon potentiates the activities of defense enzymes in the leaf sheaths of rice plants infected by Rhizoctonia solani. Trop. Plant Pathol. 2014, 39, 457–463. [Google Scholar] [CrossRef]
- Wang, J.X.; Jian, S.; Li, C.X.; Liu, H.L.; Wang, J.G.; Zhao, H.W.; Zou, D.T. Genetic dissection of the developmental behavior of plant height in rice under different water supply conditions. J. Integr. Agric. 2016, 15, 2688–2702. [Google Scholar] [CrossRef]
- Kashiwagi, T.; Ishimaru, K. Identification and functional analysis of a locus for improvement of lodging resistance in rice. Plant Physiol. 2004, 134, 676–683. [Google Scholar] [CrossRef]
- Dong, C.F.; Ding, C.L.; Xu, N.X.; Cheng, Y.H.; Shen, Y.X.; Gu, H.R. Double-purpose rice (Oryza sativa L.) variety selection and their morphological traits. Field Crops Res. 2013, 149, 276–282. [Google Scholar] [CrossRef]
- Moe, K.; Htwe, A.Z.; Thu, T.T.P.; Kajihara, Y.; Yamakawa, T. Effects on NPK status, growth, dry matter and yield of rice (Oryza sativa) by organic fertilizers applied in field condition. Agriculture 2019, 9, 109. [Google Scholar] [CrossRef]
- Biswas, A.K.; Choudhuri, M.A. Mechanism of monocarpic senescence in rice. Plant Physiol. 1980, 65, 340–345. [Google Scholar] [CrossRef]
- Xue, W.; Lindner, S.; Nay-Htoon, B.; Dubbert, M.; Otieno, D.; Ko, J.; Muraoka, H.; Werner, C.; Tenhunen, J.; Harley, P. Nutritional and developmental influences on components of rice crop light use efficiency. Agric. For. Meteorol. 2016, 223, 1–16. [Google Scholar] [CrossRef]
- Miranda-Villagómez, E.; Trejo-Téllez, L.I.; Gómez-Merino, F.C.; Sandoval-Villa, M.; Sánchez-García, P.; Aguilar-Méndez, M.Á. Nanophosphorus fertilizer stimulates growth and photosynthetic activity and improves P status in rice. J. Nanomater. 2019, 2019, 5368027. [Google Scholar] [CrossRef]
- Prasad, P.; Boote, K.; Allen, L., Jr.; Sheehy, J.; Thomas, J. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res. 2006, 95, 398–411. [Google Scholar] [CrossRef]
- Fischer, R.; Rees, D.; Sayre, K.; Lu, Z.M.; Condon, A.; Saavedra, A.L. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Garrity, D.P.; O’Toole, J.C. Selection for reproductive stage drought avoidance in rice, using infrared thermometry. Agron. J. 1995, 87, 773–779. [Google Scholar] [CrossRef]
- Deng, X.P.; Shan, L.; Ma, Y.Q.; Inanaga, S. Diurnal oscillation in the intercellular CO2 concentration of spring wheat under the semiarid conditions. Photosynthetica 2000, 38, 187–192. [Google Scholar] [CrossRef]
- Singh, P.; Mazumdar, P.; Harikrishna, J.A.; Babu, S. Sheath blight of rice: A review and identification of priorities for future research. Planta 2019, 250, 1387–1407. [Google Scholar] [CrossRef]
- Lee, F. Rice sheath blight: A major rice disease. Plant Dis. 1983, 67, 829–832. [Google Scholar] [CrossRef]
- Wu, W.; Huang, J.; Cui, K.; Nie, L.; Wang, Q.; Yang, F.; Shah, F.; Yao, F.; Peng, S. Sheath blight reduces stem breaking resistance and increases lodging susceptibility of rice plants. Field Crops Res. 2012, 128, 101–108. [Google Scholar] [CrossRef]
- Savary, S.; Castilla, N.; Elazegui, F.; McLaren, C.; Ynalvez, M.; Teng, P. Direct and indirect effects of nitrogen supply and disease source structure on rice sheath blight spread. Phytopathology 1995, 85, 959–965. [Google Scholar] [CrossRef]
- Yang, G.; Wang, X.; Peng, Y.; Rasul, F.; Hu, Y.; Zou, T. Different micro-climate response of indica rice population to nitrogen fertilizer. Plant Soil Environ. 2018, 64, 407–412. [Google Scholar]
- Tang, Q.; Peng, S.; Buresh, R.J.; Zou, Y.; Castilla, N.P.; Mew, T.W.; Zhong, X. Rice varietal difference in sheath blight development and its association with yield loss at different levels of N fertilization. Field Crops Res. 2007, 102, 219–227. [Google Scholar] [CrossRef]
- Sathe, A.P.; Kumar, A.; Mandlik, R.; Raturi, G.; Yadav, H.; Kumar, N.; Shivaraj, S.; Jaswal, R.; Kapoor, R.; Gupta, S.K.; et al. Role of silicon in elevating resistance against sheath blight and blast diseases in rice (Oryza sativa L.). Plant Physiol. Biochem. 2021, 166, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.Á.; Vale, F.X.; Datnoff, L.E.; Prabhu, A.S.; Korndörfer, G.H. Effect of rice growth stages and silicon on sheath blight development. Phytopathology 2003, 93, 256–261. [Google Scholar] [PubMed]
- Khoshkdaman, M.; Mousanejad, S.; Elahinia, S.A.; Ebadi, A.A.; Padasht-Dehkaei, F. Sheath blight development and yield loss on rice in different epidemiological conditions. J. Plant Pathol. 2021, 103, 87–96. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Length (cm) | Leaf Basal Angle (°) | Leaf Opening Angle (°) | Plant Height (cm) | Effective Panicle (×103/ha) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D1 | D2 | D3 | D1 | D2 | D3 | D1 | D2 | D3 | |||
| N0P1 | 22.00 c | 36.83 d | 47.23 c | 17.77 a | 36.77 d | 51.30 cd | 17.50 ab | 40.43 b | 45.07 e | 110.83 d | 1845.00 c |
| N1P1 | 29.93 a | 41.40 b | 51.53 b | 15.50 b | 41.37 c | 49.53 d | 18.47 a | 37.30 c | 49.40 d | 116.67 bc | 1920.00 bc |
| N1P2 | 30.30 a | 44.10 ab | 56.33 a | 15.53 b | 44.07 b | 49.83 d | 18.73 a | 46.43 a | 53.73 c | 118.67 b | 1785.00 c |
| N1P3 | 29.23 a | 47.23 a | 58.20 a | 19.33 a | 47.13 a | 54.47 b | 15.50 b | 37.43 c | 59.50 a | 114.33 cd | 1995.00 bc |
| N1P0 | 23.97 b | 34.97 d | 52.03 b | 15.43 b | 35.47 d | 60.13 a | 12.03 c | 46.37 a | 48.53 d | 114.00 cd | 2010.00 ab |
| N2P1 | 22.20 c | 37.57 c | 49.50 bc | 13.27 b | 41.40 c | 50.53 cd | 16.60 ab | 39.30 bc | 53.30 c | 120.00 b | 2295.00 a |
| N3P1 | 29.50 a | 43.43 b | 56.37 a | 12.33 b | 43.53 b | 52.27 bc | 16.70 ab | 46.70 a | 56.77 b | 128.00 a | 2025.00 ab |
| N | 58.63 ** | 22.04 ** | 28.99 ** | 28.42 ** | 49.87 ** | 3.50 ns | 2.90 ns | 37.80 ** | 57.36 ** | 59.64 ** | 6.66 ** |
| P | 26.53 ** | 60.86 ** | 20.38 ** | 17.81 ** | 117.26 ** | 64.38 ** | 37.84 ** | 62.48 ** | 56.83 ** | 5.54 * | 1.82 ns |
| P0 | P1 | P2 | P3 | |
|---|---|---|---|---|
| N0 | — | 3.41 ± 0.67 e | — | — |
| N1 | 10.69 ± 2.14 c | 8.85 ± 1.10 cd | 6.62 ± 0.54 de | 4.28 ± 1.19 e |
| N2 | — | 17.51 ± 1.00 b | — | — |
| N3 | — | 31.46 ± 2.22 a | — | — |
| Stages | N | Canopy Temperature | Baselayer Temperature | Canopy Humidity | Baselayer Humidity | |
|---|---|---|---|---|---|---|
| Heading stage | N | 1.00 | ||||
| Canopy temperature | −0.47 ns | 1.00 | ||||
| Base layer temperature | 0.52 ns | −0.96 * | 1.00 | |||
| Canopy humidity | 0.11 ns | −0.81 * | 0.63 ns | 1.00 | ||
| Base layer humidity | −0.76 ns | 0.57 ns | −0.75 ns | 0.01 ns | 1.00 | |
| Severity of diseases | 0.99 ** | −0.56 ns | 0.64 ns | 0.15 ns | −0.84 * | |
| Grain filling stage | N | 1.00 | ||||
| Canopy temperature | 0.26 ns | 1.00 | ||||
| Base layer temperature | 0.58 ns | −0.63 | 1.00 | |||
| Canopy humidity | −0.74 ns | −0.84 * | 0.11 ns | 1.00 | ||
| Base layer humidity | −0.88 * | 0.09 ns | −0.79 ns | 0.41 ns | 1.00 | |
| Severity of diseases | 0.99 ** | 0.12 ns | 0.69 ns | −0.64 ns | −0.94 * |
| Stages | P | Canopy Temperature | Baselayer Temperature | Canopy Humidity | Baselayer Humidity | |
|---|---|---|---|---|---|---|
| Heading stage | P | 1.00 | ||||
| Canopy temperature | 0.99 ** | 1.00 | ||||
| Base layer temperature | 0.78 ns | 0.72 ns | 1.00 | |||
| Canopy humidity | −0.37 ns | −0.45 ns | 0.29 ns | 1.00 | ||
| Base layer humidity | 0.05 ns | 0.14 ns | −0.58 ns | −0.94 * | 1.00 | |
| Severity of diseases | −0.81 * | −0.81 * | −0.46 ns | 0.55 ns | −0.25 ns | |
| Grain filling stage | P | 1.00 | ||||
| Canopy temperature | 0.99 ** | 1.00 | ||||
| Base layer temperature | −0.05 ns | −0.11 ns | 1.00 | |||
| Canopy humidity | −0.83 * | −0.89 * | 0.53 ns | 1.00 | ||
| Base layer humidity | −0.19 ns | −0.10 ns | −0.94 * | −0.36 ns | 1.00 | |
| Severity of diseases | −0.81 * | −0.90 * | 0.20 ns | 0.91 * | −0.12 ns |
| Grade | Severity Classification Criteria |
|---|---|
| Grade 0 | The whole plant is disease-free |
| Grade 1 | The disease starts from basal leaf and sheath |
| Grade 2 | The disease starts from the leaf sheath or leaf below the third leaf (from the top leaf, the same below) |
| Grade 3 | The disease starts from the leaf sheath or leaf below the second leaf |
| Grade 4 | Parietal leaf sheath or parietal leaf disease |
| Grade 5 | Whole plant disease and wither |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Liu, R.; Ma, P.; Chen, H.; Zhang, R.; Wang, X.; Li, Y.; Hu, Y. Effects of Nitrogen and Phosphorus Regulation on Plant Type, Population Ecology and Sheath Blight of Hybrid Rice. Plants 2022, 11, 2306. https://doi.org/10.3390/plants11172306
Yang G, Liu R, Ma P, Chen H, Zhang R, Wang X, Li Y, Hu Y. Effects of Nitrogen and Phosphorus Regulation on Plant Type, Population Ecology and Sheath Blight of Hybrid Rice. Plants. 2022; 11(17):2306. https://doi.org/10.3390/plants11172306
Chicago/Turabian StyleYang, Guotao, Rong Liu, Peng Ma, Hong Chen, Rongping Zhang, Xuechun Wang, Yongyan Li, and Yungao Hu. 2022. "Effects of Nitrogen and Phosphorus Regulation on Plant Type, Population Ecology and Sheath Blight of Hybrid Rice" Plants 11, no. 17: 2306. https://doi.org/10.3390/plants11172306