
The Effect of Ginger and Its Sub-Components on Pain

Abstract
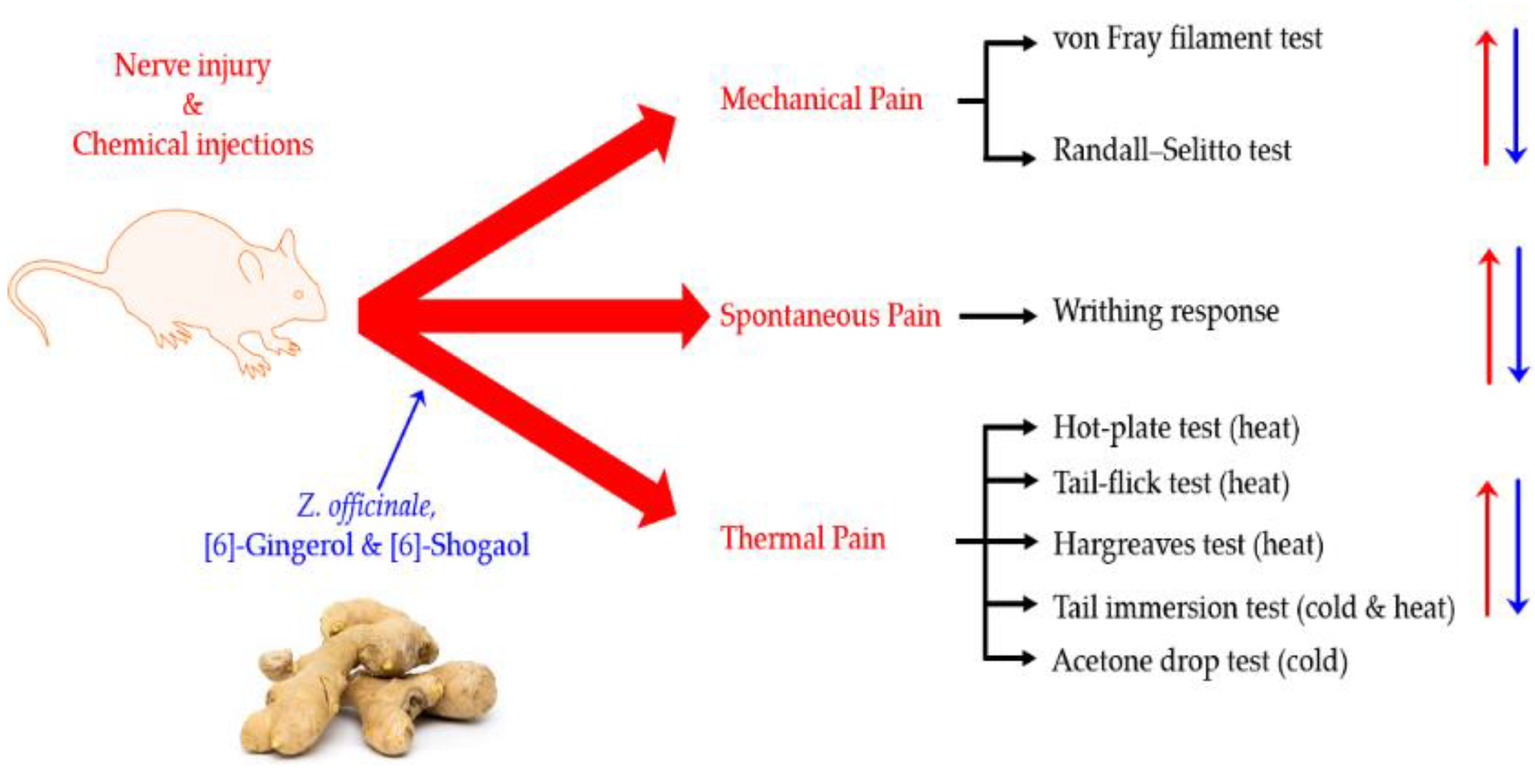
:1. Introduction
2. Results
2.1. Mechanical Pain
2.2. Spontaneous Pain
2.3. Thermal Pain
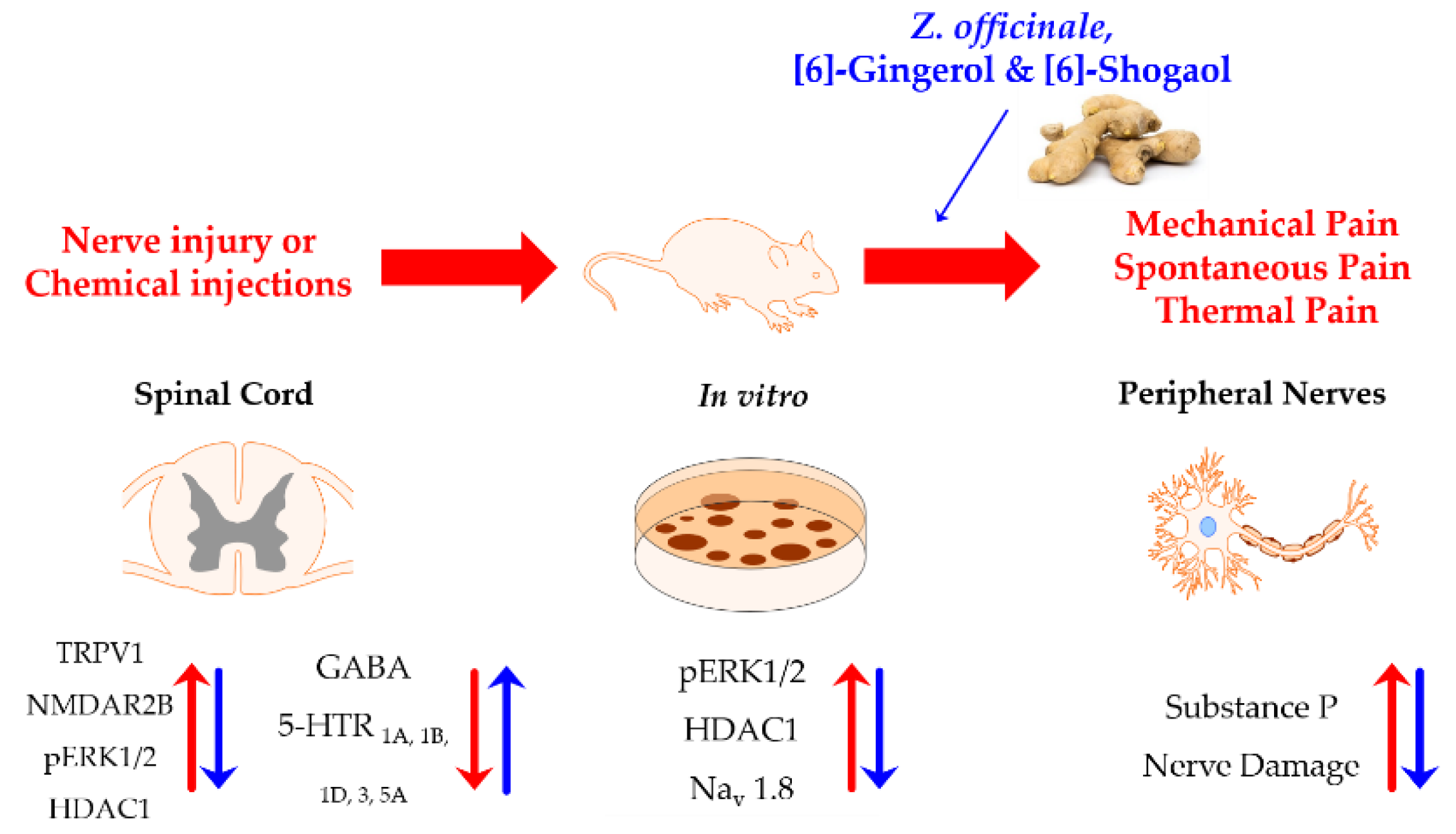
3. Discussion
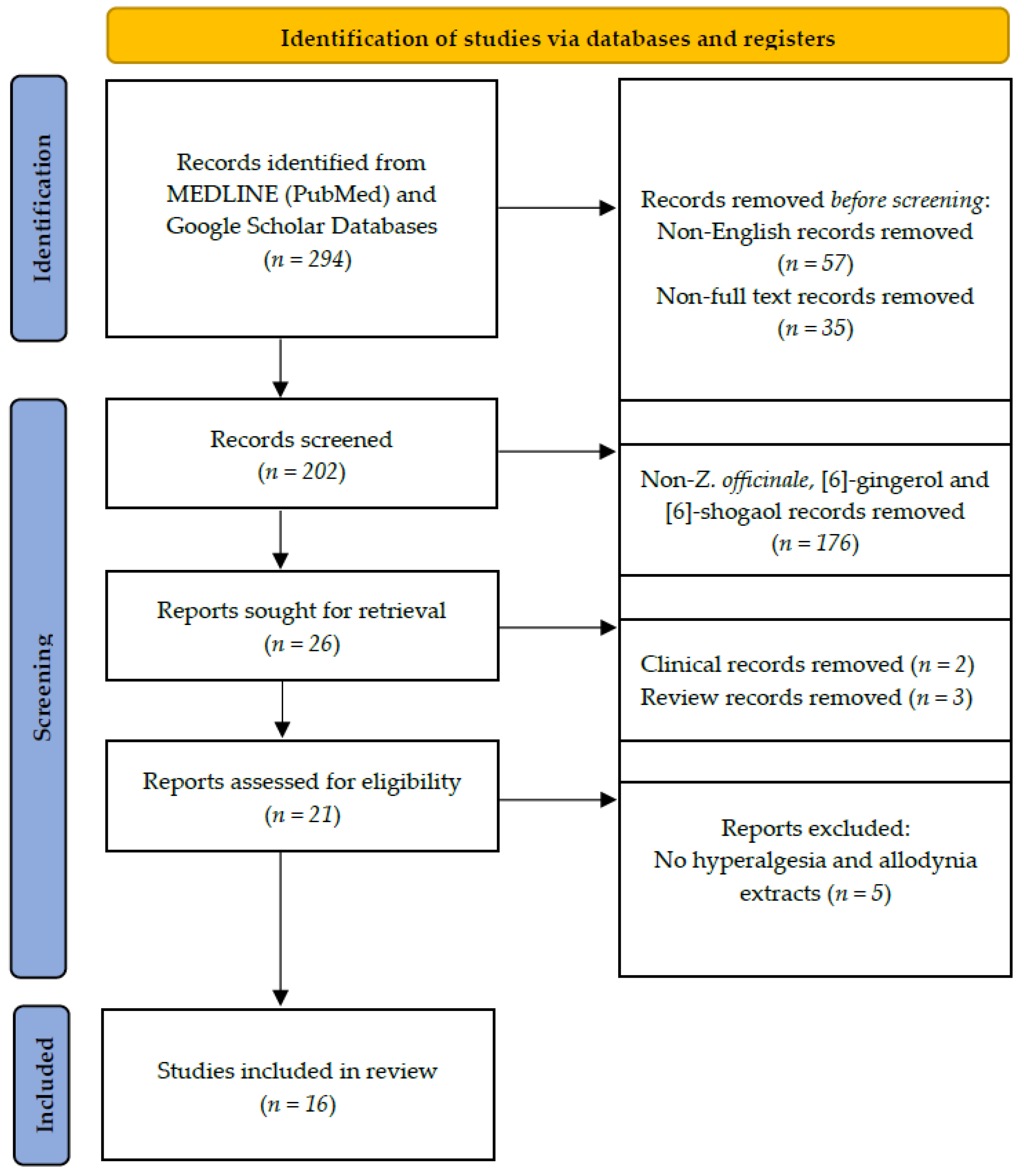
4. Materials and Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- White, B. Ginger: An overview. Am. Fam. Phys. 2007, 75, 1689–1691. [Google Scholar] [PubMed]
- Mao, Q.-Q.; Xu, X.-Y.; Cao, S.-Y.; Gan, R.-Y.; Corke, H.; Beta, T.; Li, H.-B. Bioactive compounds and bioactivities of ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, J.; Zhang, Y. Research progress on chemical constituents of Zingiber officinale Roscoe. BioMed Res. Int. 2019, 2019, 5370823. [Google Scholar] [CrossRef] [PubMed]
- Wohlmuth, H.; Leach, D.N.; Smith, M.K.; Myers, S.P. Gingerol content of diploid and tetraploid clones of ginger (Zingiber officinale Roscoe). J. Agric. Food Chem. 2005, 53, 5772–5778. [Google Scholar] [CrossRef]
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A. The revised IASP definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976. [Google Scholar] [CrossRef]
- Lavand’homme, P. The progression from acute to chronic pain. Curr. Opin. Anesthesiol. 2011, 24, 545–550. [Google Scholar] [CrossRef]
- Colloca, L.; Ludman, T.; Bouhassira, D.; Baron, R.; Dickenson, A.H.; Yarnitsky, D.; Freeman, R.; Truini, A.; Attal, N.; Finnerup, N.B. Neuropathic pain. Nat. Rev. Dis. Primers 2017, 3, 1–19. [Google Scholar] [CrossRef]
- Kidd, B.; Urban, L. Mechanisms of inflammatory pain. Br. J. Anaesth. 2001, 87, 3–11. [Google Scholar] [CrossRef]
- Bruera, E.; Kim, H.N. Cancer pain. JAMA 2003, 290, 2476–2479. [Google Scholar] [CrossRef]
- Levy, B.; Paulozzi, L.; Mack, K.A.; Jones, C.M. Trends in opioid analgesic–prescribing rates by specialty, US, 2007–2012. Am. J. Prev. Med. 2015, 49, 409–413. [Google Scholar] [CrossRef]
- Bedson, J.; Belcher, J.; Martino, O.; Ndlovu, M.; Rathod, T.; Walters, K.; Dunn, K.; Jordan, K. The effectiveness of national guidance in changing analgesic prescribing in primary care from 2002 to 2009: An observational database study. Eur. J. Pain 2013, 17, 434–443. [Google Scholar] [CrossRef]
- Brennan, M.J. The effect of opioid therapy on endocrine function. Am. J. Med. 2013, 126, S12–S18. [Google Scholar] [CrossRef]
- Martin, W.; Wikler, A.; Eades, C.; Pescor, F. Tolerance to and physical dependence on morphine in rats. Psychopharmacologia 1963, 4, 247–260. [Google Scholar] [CrossRef]
- Castellsague, J.; Riera-Guardia, N.; Calingaert, B.; Varas-Lorenzo, C.; Fourrier-Reglat, A.; Nicotra, F.; Sturkenboom, M.; Perez-Gutthann, S. Individual NSAIDs and upper gastrointestinal complications. Drug Saf. 2012, 35, 1127–1146. [Google Scholar] [CrossRef]
- Wang, X.; Wu, G.; Niu, T. Chemistry, pharmacology and application of ginger. Chin. Fruit Veg. 2016, 36, 23–29. [Google Scholar]
- Singletary, K. Ginger: An overview of health benefits. Nutr. Today 2010, 45, 171–183. [Google Scholar] [CrossRef]
- Baliga, M.S.; Haniadka, R.; Pereira, M.M.; D’Souza, J.J.; Pallaty, P.L.; Bhat, H.P.; Popuri, S. Update on the chemopreventive effects of ginger and its phytochemicals. Crit. Rev. Food Sci. Nutr. 2011, 51, 499–523. [Google Scholar] [CrossRef]
- Zick, S.M.; Djuric, Z.; Ruffin, M.T.; Litzinger, A.J.; Normolle, D.P.; Alrawi, S.; Feng, M.R.; Brenner, D.E. Pharmacokinetics of 6-gingerol, 8-gingerol, 10-gingerol, and 6-shogaol and conjugate metabolites in healthy human subjects. Cancer Epidemiol. Prev. Biomark. 2008, 17, 1930–1936. [Google Scholar] [CrossRef]
- Ojewole, J.A. Analgesic, antiinflammatory and hypoglycaemic effects of ethanol extract of Zingiber officinale (Roscoe) rhizomes (Zingiberaceae) in mice and rats. Phytother. Res. An. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2006, 20, 764–772. [Google Scholar]
- Habib, S.H.M.; Makpol, S.; Hamid, N.A.A.; Das, S.; Ngah, W.Z.W.; Yusof, Y.A.M. Ginger extract (Zingiber officinale) has anti-cancer and anti-inflammatory effects on ethionine-induced hepatoma rats. Clinics 2008, 63, 807–813. [Google Scholar] [CrossRef]
- Li, Y.; Tran, V.H.; Duke, C.C.; Roufogalis, B.D. Preventive and protective properties of Zingiber officinale (ginger) in diabetes mellitus, diabetic complications, and associated lipid and other metabolic disorders: A brief review. Evid.-Based Complement. Altern. Med. 2012, 2012, 516870. [Google Scholar] [CrossRef]
- Kamal, R.; Aleem, S. Clinical evaluation of the efficacy of a combination of zanjabeel (Zingiber officinale) and amla (Emblica officinalis) in hyperlipidaemia. Indian J. Tradit. Knowl. 2009, 8, 413–416. [Google Scholar]
- Ramakrishnan, R. Anticancer properties of Zingiber officinale–Ginger: A review. Int. J. Med. Pharm. Sci. 2013, 3, 11–20. [Google Scholar]
- Saneei Totmaj, A.; Emamat, H.; Jarrahi, F.; Zarrati, M. The effect of ginger (Zingiber officinale) on chemotherapy-induced nausea and vomiting in breast cancer patients: A systematic literature review of randomized controlled trials. Phytother Res. 2019, 33, 1957–1965. [Google Scholar] [CrossRef]
- Martins, L.B.; Rodrigues, A.M.d.S.; Rodrigues, D.F.; Dos Santos, L.C.; Teixeira, A.L.; Ferreira, A.V.M. Double-blind placebo-controlled randomized clinical trial of ginger (Zingiber officinale Rosc.) addition in migraine acute treatment. Cephalalgia 2019, 39, 68–76. [Google Scholar] [CrossRef]
- Daily, J.W.; Zhang, X.; Kim, D.S.; Park, S. Efficacy of ginger for alleviating the symptoms of primary dysmenorrhea: A systematic review and meta-analysis of randomized clinical trials. Pain Med. 2015, 16, 2243–2255. [Google Scholar] [CrossRef]
- Leach, M.J.; Kumar, S. The clinical effectiveness of Ginger (Zingiber officinale) in adults with osteoarthritis. Int. J. Evid.-Based Healthc. 2008, 6, 311–320. [Google Scholar] [PubMed]
- Lee, J.H.; Kim, B.; Ko, S.-G.; Kim, W. Analgesic Effect of SH003 and Trichosanthes kirilowii Maximowicz in Paclitaxel-Induced Neuropathic Pain in Mice. Curr. Issues Mol. Biol. 2022, 44, 718–730. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Ji, H.; Ko, S.-G.; Kim, W. Ji017 attenuates oxaliplatin-induced cold allodynia via spinal trpv1 and astrocytes inhibition in mice. Int. J. Mol. Sci. 2021, 22, 8811. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.; Kim, W. Effect of Acupuncture on Diabetic Neuropathy: A Narrative Review. Int. J. Mol. Sci. 2021, 22, 8575. [Google Scholar] [CrossRef]
- Lee, J.H.; Min, D.; Lee, D.; Kim, W. Zingiber officinale roscoe rhizomes attenuate oxaliplatin-induced neuropathic pain in mice. Molecules 2021, 26, 548. [Google Scholar] [CrossRef]
- Raji, Y.; Udoh, U.; Oluwadara, O.; Akinsomisoye, O.; Awobajo, O.; Adeshoga, K. Anti-flammatory and analgesic properties of the rhizome extract of Zingiber officinale. Afr. J. Biomed. Res. 2002, 5, 121–124. [Google Scholar] [CrossRef]
- Sepahvand, R.; Esmaeili-Mahani, S.; Arzi, A.; Rasoulian, B.; Abbasnejad, M. Ginger (Zingiber officinale Roscoe) elicits antinociceptive properties and potentiates morphine-induced analgesia in the rat radiant heat tail-flick test. J. Med. Food 2010, 13, 1397–1401. [Google Scholar] [CrossRef]
- Darvishzadeh-Mahani, F.; Esmaeili-Mahani, S.; Komeili, G.; Sheibani, V.; Zare, L. Ginger (Zingiber officinale Roscoe) prevents the development of morphine analgesic tolerance and physical dependence in rats. J. Ethnopharmacol. 2012, 141, 901–907. [Google Scholar] [CrossRef]
- Montserrat-de la Paz, S.; Garcia-Gimenez, M.D.; Quilez, A.M.; De la Puerta, R.; Fernandez-Arche, A. Ginger rhizome enhances the anti-inflammatory and anti-nociceptive effects of paracetamol in an experimental mouse model of fibromyalgia. Inflammopharmacology 2018, 26, 1093–1101. [Google Scholar] [CrossRef]
- Fajrin, F.A.; Purwandhono, A.; Christianty, F.M.; Sulistyaningrum, G.D.; Afifah, A.; Imandasari, N.; Barki, T. Antihyperalgesia potency of Zingiber officinale var. Rubrum in inflammatory and neuropathy-induced chronic pain condition in mice. Pak. J. Pharm. Sci. 2019, 32, 1663–1669. [Google Scholar]
- Kravchenko, I.; Eberle, L.; Nesterkina, M.; Kobernik, A. Anti-inflammatory and analgesic activity of ointment based on dense ginger extract (Zingiber officinale). J. Herbmed. Pharmacol. 2019, 8, 126–132. [Google Scholar] [CrossRef]
- Borgonetti, V.; Governa, P.; Biagi, M.; Pellati, F.; Galeotti, N. Zingiber officinale Roscoe rhizome extract alleviates neuropathic pain by inhibiting neuroinflammation in mice. Phytomedicine 2020, 78, 153307. [Google Scholar] [CrossRef]
- Young, H.-Y.; Luo, Y.-L.; Cheng, H.-Y.; Hsieh, W.-C.; Liao, J.-C.; Peng, W.-H. Analgesic and anti-inflammatory activities of [6]-gingerol. J. Ethnopharmacol. 2005, 96, 207–210. [Google Scholar] [CrossRef]
- Gauthier, M.L.; Beaudry, F.; Vachon, P. Intrathecal [6]-gingerol administration alleviates peripherally induced neuropathic pain in male Sprague–Dawley rats. Phytother. Res. 2013, 27, 1251–1254. [Google Scholar] [CrossRef]
- Hitomi, S.; Ono, K.; Terawaki, K.; Matsumoto, C.; Mizuno, K.; Yamaguchi, K.; Imai, R.; Omiya, Y.; Hattori, T.; Kase, Y. [6]-gingerol and [6]-shogaol, active ingredients of the traditional Japanese medicine hangeshashinto, relief oral ulcerative mucositis-induced pain via action on Na+ channels. Pharmacol. Res. 2017, 117, 288–302. [Google Scholar] [CrossRef]
- Mata-Bermudez, A.; Izquierdo, T.; de Los Monteros-Zuñiga, E.; Coen, A.; Godínez-Chaparro, B. Antiallodynic effect induced by [6]-gingerol in neuropathic rats is mediated by activation of the serotoninergic system and the nitric oxide-cyclic guanosine monophosphate-adenosine triphosphate-sensitive K(+) channel pathway. Phytother. Res. 2018, 32, 2520–2530. [Google Scholar] [CrossRef] [PubMed]
- Fajrin, F.A.; Nurrochmad, A.; Nugroho, A.E.; Susilowati, R. The improvement of pain behavior and sciatic nerves morphology in mice model of painful diabetic neuropathy upon administration of ginger (Zingiber officinale roscoe.) extract and its pungent compound, 6-shogaol. J. Nat. Sci. Biol. Med. 2019, 10, 149. [Google Scholar]
- Fajrin, F.A.; Nugroho, A.E.; Nurrochmad, A.; Susilowati, R. Ginger extract and its compound, 6-shogaol, attenuates painful diabetic neuropathy in mice via reducing TRPV1 and NMDAR2B expressions in the spinal cord. J. Ethnopharmacol. 2020, 249, 112396. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Gang, J.; Lee, J.-H.; Yang, H.; Cheon, C.; Ko, S.-G.; Bae, H.; Kim, W. [6]-Shogaol Attenuates Oxaliplatin-Induced Allodynia through Serotonergic Receptors and GABA in the Spinal Cord in Mice. Pharmaceuticals 2022, 15, 726. [Google Scholar] [CrossRef] [PubMed]
- Marzvanyan, A.; Alhawaj, A.F. Physiology, Sensory Receptors; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Council, N.R. Definition of Pain and Distress and Reporting Requirements for Laboratory Animals: Proceedings of the Workshop held June 22, 2000; National Academies Press: Washington, DC, USA, 2000; Volume 132, p. NBK99547. [Google Scholar]
- Pinho-Ribeiro, F.A.; Verri, W.A., Jr.; Chiu, I.M. Nociceptor sensory neuron–immune interactions in pain and inflammation. Trends Immunol. 2017, 38, 5–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef]
- Dib-Hajj, S.D.; Yang, Y.; Waxman, S.G. Genetics and molecular pathophysiology of Nav1. 7-related pain syndromes. Adv. Genet. 2008, 63, 85–110. [Google Scholar]
- Kim, Y.H.; Back, S.K.; Davies, A.J.; Jeong, H.; Jo, H.J.; Chung, G.; Na, H.S.; Bae, Y.C.; Kim, S.J.; Kim, J.S. TRPV1 in GABAergic interneurons mediates neuropathic mechanical allodynia and disinhibition of the nociceptive circuitry in the spinal cord. Neuron 2012, 74, 640–647. [Google Scholar] [CrossRef]
- Alloui, A.; Zimmermann, K.; Mamet, J.; Duprat, F.; Noel, J.; Chemin, J.; Guy, N.; Blondeau, N.; Voilley, N.; Rubat-Coudert, C. TREK-1, a K+ channel involved in polymodal pain perception. EMBO J. 2006, 25, 2368–2376. [Google Scholar] [CrossRef]
- Bennett, G.J.; Xie, Y.-K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Suter, M.; Siegenthaler, A.; Decosterd, I.; Ji, R. Perioperative nerve blockade: Clues from the bench. Anesthesiol. Res. Pract. 2011, 2011, 124898. [Google Scholar] [CrossRef]
- Li, Y.; Dorsi, M.J.; Meyer, R.A.; Belzberg, A.J. Mechanical hyperalgesia after an L5 spinal nerve lesion in the rat is not dependent on input from injured nerve fibers. Pain 2000, 85, 493–502. [Google Scholar] [CrossRef]
- Yoon, C.; Wook, Y.Y.; Sik, N.H.; Ho, K.S.; Mo, C.J. Behavioral signs of ongoing pain and cold allodynia in a rat model of neuropathic pain. Pain 1994, 59, 369–376. [Google Scholar] [CrossRef]
- Honoré, P.H.; Basnet, A.; Eljaja, L.; Kristensen, P.; Andersen, L.M.; Neustrup, S.; Møllgaard, P.; Bjerrum, O.J. Neuropathic pain models in the development of analgesic drugs. Scand. J. Pain 2011, 2, 172–177. [Google Scholar] [CrossRef]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Malmberg, A.B.; Basbaum, A.I. Partial sciatic nerve injury in the mouse as a model of neuropathic pain: Behavioral and neuroanatomical correlates. Pain 1998, 76, 215–222. [Google Scholar] [CrossRef]
- Jensen, T.S.; Gottrup, H.; Sindrup, S.H.; Bach, F.W. The clinical picture of neuropathic pain. Eur. J. Pharmacol. 2001, 429, 1–11. [Google Scholar] [CrossRef]
- Djouhri, L.; Koutsikou, S.; Fang, X.; McMullan, S.; Lawson, S.N. Spontaneous pain, both neuropathic and inflammatory, is related to frequency of spontaneous firing in intact C-fiber nociceptors. J. Neurosci. 2006, 26, 1281–1292. [Google Scholar] [CrossRef]
- Finnerup, N.B.; Kuner, R.; Jensen, T.S. Neuropathic pain: From mechanisms to treatment. Physiol. Rev. 2021, 101, 259–301. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.J. What is spontaneous pain and who has it? J. Pain 2012, 13, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, S.; Nakagawa, T.; Yamauchi, Y.; Minami, M.; Satoh, M. Differential contributions of the basolateral and central nuclei of the amygdala in the negative affective component of chemical somatic and visceral pains in rats. Eur. J. Neurosci. 2003, 18, 2343–2350. [Google Scholar] [CrossRef] [PubMed]
- Tjølsen, A.; Berge, O.-G.; Hunskaar, S.; Rosland, J.H.; Hole, K. The formalin test: An evaluation of the method. Pain 1992, 51, 5–17. [Google Scholar] [CrossRef]
- Thermal Hyperalgesia. In Encyclopedia of Pain; Gebhart, G.F.; Schmidt, R.F. (Eds.) Springer: Heidelberg/Berlin, Germany, 2013; p. 3948. [Google Scholar]
- Zhang, X.; Li, L.; McNaughton, P.A. Proinflammatory mediators modulate the heat-activated ion channel TRPV1 via the scaffolding protein AKAP79/150. Neuron 2008, 59, 450–461. [Google Scholar] [CrossRef]
- Jensen, T.S.; Finnerup, N.B. Allodynia and hyperalgesia in neuropathic pain: Clinical manifestations and mechanisms. Lancet Neurol. 2014, 13, 924–935. [Google Scholar] [CrossRef]
- Defrin, R.; Ohry, A.; Blumen, N.; Urca, G. Sensory determinants of thermal pain. Brain 2002, 125, 501–510. [Google Scholar] [CrossRef]
- Campbell, J.N.; LaMotte, R.H. Latency to detection of first pain. Brain Res. 1983, 266, 203–208. [Google Scholar] [CrossRef]
- Treede, R.; Meyer, R.; Raja, S.N.; Campbell, J.N. Evidence for two different heat transduction mechanisms in nociceptive primary afferents innervating monkey skin. J. Physiol. 1995, 483, 747–758. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Viana, F. Nociceptors: Thermal allodynia and thermal pain. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 156, pp. 103–119. [Google Scholar]
- Bridges, D.; Thompson, S.; Rice, A. Mechanisms of neuropathic pain. Br. J. Anaesth. 2001, 87, 12–26. [Google Scholar] [CrossRef]
- Aley, K.; Reichling, D.; Levine, J. Vincristine hyperalgesia in the rat: A model of painful vincristine neuropathy in humans. Neuroscience 1996, 73, 259–265. [Google Scholar] [CrossRef]
- Courteix, C.; Eschalier, A.; Lavarenne, J. Streptozocin-induced diabetic rats: Behavioural evidence for a model of chronic pain. Pain 1993, 53, 81–88. [Google Scholar] [CrossRef]
- Haniadka, R.; Saldanha, E.; Sunita, V.; Palatty, P.L.; Fayad, R.; Baliga, M.S. A review of the gastroprotective effects of ginger (Zingiber officinale Roscoe). Food Funct. 2013, 4, 845–855. [Google Scholar] [CrossRef]
- Nikkhah-Bodaghi, M.; Maleki, I.; Agah, S.; Hekmatdoost, A. Zingiber officinale and oxidative stress in patients with ulcerative colitis: A randomized, placebo-controlled, clinical trial. Complement. Med. 2019, 43, 1–6. [Google Scholar] [CrossRef]
- Sharifzadeh, F.; Kashanian, M.; Koohpayehzadeh, J.; Rezaian, F.; Sheikhansari, N.; Eshraghi, N. A comparison between the effects of ginger, pyridoxine (vitamin B6) and placebo for the treatment of the first trimester nausea and vomiting of pregnancy (NVP). J. Matern.-Fetal Neonatal Med. 2018, 31, 2509–2514. [Google Scholar] [CrossRef] [PubMed]
- Terry, R.; Posadzki, P.; Watson, L.K.; Ernst, E. The use of ginger (Zingiber officinale) for the treatment of pain: A systematic review of clinical trials. Pain Med. 2011, 12, 1808–1818. [Google Scholar] [CrossRef]
- Pagano, E.; Souto, E.B.; Durazzo, A.; Sharifi-Rad, J.; Lucarini, M.; Souto, S.B.; Salehi, B.; Zam, W.; Montanaro, V.; Lucariello, G. Ginger (Zingiber officinale Roscoe) as a nutraceutical: Focus on the metabolic, analgesic, and antiinflammatory effects. Phytother. Res. 2021, 35, 2403–2417. [Google Scholar] [CrossRef]
- Lakhan, S.E.; Ford, C.T.; Tepper, D. Zingiberaceae extracts for pain: A systematic review and meta-analysis. Nutr. J. 2015, 14, 1–10. [Google Scholar] [CrossRef]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Viljoen, A.M. Gingerols and shogaols: Important nutraceutical principles from ginger. Phytochemistry 2015, 117, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.; Elgharib, M.; El-awady, R.; Saleh, E. Ginger: From serving table to salient therapy. Food Biosci. 2021, 41, 100934. [Google Scholar] [CrossRef]
- Williamson, E.M.; Liu, X.; Izzo, A.A. Trends in use, pharmacology, and clinical applications of emerging herbal nutraceuticals. Br. J. Pharmacol. 2020, 177, 1227–1240. [Google Scholar] [CrossRef]
- Shalaby, M.; Hamowieh, A. Safety and efficacy of Zingiber officinale roots on fertility of male diabetic rats. Food Chem. Toxicol. 2010, 48, 2920–2924. [Google Scholar] [CrossRef]
- Purseglove, J.W.; Brown, E.; Green, C.; Robbins, S. Spices Vol. 2; Longman Group Ltd.: London, UK, 1981. [Google Scholar]
- Prasad, S.; Tyagi, A.K. Ginger and its constituents: Role in prevention and treatment of gastrointestinal cancer. Gastroenterol. Res. Pract. 2015, 2015, 142979. [Google Scholar] [CrossRef]
- Ok, S.; Jeong, W.-S. Optimization of extraction conditions for the 6-shogaol-rich extract from ginger (Zingiber officinale Roscoe). Prev. Nutr. Food Sci. 2012, 17, 166. [Google Scholar] [CrossRef]
- Chen, C.C.; Kuo, M.C.; Wu, C.M.; Ho, C.T. Pungent compounds of ginger (Zingiber officinale Roscoe) extracted by liquid carbon dioxide. J. Agric. Food Chem. 1986, 34, 477–480. [Google Scholar] [CrossRef]
- Simon, A.; Darcsi, A.; Kéry, Á.; Riethmüller, E. Blood-brain barrier permeability study of ginger constituents. J. Pharm. Biomed. Anal. 2020, 177, 112820. [Google Scholar] [CrossRef]
- Hannon, J.; Hoyer, D. Molecular biology of 5-HT receptors. Behav. Brain Res. 2008, 195, 198–213. [Google Scholar] [CrossRef]
- Hoyer, D.; Clarke, D.E.; Fozard, J.R.; Hartig, P.R.; Martin, G.R.; Mylecharane, E.J.; Saxena, P.R.; Humphrey, P. International Union of Pharmacology classification of receptors for 5-hydroxytryptamine (Serotonin). Pharmacol. Rev. 1994, 46, 157–203. [Google Scholar]
- Avila-Rojas, S.H.; Velazquez-Lagunas, I.; Salinas-Abarca, A.B.; Barragan-Iglesias, P.; Pineda-Farias, J.B.; Granados-Soto, V. Role of spinal 5-HT5A, and 5-HT1A/1B/1D, receptors in neuropathic pain induced by spinal nerve ligation in rats. Brain Res. 2015, 1622, 377–385. [Google Scholar] [CrossRef]
- Cortes-Altamirano, J.L.; Olmos-Hernandez, A.; Jaime, H.B.; Carrillo-Mora, P.; Bandala, C.; Reyes-Long, S.; Alfaro-Rodríguez, A. 5-HT1, 5-HT2, 5-HT3 and 5-HT7 receptors and their role in the modulation of pain response in the central nervous system. Curr. Neuropharmacol. 2018, 16, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Bardin, L. The complex role of serotonin and 5-HT receptors in chronic pain. Behav. Pharmacol. 2011, 22, 390–404. [Google Scholar] [CrossRef] [PubMed]
- Morera, E.; De Petrocellis, L.; Morera, L.; Moriello, A.S.; Nalli, M.; Di Marzo, V.; Ortar, G. Synthesis and biological evaluation of [6]-gingerol analogues as transient receptor potential channel TRPV1 and TRPA1 modulators. Bioorg. Med. Chem. Lett. 2012, 22, 1674–1677. [Google Scholar] [CrossRef] [PubMed]
- Fajrin, F.A.; Nugroho, A.E.; Nurrochmad, A.; Susilowati, R. Molecular docking analysis of ginger active compound on transient receptor potential cation channel subfamily V member 1 (TRPV1). Indones. J. Chem. 2018, 18, 179–185. [Google Scholar] [CrossRef]
- Bishnoi, M.; Bosgraaf, C.A.; Abooj, u.; Zhong, L.; Premkumar, L.S. Streptozotocin-induced early thermal hyperalgesia is independent of glycemic state of rats: Role of transient receptor potential vanilloid 1 (TRPV1) and inflammatory mediators. Mol. Pain 2011, 7, 1744–8069. [Google Scholar] [CrossRef]
- Kamata, Y.; Kambe, T.; Chiba, T.; Yamamoto, K.; Kawakami, K.; Abe, K.; Taguchi, K. Paclitaxel induces upregulation of transient receptor potential vanilloid 1 expression in the rat spinal cord. Int. J. Mol. Sci. 2020, 21, 4341. [Google Scholar] [CrossRef]
- Valtschanoff, J.G.; Rustioni, A.; Guo, A.; Hwang, S.J. Vanilloid receptor VR1 is both presynaptic and postsynaptic in the superficial laminae of the rat dorsal horn. J. Comp. Neurol. 2001, 436, 225–235. [Google Scholar] [CrossRef]
- Kanai, Y.; Nakazato, E.; Fujiuchi, A.; Hara, T.; Imai, A. Involvement of an increased spinal TRPV1 sensitization through its up-regulation in mechanical allodynia of CCI rats. Neuropharmacology 2005, 49, 977–984. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, Q.; Crook, R.J.; O’Neil, R.G.; Walters, E.T. TRPV1 channels make major contributions to behavioral hypersensitivity and spontaneous activity in nociceptors after spinal cord injury. Pain 2013, 154, 2130–2141. [Google Scholar] [CrossRef]
- Arendt-Nielsen, L.; Harris, S.; Whiteside, G.T.; Hummel, M.; Knappenberger, T.; O’Keefe, S.; Kapil, R.; Kyle, D. A randomized, double-blind, positive-controlled, 3-way cross-over human experimental pain study of a TRPV1 antagonist (V116517) in healthy volunteers and comparison with preclinical profile. Pain 2016, 157, 2057–2067. [Google Scholar] [CrossRef]
- Chen, Y.; Willcockson, H.H.; Valtschanoff, J.G. Influence of the vanilloid receptor TRPV1 on the activation of spinal cord glia in mouse models of pain. Exp. Neurol. 2009, 220, 383–390. [Google Scholar] [CrossRef]
- Palazzo, E.; Luongo, L.; de Novellis, V.; Berrino, L.; Rossi, F.; Maione, S. Moving towards supraspinal TRPV1 receptors for chronic pain relief. Mol. Pain 2010, 6, 1–11. [Google Scholar] [CrossRef]
- Lee, J.; Saloman, J.L.; Weiland, G.; Auh, Q.-S.; Chung, M.-K.; Ro, J.Y. Functional interactions between NMDA receptors and TRPV1 in trigeminal sensory neurons mediate mechanical hyperalgesia in the rat masseter muscle. Pain 2012, 153, 1514–1524. [Google Scholar] [CrossRef]
- Kang, S.-Y.; Seo, S.Y.; Bang, S.K.; Cho, S.J.; Choi, K.-H.; Ryu, Y. Inhibition of spinal TRPV1 reduces NMDA receptor 2B phosphorylation and produces anti-nociceptive effects in mice with inflammatory pain. Int. J. Mol. Sci. 2021, 22, 11177. [Google Scholar] [CrossRef]
- Roskov, Y.; Kunze, T.; Paglinawan, L.; Orrell, T.; Nicolson, D.; Culham, A.; Bailly, N.; Kirk, P.; Bourgoin, T.; Baillargeon, G. Species 2000 & ITIS Catalogue of Life, 2013 Annual Checklist. 2013. Available online: http://www.catalogueoflife.org/annual-checklist/2013/ (accessed on 9 August 2022).
- Khalid, M.H.; Akhtar, M.N.; Mohamad, A.S.; Perimal, E.K.; Akira, A.; Israf, D.A.; Lajis, N.; Sulaiman, M.R. Antinociceptive effect of the essential oil of Zingiber zerumbet in mice: Possible mechanisms. J. Ethnopharmacol. 2011, 137, 345–351. [Google Scholar] [CrossRef]
- Lawson, K. Potassium channel activation: A potential therapeutic approach? Pharmacol. Ther. 1996, 70, 39–63. [Google Scholar] [CrossRef]
- Slavin, M.; Bourguignon, J.; Jackson, K.; Orciga, M.-A. Impact of food components on in vitro calcitonin gene-related peptide secretion—A potential mechanism for dietary influence on migraine. Nutrients 2016, 8, 406. [Google Scholar] [CrossRef]
- Durham, P.L.; Vause, C.V. Calcitonin gene-related peptide (CGRP) receptor antagonists in the treatment of migraine. CNS Drugs 2010, 24, 539–548. [Google Scholar] [CrossRef]
- Farajdokht, F.; Mohaddes, G.; Shanehbandi, D.; Karimi, P.; Babri, S. Ghrelin attenuated hyperalgesia induced by chronic nitroglycerin: CGRP and TRPV1 as targets for migraine management. Cephalalgia 2018, 38, 1716–1730. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Authors | Strain | Pain | Z. officinale Roscoe | Findings | ||
|---|---|---|---|---|---|---|
| Rats | ||||||
| Sepahvand et al., 2010 [33] | Wistar Rat | Tail-Flick Test | 200, 400 and 600 mg/kg (i.p. 80% Ethanol Extract) | Control: | Heat Pain | - |
| Z. officinale: | ↓ | |||||
| Z. officinale + Morphine (2.5 mg/kg): | ↓ | |||||
| Mechanism of Actions: | - | |||||
| Darvishzadeh-Mahani et al., 2012 [34] | Wistar Rat | Tail-Flick Test | 50 and 100 mg/kg (p.o. 96% Ethanol Extract) | Control: | Heat Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Mechanism of Actions: | - | |||||
| Mice | ||||||
| Y et al., 2002 [32] | Swiss Mice | Acetic Acid 3% (i.p.) | 50 and 100 mg/kg (i.p., 100% Ethanol Extract) | Control: | Spontaneous Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Aspirin (150 mg/kg, i.p.): | ↓ | |||||
| Mechanism of Actions: | - | |||||
| Ojewole 2006 [19] | Balb C Mice | Acetic Acid 3% (i.p.) and Hot Plate Test | 100, 200, 400 and 800 mg/kg (i.p. 96% Ethanol Extract) | Control: | Spontaneous and Heat Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Morphine (10 mg/kg, i.p.): Diclofenac (100 mg/kg, i.p.): | ↓ | |||||
| Mechanism of Actions: | - | |||||
| Montserrat-de la Paz et al., 2018 [35] | C57BL/6J Mice | ICS-induced FMS models | 0.5 and 1% (p.o. Mixed with Standard Diet) | Control: | Cold, Heat and Mechanical Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Z. officinale + Paracetamol: | ↓ | |||||
| Mechanism of Actions: | - | |||||
| Fajrin et al., 2019 [36] | Mice | CFA 40 μL (Intraplantar Injection) and PSNL | 100, 200, 400 and 600 mg/kg (p.o., Destilator with Aquadest) | Control: | Heat Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Mechanism of Actions: | - | |||||
| Kravchenko et al., 2019 [37] | White Mice | AITC 0.5% (Subplantar Injection) | 0.0125, 0.025, 0.05, 0.1, 0.5, 1 and 5% of Extract Ointment | Control: | Spontaneous Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Benzocaine (Ointment): | ↓ | |||||
| Mechanism of Actions: | - | |||||
| Fajrin et al., 2019 [43] | Balb/c Mice | STZ 110 mg/kg (i.p.) | 100, 200 and 400 mg/kg (p.o., 96% Ethanol Extract) | Control: | Heat and Mechanical Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Gabapetin (100 mg/kg, p.o.): | ↓ | |||||
| Mechanism of Actions: | Prevention of sciatic nerve damage | |||||
| Fajrin et al., 2020 [44] | Balb/c Mice | STZ 110 mg/kg (i.p.) | 100, 200 and 400 mg/kg (p.o., 96% Ethanol Extract) | Control: | Heat and Mechanical Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Gabapetin (100 mg/kg, p.o.): | ↓ | |||||
| Mechanism of Actions: | ↓ TRPV1 and NMDAR2B mRNA expression (spinal cord) | |||||
| Borgonetti et al., 2020 [38] | CD1 Mice | SNI | 200 and 400 mg/kg (p.o., Supercritical CO2 extraction) | Control: | Mechanical and Heat Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Pregabalin (30 mg/kg, p.o.): | ↓ | |||||
| Mechanism of Actions: | ↓ pERK1/2 activation (in BV2 cells and spinal cord) ↓ HDAC1 expression (in BV2 cells and spinal cord) | |||||
| Lee et al., 2021 [31] | C57BL/6 Mice | Oxaliplatin 6 mg/kg (i.p.) | 100, 300 and 500 mg/kg (p.o., 100% Water Extract) | Control: | Cold and Mechanical Pain | ↑ |
| Z. officinale: | ↓ | |||||
| Mechanism of Actions: | Analgesic Effect Blocked by Mixed 5-HT1 and 5-HT2 receptor, 5-HT1A and 5-HT3 antagonists’ injections (i.t.) ↑ mRNA expression level of 5-HT1A receptor | |||||
| Authors | Strain | Pain | Treatments | Findings | ||
|---|---|---|---|---|---|---|
| Rats | ||||||
| Gauthier et al., 2012 [40] | SD Rat | CCI | [6]-Gingerol 10 μg (i.t.) | Control: | Heat and Mechanical Pain | ↑ |
| [6]-Gingerol: | ↓ | |||||
| Cyclodextrin Formulation (20 μL, i.t.): | ↑ | |||||
| Mechanism of Action: | - | |||||
| Hitomi et al., 2017 [41] | Wistar Rat | OUM | [6]-Shogaol 150 μM [6]-Gingerol 300 μM (Swab Application) | Control: | Mechanical Pain | ↑ |
| [6]-Shogaol + [6]-Gingerol: | - | |||||
| Mechanism of Action: | ↓ Evoked currents on Nav1.8. (CHO cell) ↓ SP release (CHO cells) | |||||
| Mata-Bermudez et al., 2018 [42] | Wistar Rat | SNL | [6]-Gingerol 1, 3, 6 and 10 μg/rat (i.t.) | Control: | Mechanical Pain | ↑ |
| [6]-Gingerol: | ↓ | |||||
| Gabapentin (100 μg/rat, i.t.): | ↓ | |||||
| Mechanism of Action: | Effect not blocked by nonselective opioid receptor antagonist (naloxone, i.t.) Effect blocked by nonselective 5-HT, 5-HT1A, 1B, 1D, 5A receptor antagonists (methiothepin, WAY-100635, SB-224289, BRL-15572, SB-659551, i.t.) Effect blocked by nonselective NO synthase inhibitor, inhibitor of guanylate cyclase, channel blocker of ATP-sensitive K+ channels (L-NAME, ODQ, glibenclamide, i.t.) | |||||
| Mice | ||||||
| Young et al., 2005 [39] | ICR Mice | Acetic Acid 1% (i.p.) and 10% Formalin (s.c.) | [6]-Gingerol 25 and 50 mg/kg (i.p.) | Control: | Spontaneous Pain | ↑ |
| [6]-Gingerol: | ↓ | |||||
| Indomethacin (10 mg/kg, i.p.): | ↓ | |||||
| Mechanism of Action: | - | |||||
| Fajrin et al., 2019 [43] | Balb/c Mice | STZ 110 mg/kg (i.p.) | [6]-Shogaol 5, 10 and 15 mg/kg (p.o.) | Control: | Heat and Mechanical Pain | ↑ |
| [6]-Shogaol: | ↓ | |||||
| Gabapentin (100 mg/kg, p.o.): | ↓ | |||||
| Mechanism of Action: | Prevention of sciatic nerve damage | |||||
| Fajrin et al., 2020 [44] | Balb/c Mice | STZ 110 mg/kg (i.p.) | [6]-Shogaol 5, 10 and 15 mg/kg (p.o.) | Control | Heat and Mechanical Pain | ↑ |
| [6]-shogaol | ↓ | |||||
| Gabapentin (100 mg/kg, p.o.) | ↓ | |||||
| Mechanism of Action | ↓ TRPV1 and NMDAR2B mRNA expression (spinal cord) | |||||
| Kim et al., 2022 [45] | C57BL/6 Mice | Oxaliplatin 6 mg/kg (i.p.) | [6]-Shogaol 10 mg/kg (i.p.) | Control: | Cold and Mechanical pain: | ↑ |
| [6]-shogaol: | ↓ | |||||
| Mechanism of Action: | Effect blocked by 5-HT1A, 3 receptor antagonists (NAN-190, MDL-72222, i.t.) Effect blocked by GABAB receptor antagonist (CGP 55845, i.t.) ↑ GABA and GAD65 concentration (spinal cord) | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Cheon, C.; Kim, B.; Kim, W. The Effect of Ginger and Its Sub-Components on Pain. Plants 2022, 11, 2296. https://doi.org/10.3390/plants11172296
Kim S, Cheon C, Kim B, Kim W. The Effect of Ginger and Its Sub-Components on Pain. Plants. 2022; 11(17):2296. https://doi.org/10.3390/plants11172296
Chicago/Turabian StyleKim, Suyong, Chunhoo Cheon, Bonglee Kim, and Woojin Kim. 2022. "The Effect of Ginger and Its Sub-Components on Pain" Plants 11, no. 17: 2296. https://doi.org/10.3390/plants11172296