
In Vitro and In Silico Screening of Anti-Vibrio spp., Antibiofilm, Antioxidant and Anti-Quorum Sensing Activities of Cuminum cyminum L. Volatile Oil


Abstract
:1. Introduction
2. Results
2.1. Phytochemical Composition
2.2. Antioxidant Activities
2.3. Antimicrobial Activity
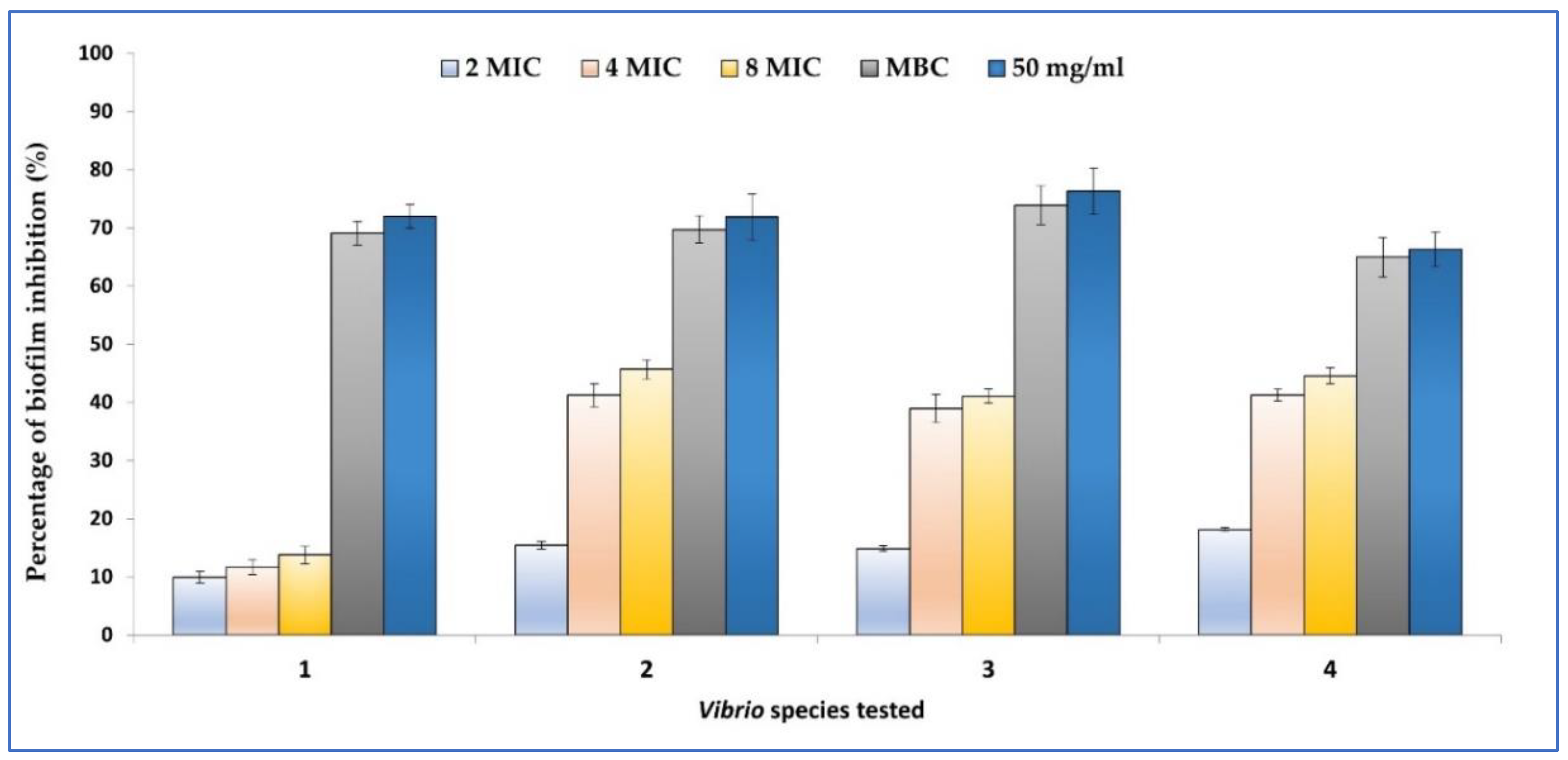
2.4. Biofilm Inhibition
2.5. Anti-QS Activity
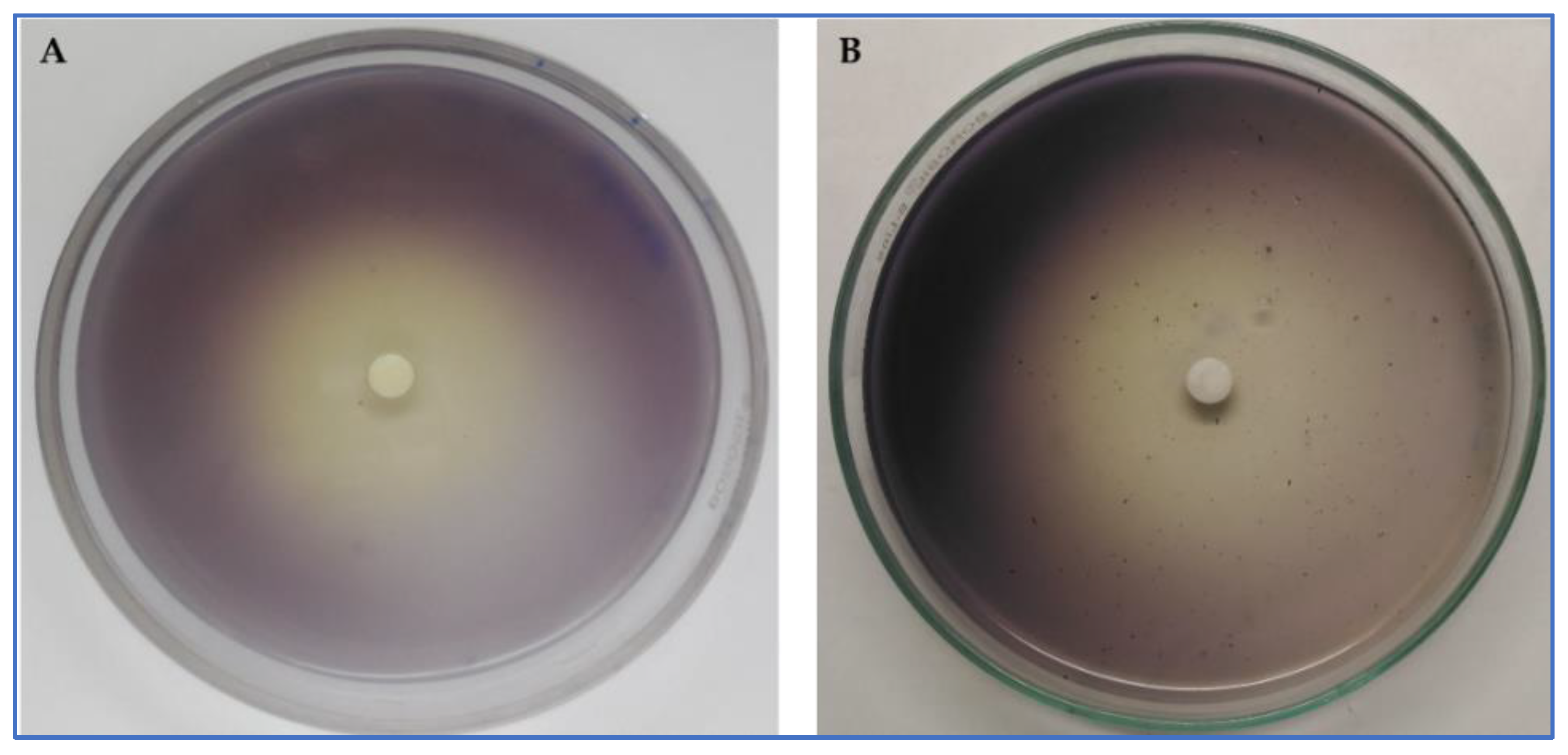
2.5.1. Qualitative and Quantitative Violacein Inhibition Estimation
2.5.2. Anti-Swarming Activity
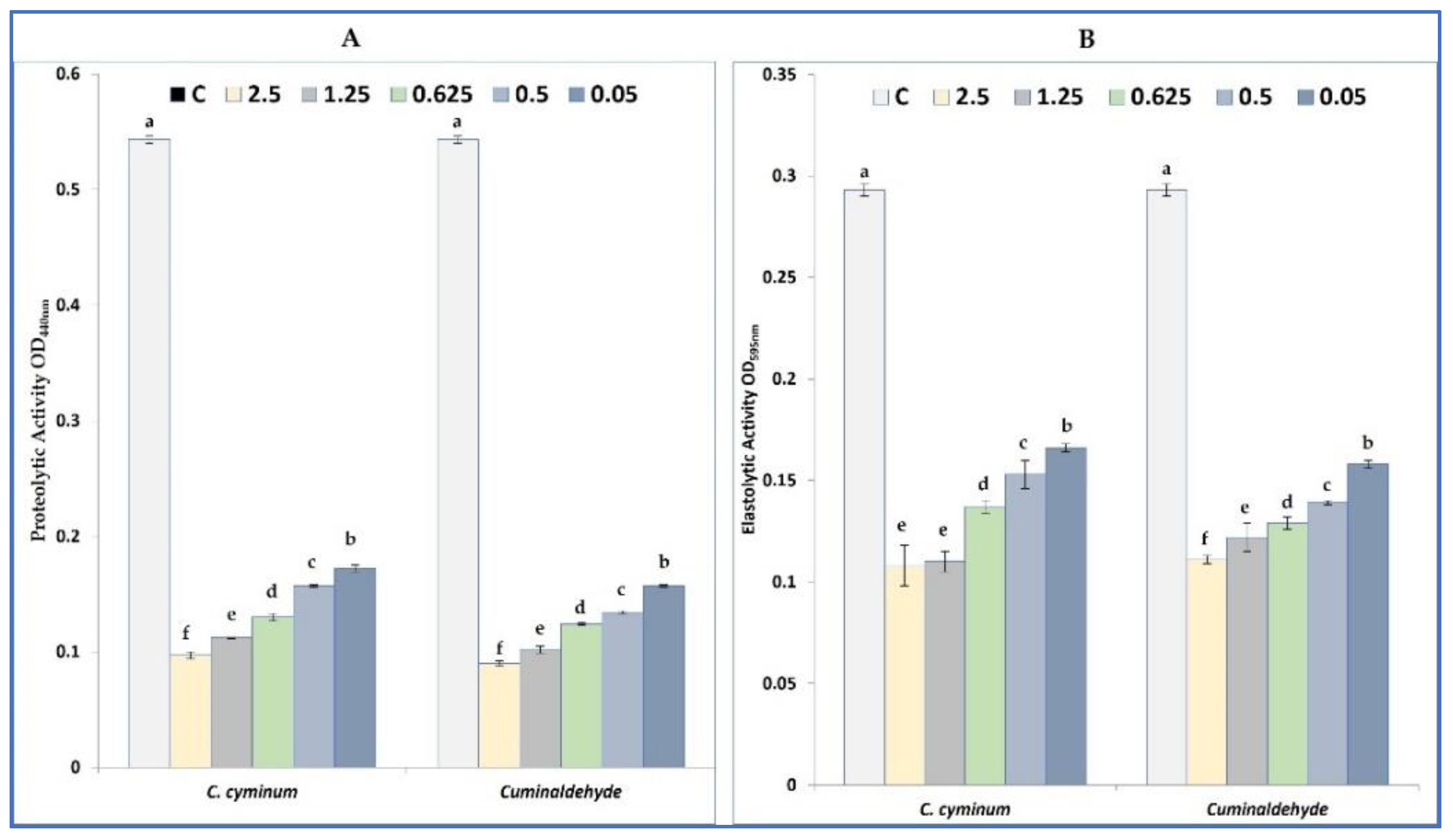
2.5.3. Elastase and Protease Inhibition
2.6. ADMET Analysis
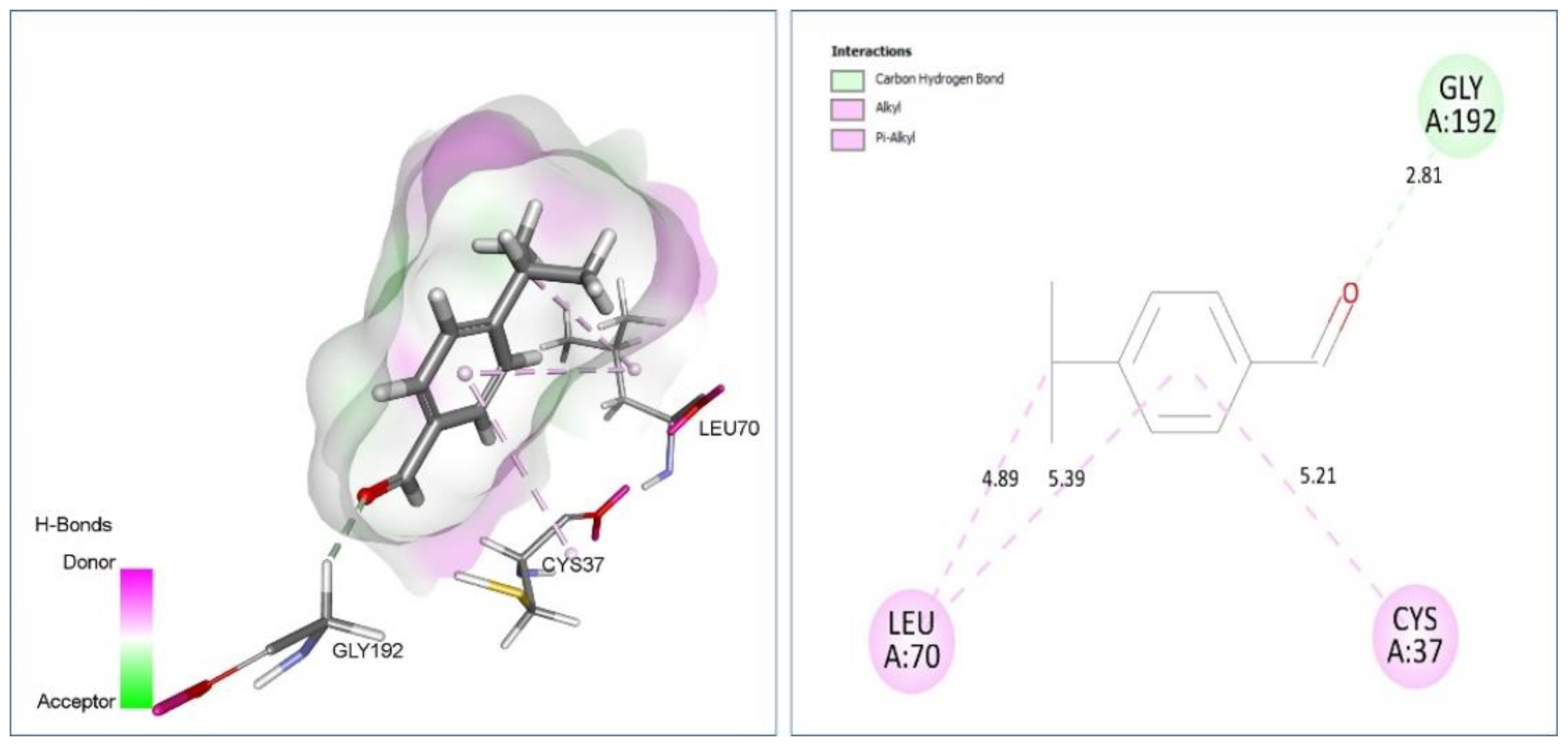
2.7. Molecular Docking Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction Procedure
4.2. Analysis of the Volatile Compounds
4.3. Sceening of the Anti-Vibrio spp. Activity
4.4. Evaluation of the Antioxidant Activities
4.5. Inhibition of Virulence Factors Regulated by QS System
4.5.1. Inhibition of Violacein
4.5.2. Biofilm Inhibition
4.5.3. Effect on Flagellar Motility
4.5.4. Elastase and Protease Inhibition in P. aeruginosa PAO1
4.6. Computational Approach
4.7. ADMET Predicted Properties
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dhingra, S.; Rahman, N.A.A.; Peile, E.; Rahman, M.; Sartelli, M.; Hassali, M.A.; Islam, T.; Islam, S.; Haque, M. Microbial Resistance Movements: An Overview of Global Public Health Threats Posed by Antimicrobial Resistance, and How Best to Counter. Front. Public Health 2020, 8, 535668. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.F. Fighting Back against Antimicrobial Resistance with Comprehensive Policy and Education: A Narrative Review. Antibiotics 2022, 11, 644. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Asghar, A.; Algburi, A.; Huang, Q.; Ahmad, T.; Zhong, H.; Javed, H.U.; Ermakov, A.M.; Chikindas, M.L. Anti-biofilm Potential of Elletaria cardamomum Essential Oil Against Escherichia coli O157:H7 and Salmonella Typhimurium JSG 1748. Front. Microbiol. 2021, 12, 620227. [Google Scholar]
- Singh, S.; Datta, S.; Narayanan, K.B.; Rajnish, K.N. Bacterial exo-polysaccharides in biofilms: Role in antimicrobial resistance and treatments. J. Genet. Eng. Biotechnol. 2021, 19, 140. [Google Scholar] [CrossRef] [PubMed]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Dantas, L.R.; Suss, P.H.; Tasca Ribeiro, V.S. Pathogenesis of the Pseudomonas aeruginosa Biofilm: A Review. Pathogens 2022, 11, 300. [Google Scholar] [CrossRef]
- Rumbaugh, K.P.; Diggle, S.P.; Watters, C.M.; Ross-Gillespie, A.; Griffin, A.S.; West, S.A. Quorum Sensing and the Social Evolution of Bacterial Virulence. Curr. Biol. 2009, 19, 341. [Google Scholar] [CrossRef]
- Simanek, K.A.; Paczkowski, J.E. Resistance Is Not Futile: The Role of Quorum Sensing Plasticity in Pseudomonas aeruginosa Infections and Its Link to Intrinsic Mechanisms of Antibiotic Resistance. Microorganisms 2022, 10, 1247. [Google Scholar] [CrossRef]
- Daoud, A.; Ben Mefteh, F.; Mnafgui, K.; Turki, M.; Jmal, S.; Ben Amar, R.; Ayadi, F.; ElFeki, A.; Abid, L.; Rateb, M.E.; et al. Cardiopreventive effect of ethanolic extract of date palm pollen against isoproterenol induced myocardial infarction in rats through the inhibition of the angiotensin-converting enzyme. Exp. Toxicol. Pathol. 2017, 69, 656. [Google Scholar] [CrossRef]
- Ben Mefteh, F.; Daoud, A.; Chenari Bouket, A.; Thissera, B.; Kadri, Y.; Cherif-Silini, H.; Eshelli, M.; Alenezi, F.N.; Vallat, A.; Oszako, T.; et al. Date Palm Trees Root-Derived Endophytes as Fungal Cell Factories for Diverse Bioactive Metabolites. Int. J. Mol. Sci. 2018, 19, 1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alminderej, F.; Bakari, S.; Almundarij, T.I.; Snoussi, M.; Aouadi, K.; Kadri, A. Antioxidant Activities of a New Chemotype of Piper cubeba L. Fruit Essential Oil (Methyleugenol/Eugenol): In Silico Molecular Docking and ADMET Studies. Plants 2020, 9, 1534. [Google Scholar] [CrossRef] [PubMed]
- Bakari, S.; Ncir, M.; Felhi, S.; Hajlaoui, H.; Saoudi, M.; Gharsallah, N.; Kadri, A. Chemical composition and in vitro evaluation of total phenolic, flavonoid, and antioxidant properties of essential oil and solvent extract from the aerial parts of Teucrium polium grown in Tunisia. Food. Sci. Biotechnol. 2015, 24, 1943. [Google Scholar] [CrossRef]
- Kadri, A.; Zarai, Z.; Chobba, I.B.; Gharsallah, N.; Damak, M.; Békir, A. Chemical composition and in vitro antioxidant activities of Thymelaea hirsuta L: Essential oil from Tunisia. Afr. J. Biotechnol. 2011, 10, 2930. [Google Scholar]
- Aouadi, K.; Hajlaoui, H.; Arraouadi, S.; Ghannay, S.; Snoussi, M.; Kadri, A. HPLC/MS Phytochemical Profiling with Antioxidant Activities of Echium humile Desf. Extracts: ADMET Prediction and Computational Study Targeting Human Peroxiredoxin 5 Receptor. Agronomy 2021, 11, 2165. [Google Scholar] [CrossRef]
- Felhi, S.; Saoudi, M.; Daoud, A.; Hajlaoui, H.; Ncir, M.; Chaabane, R.; El Feki, A.; Gharsallah, N.; Kadri, A. Investigation of phytochemical contents, in vitro antioxidant and antibacterial behavior and in vivo anti-inflammatory potential of Ecballium elaterium methanol fruits extract. Food Sci. Technol. 2017, 37, 558. [Google Scholar] [CrossRef]
- Bakari, S.; Daoud, A.; Felhi, S.; Smaoui, S.; Gharsallah, N.; Kadri, A. Proximate analysis, mineral composition, phytochemical contents antioxidant and antimicrobial activities and GC-MS investigation of various solvent extracts of Cactus cladode. Food Sci. Technol. 2017, 27, 286. [Google Scholar] [CrossRef]
- Mseddi, K.; Alimi, F.; Noumi, E.; Veettil, V.N.; Deshpande, S.; Adnan, M.; Hamdi, A.; Elkahoui, S.; Alghamdi, A.; Kadri, A.; et al. Thymus musilii Velen. as a promising source of potent bioactive compounds with its pharmacological properties: In vitro and in silico analysis. Arab. J. Chem. 2020, 13, 6782. [Google Scholar] [CrossRef]
- Bakari, S.; Hajlaoui, H.; Daoud, A.; Mighri, H.; Ross-Garcia, J.M.; Gharsallah, N.; Kadri, A. Phytochemicals, antioxidant and antimicrobial potentials and LC-MS analysis of hydroalcoholic extracts of leaves and flowers of Erodium glaucophyllum collected from Tunisian Sahara. Food Sci. Technol. 2018, 38, 310. [Google Scholar] [CrossRef]
- Alminderej, F.; Bakari, S.; Almundarij, T.I.; Snoussi, M.; Aouadi, K.; Kadri, A. Antimicrobial and Wound Healing Potential of a New Chemotype from Piper cubeba L. Essential Oil and In Silico Study on S. aureus tyrosyl-tRNA Synthetase Protein. Plants 2021, 10, 205. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Mighri, H.; Aouni, M.; Gharsallah, N.; Kadri, A. Chemical composition and in vitro evaluation of antioxidant antimicrobial cytotoxicity and anti-acetylcholinesterase properties of Tunisian Origanum majorana L. essential oil. Microb. Pathog. 2016, 95, 86. [Google Scholar] [CrossRef] [PubMed]
- Merghni, A.; Noumi, E.; Hadded, O.; Dridi, N.; Panwar, H.; Ceylan, O.; Mastouri, M.; Snoussi, M. Assessment of the antibiofilm and antiquorum sensing activities of Eucalyptus globulus essential oil and its main component 1,8-cineole against methicillin-resistant Staphylococcus aureus strains. Microb. Pathog. 2018, 118, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Bieżanowska-Kopeć, R.; Piątkowska, E. Total Polyphenols and Antioxidant Properties of Selected Fresh and Dried Herbs and Spices. Appl. Sci. 2022, 12, 4876. [Google Scholar] [CrossRef]
- Felhi, S.; Hajlaoui, H.; Ncir, M.; Bakari, S.; Ktari, N.; Saoudi, M.; Gharsallah, N.; Kadri, A. Nutritional, phytochemical and antioxidant evaluation and FT-IR analysis of freeze-dried extracts of Ecballium elaterium fruit juice from three localities. Food Sci. Technol. 2016, 36, 646. [Google Scholar] [CrossRef]
- Marc, R.A.; Mureșan, V.; Mureșan, A.E.; Mureșan, C.C.; Tanislav, A.E.; Pușcaș, A.; Marţiș, G.S.; Ungur, R.A. Spicy and Aromatic Plants for Meat and Meat Analogues Applications. Plants 2022, 11, 960. [Google Scholar] [CrossRef]
- Pandey, S.; Patel, M.K.; Mishra, A.; Jha, B. Physio-Biochemical Composition and Untargeted Metabolomics of Cumin (Cuminum cyminum L.) Make It Promising Functional Food and Help in Mitigating Salinity Stress. PLoS ONE 2015, 10, e0144469. [Google Scholar] [CrossRef]
- Johri, R.K. Cuminum cyminum and Carumcarvi: An update. Pharmacogn Rev. 2011, 5, 63. [Google Scholar] [CrossRef]
- Singh, R.P.; Gangadharappa, H.V.; Mruthunjaya, K. Cuminum cyminum—A Popular Spice: An Updated Review. Pharmacogn. J. 2017, 9, 292. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Prasad, S.; Reuter, S.; Kannappan, R.; Yadev, V.R.; Park, B.; Kim, J.H.; Gupta, S.C.; Phromnoi, K.; Sundaram, C.; et al. Identification of novel anti-inflammatory agents from Ayurvedic medicine for prevention of chronic diseases: “reverse pharmacology” and “bedside to bench” approach. Curr. Drug Targets. 2011, 12, 1595. [Google Scholar] [CrossRef]
- Ramya, S.; Loganathan, T.; Chandran, M.; Priyanka, R.; Kavipriya, K.; Grace Lydial Pushpalatha, G.; Aruna, D.; Ramanathan, L.; Jayakumararaj, R.; Saluja, V. Phytochemical Screening, GCMS, FTIR profile of Bioactive Natural Products in the methanolic extracts of Cuminum cyminum seeds and oil. JDDT 2022, 12, 110. [Google Scholar] [CrossRef]
- Li, R.; Jiang, Z.-T. Chemical composition of the essential oil of Cuminum cyminum L. from China. Flavour Fragr. J. 2004, 19, 311. [Google Scholar] [CrossRef]
- Gachkar, L.; Yadegari, D.; Rezaei, M.B.; Taghizadeh, M.; Astaneh, S.A.; Rasooli, I. Chemical and biological characteristics of Cuminum cyminum and Rosmarinus officinalis essential oils. Food Chem. 2007, 102, 898. [Google Scholar] [CrossRef]
- Derakhshan, S.; Sattari, M.; Bigdeli, M. Effect of subinhibitory concentrations of cumin (Cuminum cyminum L.) seed essential oil and alcoholic extract on the morphology, capsule expression and urease activity of Klebsiella pneumoniae. Int. J. Antimicrob. Agents. 2008, 32, 432. [Google Scholar] [CrossRef] [PubMed]
- Mohammadpour, H.; Moghimipour, E.; Rasooli, I.; Fakoor, M.H.; Astaneh, S.A.; Moosaie, S.S.; Jalili, Z. Chemical Composition and Antifungal Activity of Cuminum cyminum L. Essential Oil From Alborz Mountain Against Aspergillus species. Jundishapur J. Nat. Pharm. Prod. 2012, 7, 50. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, F. Composition of Essential Oil of Cuminum cyminum. TEOP 2015, 18, 507. [Google Scholar]
- Ekhtelat, M.; Khalili Borujeni, F.; Siahpoosh, A.; Ameri, A. Chemical composition and antibacterial effects of some essential oils individually and in combination with sodium benzoate against methicillin-resistant Staphylococcus aureus and Yersinia enterocolitica. Vet. Res. Forum 2020, 11, 333. [Google Scholar] [PubMed]
- Ghasemi, G.; Fattahi, M.; Alirezalu, A. A new source of oxygenated monoterpenes with phytotoxic activity: Essential oil of Cuminum Cyminum L. from Iran. Nat. Prod. Res. 2020, 34, 843. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, A.; Mohammadzadeh, A.; Salehi, T.Z.; Mahmoodi, P.; Nourian, A. Cuminum cyminum L. Essential Oil: A Promising Antibacterial and Antivirulence Agent Against Multidrug-Resistant Staphylococcus aureus. Front. Microbiol. 2021, 12, 667833. [Google Scholar] [CrossRef]
- Pajohi Alamoti, M.; Bazargani-Gilani, B.; Mahmoudi, R.; Reale, A.; Pakbin, B.; Di Renzo, T.; Kaboudari, A. Essential Oils from Indigenous Iranian Plants: A Natural Weapon vs. Multidrug-Resistant Escherichia coli. Microorganisms 2022, 10, 109. [Google Scholar] [CrossRef]
- Hashemian, N.; Pirbalouti, A.G.; Hashemi, M.; Golparvar, A.; Hamedi, B. Diversity in chemical composition and antibacterial activity of essential oils of cumin (Cuminum cyminum L.) diverse from northeast of Iran. AJCS 2013, 7, 1752. [Google Scholar]
- Rana, V.S. Chemical composition of the essential oil of Cuminum cyminum L. seeds from Western India. J. Med. Plants. By-Prod. 2014, 3, 207. [Google Scholar]
- Patil, S.D.; Maknikar, P.P.; Wankhade, S.J.; Ukesh, C.S.; Rai, M.K. Chemical composition, antimicrobial and antioxidant activity of essential oils from cumin and ajowan. Nus. Biosci. 2016, 8, 60. [Google Scholar]
- Huo, Y.Y.; Li, T.T.; Yang, J.; Huang, H.Y.; Chen, C.J.; Xu, F.R.; Dong, X. Chemical Constituents of the Essential oil from Cuminum cyminum L. and Its Antifungal Activity against Panax notoginseng Pathogens. Chem. Biodivers. 2021, 18, e2100638. [Google Scholar] [CrossRef] [PubMed]
- Tanapichatsakul, C.; Khruengsai, S.; Pripdeevech, P. In vitro and in vivo antifungal activity of Cuminum cyminum essential oil against Aspergillus aculeatus causing bunch rot of postharvest grapes. PLoS ONE 2020, 15, e0242862. [Google Scholar] [CrossRef]
- Wongkattiya, N.; Sanguansermsri, P.; Fraser, I.H.; Sanguansermsri, D. Antibacterial activity of cuminaldehyde on food-borne pathogens, the bioactive component of essential oil from Cuminum cyminum L. collected in Thailand. J. Complement Integr. Med. 2019, 16, 20180195. [Google Scholar] [CrossRef] [PubMed]
- Bettaieb Rebey, R.; Jabri-Karoui, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Effect of drought on the biochemical composition and antioxidant activities of cumin (Cuminum cyminum L.) seeds. Ind. Crops Prod. 2012, 36, 238. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Mighri, H.; Noumi, E.; Snoussi, M.; Trabelsi, N.; Ksouri, R.; Bakhrouf, A. Chemical composition and biological activities of Tunisian Cuminum cyminum L. essential oil: A high effectiveness against Vibrio spp. strains. Food Chem. Toxicol. 2010, 48, 2186. [Google Scholar] [CrossRef]
- Jardak, M.; Mnif, S.; Ben Ayed, R.; Rezgui, F.; Aifa, S. Chemical composition, antibiofilm activities of Tunisian spices essential oils and combinatorial effect against Staphylococcus epidermidis biofilm. LWT 2021, 14, 110691. [Google Scholar] [CrossRef]
- Abushama, M.F.; Yasmin, H.; Abdalgadir, H.; Khalid, H. Chemical composition, antimicrobial and Brine Shrimp Lethality of the essential oil of Cuminum cyminum L. Int. J. Pharm. Chem. Sci. 2013, 2, 1666. [Google Scholar]
- Gómez-Mateos Pérez, M.; Navarro Moll, C.; Merino Espinosa, G.; Valero López, A. Evaluation of different Mediterranean essential oils as prophylactic agents in anisakidosis. Pharm. Biol. 2017, 55, 456. [Google Scholar] [CrossRef]
- Wanner, J.; Bail, S.; Jirovetz, L.; Buchbauer, G.; Schmidt, E.; Gochev, V.; Girova, T.; Atanasova, T.; Stoyanova, A. Chemical composition and antimicrobial activity of cumin oil (Cuminum cyminum, Apiaceae). Nat. Prod. Commun. 2010, 5, 1355. [Google Scholar] [CrossRef]
- Petretto, G.L.; Fancello, F.; Bakhy, K.; Faiz, C.A.L.; Sibawayh, Z.; Chessa, M.; Zara, S.; Sanna, M.L.; Maldini, M.; Rourke, J.P.; et al. Chemical composition and antimicrobial activity of essential oils from Cuminum cyminum L. collected in different areas of Morocco. Food Biosci. 2018, 22, 50. [Google Scholar] [CrossRef]
- Ghannay, S.; Aouadi, K.; Kadri, A.; Snoussi, M. GC-MS Profiling, Vibriocidal, Antioxidant, Antibiofilm, and Anti-Quorum Sensing Properties of Carum carvi L. Essential Oil: In Vitro and In Silico Approaches. Plants 2022, 11, 1072. [Google Scholar] [CrossRef] [PubMed]
- Pokhrel, N.R.; Adhikari, R.; Baral, M. In-vitro evaluation of the antimicrobial activity of Bauhinia variegata, locally known as koiralo. World J. Microbiol. Biotechnol. 2002, 18, 69. [Google Scholar] [CrossRef]
- Mahfuzul Hoque, M.D.; Bari, M.; Inatsu, Y.; Juneja, V.K.; Kawamoto, S. Antibacterial activity of guava (Psidium guajava L.) and neem (Azadirachta indica A. Juss.) extracts against foodborne pathogens and spoilage bacteria. Foodborne Pathog. Dis. 2007, 4, 481. [Google Scholar] [CrossRef]
- Mahboubi, M.; Haghi, G. Antimicrobial activity and chemical composition of Mentha pulegium L. essential oil. J. Ethnopharmacol. 2008, 119, 325. [Google Scholar] [CrossRef]
- Yano, Y.; Satomi, M.; Oikawa, H. Antimicrobial effect of spices and herbs on Vibrio parahaemolyticus. Int. J. Food Microbiol. 2006, 11, 6. [Google Scholar] [CrossRef]
- Snoussi, M.; Hajlaoui, H.; Noumi, E.; Usai, D.; Sechi, L.A.; Zanetti, S.; Bakhrouf, A. In-vitro anti-Vibrio spp. activity and chemical composition of some Tunisian aromatic plants. World J. Microbiol. Biotechnol. 2008, 24, 3071. [Google Scholar] [CrossRef]
- Hossain, M.A.; Kabir, M.; Salehuddin, S.; Rahman, S.M.; Das, A.; Singha, S.K.; Alam, M.K.; Rahman, A. Antibacterial properties of essential oils and methanol extracts of sweet basil Ocimum basilicum occurring in Bangladesh. Pharm. Biol. 2010, 48, 504. [Google Scholar] [CrossRef]
- Pirbalouti, A.G.; Hamedi, B.; Poor, F.M.; Rahimi, E.; Nejhad, R.N. Inhibitory activity of Iranian endemic medicinal plants against Vibrio parahaemolyticus and Vibrio harveyi. J. Med. Plants Res. 2011, 5, 7049. [Google Scholar]
- Wei, L.S.; Wee, W. Chemical composition and antimicrobial activity of Cymbopogon nardus citronella essential oil against systemic bacteria of aquatic animals. Iran J. Microbiol. 2013, 5, 147. [Google Scholar] [PubMed]
- Gracia-Valenzuela, M.H.; Vergara-Jiménez, M.J.; Baez-Flores, M.E.; Cabrera-Chavez, F. Antimicrobial effect of dietary oregano essential oil against Vibrio bacteria in shrimps. Arch. Biol. Sci. 2014, 66, 1367. [Google Scholar] [CrossRef]
- Melissa, M.; Ana, G.; Samuel, M.; Elizabeth, P.; Marisol, Ã.; Rocà o, S.; Julieta, O.; Manuel, J.; Carlos, C.J.; Ignacio, P.; et al. Antimicrobial activity of essential oil of Cordia globosa. Afr. J. Pharm. Pharmacol. 2016, 10, 179. [Google Scholar]
- Mahbobi, M.; Akbari, M.; Haghi, G.; Kazempuor, N. Comparison of Antimicrobial Activity of Respitol-B With Mentofin Containing Menthol, Eucalyptus Oil. Iran J. Med. Microbiol. 2007, 1, 39. [Google Scholar]
- Park, J.-W.; Wendt, M.; Heo, G.-J. Antimicrobial activity of essential oil of Eucalyptus globulus against fish pathogenic bacteria. Lab. Anim. Res. 2016, 32, 87. [Google Scholar] [CrossRef]
- Al-Sahlany, S.T.G. Effect of Mentha piperita essential oil against Vibrio spp. isolated from local cheeses. Pak. J. Food Sci. 2016, 26, 65. [Google Scholar]
- Snoussi, M.; Noumi, E.; Dehmani, A.; Flamini, G.; Aouni, M.; Alsieni, M.; Al-sieni, A. Chemical composition and antimicrobial activities of Elettaria cardamomum L. (Manton) essential oil: A high activity against a wide range of food borne and medically important bacteria and fungi. J. Chem. Biol. Phys. Sci. 2015, 6, 248. [Google Scholar]
- Snoussi, M.; Noumi, E.; Trabelsi, N.; Flamini, G.; Papetti, A.; De Feo, V. Mentha spicata essential oil: Chemical composition, antioxidant and antibacterial activities against planktonic and biofilm cultures of Vibrio spp. strains. Molecules 2015, 20, 14402. [Google Scholar] [CrossRef]
- Snoussi, M.; Dehmani, A.; Noumi, E.; Flamini, G.; Papetti, A. Chemical composition and antibiofilm activity of Petroselinum crispum and Ocimum basilicum essential oils against Vibrio spp. strains. Microb. Pathog. 2016, 90, 13. [Google Scholar] [CrossRef]
- Manju, S.; Malaikozhundan, B.; Withyachumnarnkul, B.; Vaseeharan, B. Essential oils of Nigella sativa protects Artemia from the pathogenic effect of Vibrio parahaemolyticus Dahv2. J. Invertebr. Pathol. 2016, 136, 43. [Google Scholar] [CrossRef]
- Partovi, R.; Khanjari, A.; Abbaszadeh, S.; Sharifzadeh, A. Chemical composition and antimicrobial effect of five essential oils on pathogenic and non-pathogenic Vibrio parahaemolyticus. Nut. Food Sci. Res. 2017, 4, 43. [Google Scholar] [CrossRef]
- Hamad, A.; Alifah, A.; Permadi, A.; Hartanti, D. Chemical constituents and antibacterial activities of crude extract and essential oils of Alpinia galanga and Zingiber officinale. Int. Food Res. J. 2016, 23, 837. [Google Scholar]
- See, C.; Jenwitheesuk, E. Antimicrobial Activity of a Blend of Essential Oils Extracted from Oregano and Cinnamon. J. Anim. Sci. Res. 2018, 2, 1–3. [Google Scholar]
- Mendes, J.L.; de Araújo, T.F.; Geraldo de Carvalho, M.; Catunda, A.; Júnior, F.E. Chemical composition and mechanism of vibriocidal action of essential oil from resin of Protium heptaphyllum. Sci. J. 2019, 2019, 9563213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Feyaerts, A.F.; Van Dijck, P.; Bossier, P. Inhibitory Activity of Essential Oils against Vibrio campbellii and Vibrio parahaemolyticus. Microorganisms 2020, 8, 1946. [Google Scholar] [CrossRef] [PubMed]
- Mizan, M.F.R.; Ashrafudoulla, M.; Hossain, M.I.; Cho, H.R.; Ha, S.D. Effect of essential oils on pathogenic and biofilm-forming Vibrio parahaemolyticus strains. Biofouling 2020, 36, 467. [Google Scholar] [CrossRef]
- Hajlaoui, H.; Arraouadi, S.; Noumi, E.; Aouadi, K.; Adnan, M.; Khan, M.A.; Kadri, A.; Snoussi, M. Antimicrobial, Antioxidant, Anti-Acetylcholinesterase, Antidiabetic, and Pharmacokinetic Properties of Carum carvi L. and Coriandrum sativum L. Essential Oils Alone and in Combination. Molecules 2021, 26, 3625. [Google Scholar] [CrossRef]
- Chen, Q.; Gan, Z.; Zhao, J.; Wang, Y.; Zhang, S.; Li, J.; Ni, Y. In vitro comparison of antioxidant capacity of cumin (Cuminum cyminum L.) oils and their main components. LWT-Food Sci. Technol. 2014, 55, 632. [Google Scholar] [CrossRef]
- Moghaddam, M.; Miranb, S.N.K.; Pirbalouti, A.G.; Mehdizadehd, l.; Ghaderi, Y. Variation in essential oil composition and antioxidant activity of cumin (Cuminum cyminum L.) fruits during stages of maturity. Ind. Crops Prod. 2015, 70, 163. [Google Scholar] [CrossRef]
- Akrami, F.; Rodríguez-Lafuente, A.; Bentayeb, K.; Pezo, D.; Ghalebi, S.R.; Nerín, C. Antioxidant and antimicrobial active paper based on Zataria (Zataria multiflora) and two cumin cultivars (Cuminum cyminum). LWT-Food Sci. Technol. 2015, 60, 929. [Google Scholar] [CrossRef]
- Derakhshan, S.; Sattari, M.; Bigdeli, M. Effect of cumin (Cuminum cyminum) seed essential oil on biofilm formation and plasmid Integrity of Klebsiella pneumoniae. Pharmacogn. Mag. 2010, 6, 57. [Google Scholar] [PubMed]
- Walmiki, M.R.; Rai, V.R. Cell Attachment Inhibition and Anti-biofilm Activity of Syzygium aromaticum, Cuminum cyminum and Piper nigrum Essential Oils Against Pathogenic Bacteria. J. Essent. Oil Bear. Plants 2017, 20, 59–68. [Google Scholar] [CrossRef]
- Packiavathy, I.A.S.V.; Agilandeswari, P.; Musthafa, K.S.; Pandian, S.K.; Ravi, A.V. Antibiofilm and quorum sensing inhibitory potential of Cuminum cyminum and its secondary metabolite methyl eugenol against Gram negative bacterial pathogens. Food Res. Int. 2012, 45, 85. [Google Scholar] [CrossRef]
- Jayalekshmi, S.K.; Ramasamy, S. Antibiofilm and antiquorum sensing potential of Cuminum cyminum against Aaeromonas veronii. J. Adv. Sci. Res. 2020, 11, 105. [Google Scholar]
- Chatterjee, S.; Paul, P.; Chakraborty, P.; Das, S.; Sarker, R.K.; Sarkar, S.; Das, A.; Tribedi, P. Cuminaldehyde exhibits potential antibiofilm activity against Pseudomonas aeruginosa involving reactive oxygen species (ROS) accumulation: A way forward towards sustainable biofilm management. 3 Biotech 2021, 11, 485. [Google Scholar] [CrossRef]
- Naveed, R.; Hussain, I.; Tawab, T.; Tariq, M.; Rahman, M.; Hameed, S.; Mahmood, M.S.; Siddique, A.B.; Iqbal, M. Antimicrobial activity of the bioactive components of essential oils from Pakistani spices against Salmonella and other multi-drug resistant bacteria. BMC Complement Altern. Med. 2013, 13, 265. [Google Scholar] [CrossRef]
- Monteiro-Neto, V.; de Souza, C.D.; Gonzaga, L.F.; da Silveira, B.C.; Sousa, N.C.F.; Pontes, J.P.; Santos, D.M.; Martins, W.C.; Pessoa, J.F.V.; Carvalho Júnior, A.R.; et al. Cuminaldehyde potentiates the antimicrobial actions of ciprofloxacin against Staphylococcus aureus and Escherichia coli. PLoS ONE 2020, 15, e0232987. [Google Scholar] [CrossRef]
- Snoussi, M.; Noumi, E.; Punchappady-Devasya, R.; Trabelsi, N.; Kanekar, S.; Nazzaro, F.; Fratianni, F.; Flamini, G.; De Feo, V.; Al-Sieni, A. Antioxidant properties and anti-quorum sensing potential of Carum copticum essential oil and phenolics against Chromobacterium violaceum. J. Food Sci. Technol. 2018, 55, 2824. [Google Scholar] [CrossRef]
- Othman, I.M.M.; Gad-Elkareem, M.A.M.; Anouar, E.H.; Snoussi, M.; Aouadi, K.; Kadri, A. Novel fused pyridine derivatives containing pyrimidine moiety as prospective tyrosyl-tRNA synthetase inhibitors: Design, synthesis, pharmacokinetics and molecular docking studies. J. Mol. Struct. 2020, 1219, 128651. [Google Scholar] [CrossRef]
- Othman, I.M.; Gad-Elkareem, M.A.; Aouadi, K.; Kadri, A.; Snoussi, M. Design, synthesis ADMET and molecular docking of new imidazo [4,5-b] pyridine-5-thione derivatives as potential tyrosyl-tRNA synthetase inhibitors. Bioorganic Chem. 2020, 102, 104105. [Google Scholar] [CrossRef]
- Ghannay, S.; Kadri, A.; Aouadi, K. Synthesis, in vitro antimicrobial assessment, and computational investigation of pharmacokinetic and bioactivity properties of novel trifluoromethylated compounds using in silico ADME and toxicity prediction tools. Monatsh. Chem. 2020, 151, 267–280. [Google Scholar] [CrossRef]
- Kadri, A.; Aouadi, K. In vitro antimicrobial and α-glucosidase inhibitory potential of enantiopure cycloalkylglycine derivatives: Insights into their in silico pharmacokinetic, druglikeness, and medicinal chemistry properties. J. App. Pharm. Sci. 2020, 10, 107–115. [Google Scholar]
- Ghannay, S.; Bakari, S.; Msaddek, M.; Vidal, S.; Kadri, A.; Aouadi, K. Design, synthesis, molecular properties and in vitro antioxidant and antibacterial potential of novel enantiopure isoxazolidine derivatives. Arab. J. Chem. 2020, 13, 2121–2131. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Components | l.r.i. a | Percentage | Molecular Weight | Chemical Formula |
|---|---|---|---|---|---|
| 1 | α-thujene | 933 | 0.4 | 136.23 | C10H16 |
| 2 | α-pinene | 941 | 0.9 | 136.23 | C10H16 |
| 3 | Sabinene | 978 | 0.3 | 136.23 | C10H16 |
| 4 | β-pinene | 982 | 15.1 | 136.238 | C10H16 |
| 5 | Myrcene | 993 | 0.6 | 136.238 | C10H16 |
| 6 | α-phellandrene | 1006 | 0.3 | 136.23 | C10H16 |
| 7 | p-cymene | 1028 | 14.2 | 134.22 | C10H14 |
| 8 | Limonene | 1032 | 0.5 | 136.24 | C10H16 |
| 9 | γ-terpinene | 1064 | 14.4 | 136.234 | C10H16 |
| 10 | Linalool | 1101 | 0.1 | 154.253 | C10H18O |
| 11 | 4-terpineol | 1179 | 0.4 | 154.25 | C10H18O |
| 12 | α-terpineol | 1191 | 0.2 | 154.25 | C10H18O |
| 13 | Cuminaldehyde | 1240 | 42.4 | 148.205 | C10H12O |
| 14 | Carvone | 1242 | 0.1 | 150.22 | C10H14O |
| 15 | Phellandral | 1274 | 0.2 | 152.23 | C10H16O |
| 16 | α-terpin-7-al | 1283 | 5.2 | 150.22 | C10 H14O |
| 17 | γ-terpin-7-al | 1288 | 2.7 | 150.22 | C10H14O |
| 18 | β-caryophyllene | 1419 | 0.3 | 204.36 | C15H24 |
| 19 | γ-muurolene | 1478 | 0.4 | 204.35 | C15H24 |
| 20 | Carotol | 1595 | 0.4 | 222.37 | C15H26O |
| Chemical classes | |||||
| Monoterpene hydrocarbons | 46.7% | ||||
| Oxygenated monoterpenes | 51.3% | ||||
| Sesquiterpene hydrocarbons | 0.7% | ||||
| Oxygenated sesquiterpenes | 0.4% | ||||
| Total identified | 99.1% | ||||
| Antioxidant Tests | Cumin EO IC50 (mg/mL) | AA EC50 (mg/mL) | BHT IC50 (mg/mL) | BHA IC50 (mg/mL) |
|---|---|---|---|---|
| DPPH (IC50 mg/mL) | 8 ± 0.54 b | 12 ± 0.01 a | 11.50 ± 0.62 a | - |
| Reducing power (EC50 mg/mL) | 3.50 ± 0.03 c | 25 ± 0.01 a | 23.00 ± 1.0 b | - |
| β-carotene (IC50 mg/mL) | 3.80 ± 0.34 b | - | 4.60 ± 1.60 a | - |
| Chelating Power (IC50 mg/mL) | 8.40 ± 0.14 b | - | - | 32.50 ± 1.32 a |
| Vibrio spp. Tested | Cumin EO | |||
|---|---|---|---|---|
| mZI ± SD (mm) | MIC ± SD (mg/mL) | MBC ± SD (mg/mL) | MBC/MIC Ratio | |
| V. cholerae ATCC 9459 | 34.33 ± 0.58 ef | 0.023 | 6 | >4; Bacteriostatic |
| V. vulnificus ATCC 27562 | 30.33 ± 0.58 g | 0.023 | 1.5 | >4; Bacteriostatic |
| V. parahaemolyticus ATCC 17802 | 39.67 ± 0.58 b | 0.046 | 12 | >4; Bacteriostatic |
| V. parahaemolyticus ATCC 43996 | 28.67 ± 1.15 h | 0.023 | 1.5 | >4; Bacteriostatic |
| V. alginolyticus ATCC 33787 | 34.67 ± 0.58 de | 0.023 | 3 | >4; Bacteriostatic |
| V. alginolyticus ATCC 17749 | 33.33 ± 0.58 f | 0.023 | 6 | >4; Bacteriostatic |
| V. furnisii ATCC 35016 | 11.33 ± 0.58 k | 0.023 | 3 | >4; Bacteriostatic |
| V. cincinnatiensis ATCC 35912 | 14.67 ± 0.28 j | 0.046 | 12 | >4; Bacteriostatic |
| V. proteolyticus ATCC 15338 | 30.33 ± 0.58 g | 0.023 | 6 | >4; Bacteriostatic |
| V. natrigens ATCC 14048 | 36.67 ± 0.58 c | 0.023 | 3 | >4; Bacteriostatic |
| V. mimicus ATCC 33653 | 28.67 ± 0.58 h | 0.046 | 12 | >4; Bacteriostatic |
| V. fluvialis ATCC 33809 | 41.33 ± 1.15 a | 0.046 | 3 | >4; Bacteriostatic |
| V. carhiaccae ATCC 35084 | 35.33 ± 0.58 d | 0.046 | 6 | >4; Bacteriostatic |
| V. harveyi ATCC 18293 | 35.67 ± 0.58 cd | 0.023 | 3 | >4; Bacteriostatic |
| V. diazotrophicus ATCC 33466 | 11.00 ± 0.00 k | 0.023 | 3 | >4; Bacteriostatic |
| V. tapetis CECT 4600T | 30.67 ± 0.58 g | 0.046 | 6 | >4; Bacteriostatic |
| V. splendidus ATCC 33125 | 26.33 ± 0.58 i | 0.046 | 6 | >4; Bacteriostatic |
| Control | Concentrations Tested (mg/mL) | |||||
|---|---|---|---|---|---|---|
| 10 | 50 | 125 | 250 | 500 | ||
| Diameter of the colony (mm± SD) | ||||||
| Cumin EO | 54.00 ± 0.00 a | 19.33 ± 0.57 b | 14.67 ± 0.57 c | 12.00 ± 0.00 d | 10.33 ± 0.57 e | 8.67 ± 0.57 f |
| Cuminaldehyde | 54.00 ± 0.00 a | 15.67 ± 0.57 b | 13.67 ± 0.57 c | 12.00 ± 0.00 d | 10.33 ± 0.57 e | 9.00 ± 0.00 f |
| Percentage of motility inhibition (%) | ||||||
| Cumin EO | 100 ± 0.00 | 64.20 ± 0.57 | 77.15 ± 0.57 | 84.45 ± 0.00 | 87.76 ± 0.57 | 90.12 ± 0.57 |
| Cuminaldehyde | 100 ± 0.00 | 70.99 ± 0.57 | 80.75 ± 0.57 | 85.96 ± 0.57 | 87.98 ± 0.57 | 89.77 ± 0.00 |
| Entry | 4 | 7 | 9 | 13 | 16 | 17 | Reference |
|---|---|---|---|---|---|---|---|
| Absorption | |||||||
| Water solubility | −4.221 | −5.163 | −3.941 | −3.923 | −2.79 | −2.79 | - |
| Caco2 permeability | 1.373 | 1.399 | 1.414 | 1.503 | 1.517 | 1.517 | >0.9 |
| Intestinal absorption (human) | 94.607 | 94.256 | 96.219 | 95.543 | 97.506 | 97.506 | <30% is poorly |
| Skin Permeability (log Kp) | −1.646 | −1.2 | −1.489 | −1.425 | −2.624 | −2.624 | >−2.5 is low |
| Distribution | |||||||
| P-glycoprotein substrate | No | No | No | No | No | No | No |
| P-glycoprotein I inhibitor | No | No | No | No | No | No | No |
| P-glycoprotein II inhibitor | No | No | No | No | No | No | No |
| VDss (human) | 0.68 | 0.455 | 0.412 | 0.274 | 0.233 | 0.233 | Low is <−0.15, High is >0.45 |
| Fraction unbound (human) | 0.353 | 0.262 | 0.42 | 0.305 | 0.465 | 0.465 | - |
| BBB permeability | 0.812 | 0.785 | 0.754 | 0.664 | 0.633 | 0.633 | Poorly is <−1, High is >0.3 |
| CNS permeability | −1.837 | −1.359 | −2.049 | −1.506 | −2.197 | −2.197 | Penetrate is >−2, Unable is <−3 |
| Metabolism | |||||||
| CYP2D6 substrate | No | No | No | No | No | No | No |
| CYP3A4 substrate | No | No | No | No | No | No | - |
| CYP1A2 inhibitior | No | No | No | No | No | No | No |
| CYP2C19 inhibitior | No | No | No | No | No | No | No |
| CYP2C9 inhibitior | No | No | No | No | No | No | No |
| CYP2D6 inhibitior | No | No | No | No | No | No | No |
| CYP3A4 inhibitior | No | No | No | No | No | No | No |
| Excretion | |||||||
| Total Clearance | 0.03 | 1.163 | 0.217 | 0.212 | 0.182 | 0.182 | - |
| Renal OCT2 substrate | No | No | No | No | No | No | - |
| Toxicity | |||||||
| AMES toxicity | No | No | No | No | No | No | No |
| Max. tolerated dose (human) | 0.24 | 0.193 | 0.756 | 0.128 | 0.723 | 0.723 | Low is ≤0.477, High is >0.477 |
| hERG I inhibitor | No | No | No | No | No | No | No |
| hERG II inhibitor | No | No | No | No | No | No | No |
| Oral Rat Acute Toxicity (LD50) | 1.617 | 1.533 | 1.766 | 1.499 | 1.971 | 1.971 | - |
| Oral Rat Chronic Toxicity (LOAEL) | 2.247 | 2.411 | 2.394 | 2.052 | 2.034 | 2.034 | - |
| Hepatotoxicity | No | No | No | No | No | No | No |
| Skin Sensitisation | No | No | No | Yes | Yes | Yes | No |
| T.Pyriformis toxicity | 0.633 | 0.767 | 0.627 | 0.765 | 0.732 | 0.732 | >−0.5 is toxic |
| Minnow toxicity | 1.131 | 0.65 | 0.906 | 0.862 | 1.118 | 1.118 | <−0.3 is toxic |
| Compounds | Interacting Residues Receptor vs. Targets | Binding Energy (kcal/mol) |
|---|---|---|
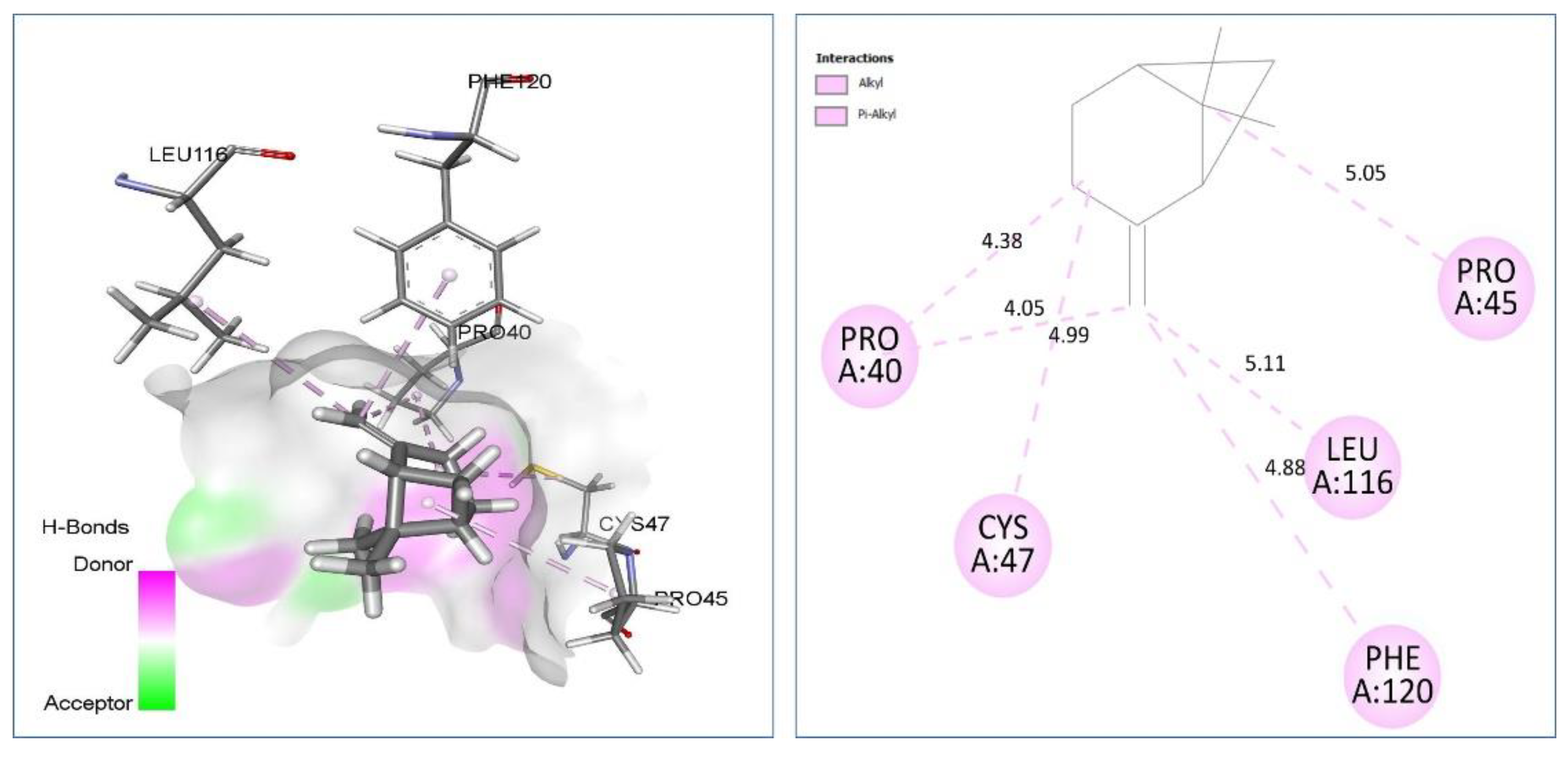
| β-pinene vs. 1HD2 | Alkyl/Pi-Alkyl: Pro40 (4.05) (4.38), Pro45 (5.05), Cys47 (4.99), Leu116 (5.11), Phe120 (4.88). | −4.6 |
| Cuminaldehyde vs. 1HD2 | H bond: Thr147 (2.10). Alkyl: Pro45 (5.14), Cys47 (5.00). | −5.4 |
| Cuminaldehyde vs. 1JIJ | C-H bond: Gly192 (2.81). Alkyl/Pi-Alkyl: Cys37 (5.21), Leu70 (4.89) (5.39). | −7.4 |
| β-Caryophyllene vs. IJIJ | Alkyl/Pi-Alkyl: Cys37(4.64), Ala39 (4.28) (4.53) (4.75), Pro53 (5.37) (4.50), His50 (4.00) (5.06). | −6.4 |
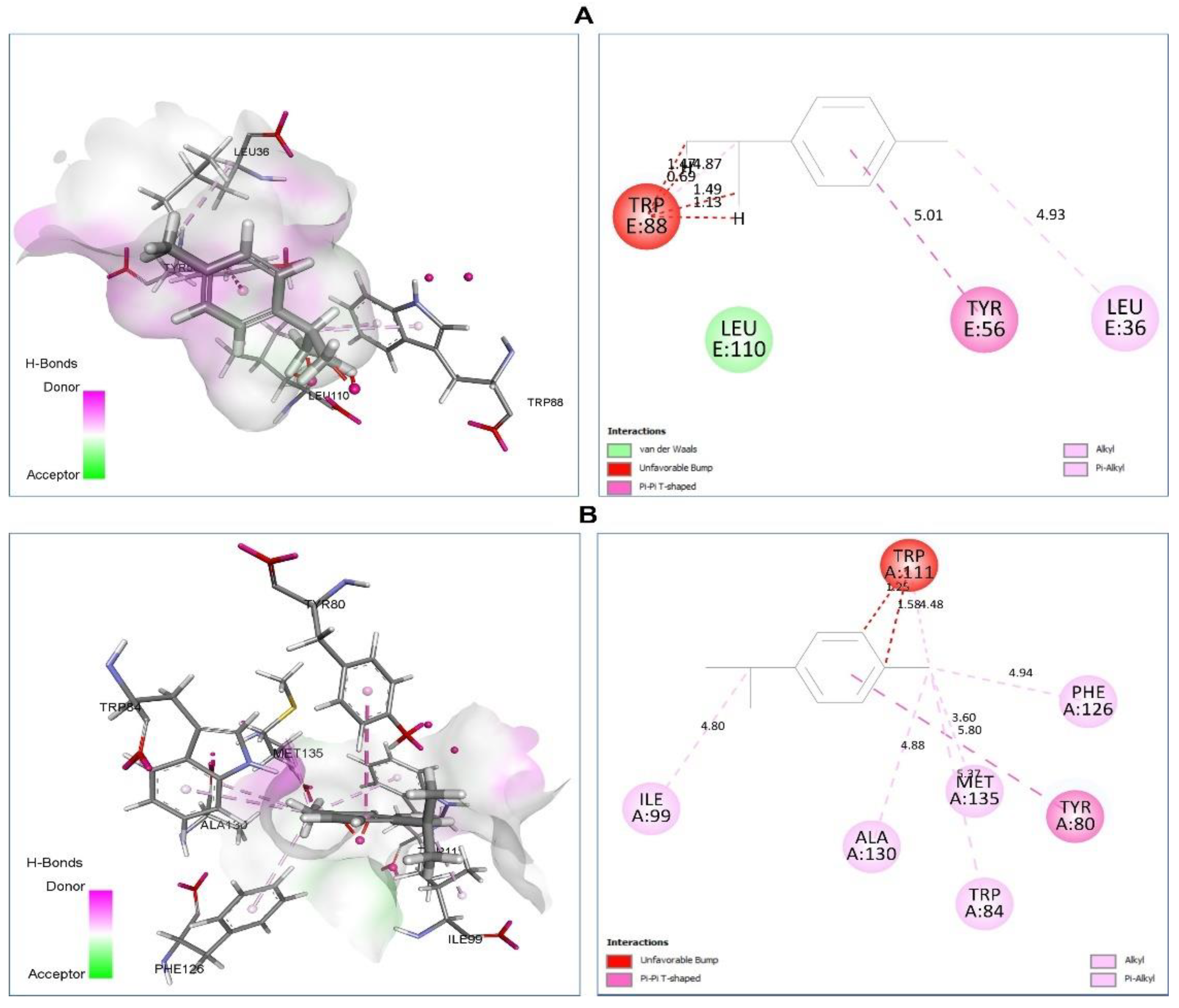
| p-Cymene vs. 2UV0 | van der Waals: Leu110. Unfavorable Bump: Trp88 (0.69) (1.13) (1.47) (1.49). Pi-Pi T-Shaped: Tyr56 (5.01). Alkyl/Pi-Alkyl: Leu36 (4.93), Trp88 (4.87). | −7.4 |
| γ-Terpinene vs. 2UV0 | Unfavorable Bump: Trp88 (0.61) (1.34) (1.54). Alkyl/Pi-Alkyl: Leu36 (4.93), Tyr56 (5.20), Tyr64 (3.76) (4.88), Trp88 (4.67). | −7.4 |
| Cuminaldehyde vs. 2UV0 | H bond: Arg61 (4.31). Pi-Lone Pair: Tyr64 (2.79). Unfavorable Bump: Trp88 (0.71) (0.75) (0.22) (1.39). Pi-Pi T-Shaped: Tyr56 (4.90). Alkyl/Pi-Alkyl: Trp88 (4.75). | −7.4 |
| p-Cymene vs. 3QP1 | Unfavorable Bump: Trp111 (1.25) (1.58). Pi-Pi T-Shaped: Tyr80 (5.80). Alkyl/Pi-Alkyl: Trp84 (3.37) Ile99 (4.80), Phe126m(4.94), Ala130 (4.88). Met135 (5.80). | −7.5 |
| γ-Terpinene vs. 3QP1 | van der Waals: Leu57, trp84, Tyr88, Ile99, Leu100. Unfavorable Bump: Trp111 (0.90) (1.41). Alkyl/Pi-Alkyl: Tyr80 (5.36), Phe115 (4.99), Phe126 (4.99), Ala130 (5.01), Met135 (3.80) (5.35), Trp111 (4.20) (4.45). | −7.5 |
| Cuminaldehyde vs. 3QP1 | Pi-Pi T-Shaped: Tyr80 (5.82). Pi-Alkyl: Ile99 (4.76). | −7.2 |
| Origin | Chemical Composition (Main Constituents) | References |
|---|---|---|
| China | Cuminaldehyde (36.31%), cuminic alcohol (16.92%), γ-terpinene (11.14%), safranal (10.87%), p-cymene (9.85%) and β-pinene (7.75%) | [31] |
| Iran | α-pinene (29.1%), limonene (21.5%), 1,8-cineole (17.9%), and linalool (10.4%) | [32] |
| Cuminaldehyde (25.2%), p-mentha-1,3-dien-7-al (13%), p-mentha-1,4-dien-7-al (16.6%), γ-terpinene (19%), p-cymene (7.2%), and β-pinene (10.4%). | [33] | |
| α-Pinene (29.2%), limonene (21.7%), 1,8-cineole (18.1%), linalool (10.5%), linalyl acetate (4.8%), and α-terpineole (3.17%). | [34] | |
| α-pinene (30.12%), limonene (10.11%), 1,8-cineole (11.54%), γ-terpinene (3.56%), linalool (10.3%), sabinene (1.11%), p-cymene (0.6%), α-campholenal (1.76%), linalyl acetate (4.76%), α-terpinyl acetate (1.8%), neryl acetate (1%). | [35] | |
| Cuminaldehyde (28.24%), γ-terpinene (21.39%), o-Cymene (13.78%), β-pinene (3.14%), and β-Acoradiene (1.68%). | [36] | |
| 3-caren-10-al (47.27%), cuminal (25.92%), 2-caren-10-al (8.05%), γ-terpinene (7.66%), (-)-β-pinene (5.11%), and p-cymene (2.71%). | [37] | |
| Cuminaldehyde (38.26%), α,β-dihydroxy ethylbenzene (29.16%), 2-caren-10-al (11.20%), γ-terpinene (6.49%), and β-pinene (5.25%). | [38] | |
| Cuminaldehyde (29.0%), α-terpinen-7-al (20.7%), γ-terpinene (12.94%), γ-terpinen-7-al (8.91%), p-cymene (8.55%), and β-pinene (7.72%). | [39] | |
| India | Safranal (16.8–29.0%), γ-terpinene (14.1–19.6%), γ-terpinene-7-al (13.5–25.5%), cuminaldehyde (17.5–22.3%), β-pinene (6.8–10.4%), and p-cymene (4.1–8.8%). | [40] |
| Cuminaldehyde (49.4%), p-cymene (17.4%), β-pinene (6.3%), α-terpinen-7-al (6.8%), γ-terpinene (6.1%), p-cymen-7-ol (4.6%), and thymol (2.8%). | [41] | |
| Cuminaldehyde (36.67%), caren-10-al (21.34%), β-pinene (18.76%), γ-terpinene (16.86%), terpinen-4-ol (2.44%), α-thujene (1.88%), α-pinene (1.41%), p-cymene (0.30%), carbicol (0.19%) and α-terpineol (0.09%). | [42] | |
| China | Cuminaldehyde (44.53%), p-cymene (12.14%), β-pinene (10.47%) and γ-terpinene (8.40%) | [43] |
| Thailand | Cumin aldehyde (33.94%), α-terpinen-7-al (32.20%), γ-terpinen-7-al (13.74%), γ-terpinene (6.67%), β-pinene (5.34%) and p-cymene (3.58%). | [44] |
| Cuminaldehyde (27.10%), β-pinene (25.04%) and γ-terpinene (15.68%). | [45] | |
| Tunisia | γ-terpinen (25.58%), 1-phenyl-1,2 ethanediol (23.16%), cuminaldehyde (15.31%), β-pinene (15.16%), and ρ-cymene (9.05%) | [46] |
| Cuminaldehyde (39.48%), γ-terpinene (15.21%), O-cymene (11.82%), β-pinene (11.13%), 2-caren-10-al 7.93%), trans-carveol (4.49%) and myrtenal (3.5%). | [47] | |
| Cuminaldehyde (28.22%), 1-phenyl-1-butanol (23.33%), β-pinene (12.61%) and p-cymene (11.72%). | [48] | |
| Sudan | 2-Caren-10-al (29.64%), benzaldehyde, 4-1-methyethyl (16.58%), and 2-J-pinene (12.06%) | [49] |
| Spain | Cuminaldehyde (34.11%), Δ2-Caren-10-al (20.78%), p-cymene (12.25%), Δ3-C10-al (11.80%), Δ4-Carene (10.47%), β-pinene (7.3%). | [50] |
| Iran | Cuminaldehyde (41.5%), p-cymene (17.4%), β-pinene (10.7%), γ-Terpinene (6.5%), p-mentha-1,3-dien-7-al (5.5%), p-mentha-1,4-dien-7-al (1.5%), β-acoradiene (3.5%). | [51] |
| Egypt | Cuminaldehyde (29.3%), γ-Terpinene (18.5%), β-pinene (15.7%), p-mentha-1,3-dien-7-al (10.6%), p-cymene (10.1%), p-mentha-1,4-dien-7-al (7.6%), β-acoradiene (0.2%). | |
| India | γ-Terpinene (31.1%), cuminaldehyde (23.2%), p-cymene (18.4%), β-pinene (12.6%), p-mentha-1,3-dien-7-al (7.2%), p-mentha-1,4-dien-7-al (0.4%), β-acoradiene (0.1%). | |
| Europe | γ-Terpinene (26.5%), cuminaldehyde (22.4%), p-cymene (20.2%), β-pinene (14.1%), p-mentha-1,3-dien-7-al (6.6%), p-mentha-1,4-dien-7-al (1.4%), β-acoradiene (0.3%). | |
| Morocco | β-pinene (20.8–86.4%), p-cymene (6.2–24.7%), γ-terpinene (18.1–90.7%), cuminaldehyde (51.5–91.5%), α-terpinen-7-al (21.2–95.3%) and α-terpinen-7-al (22.6–55.06%) | [52] |
| Plant Species Tested | Vibrio Species Tested | References |
|---|---|---|
| Bauhinia variegata | V. cholerae | [54] |
| Psidium guajava, Azadirachta indica | V. cholerae | [55] |
| Mentha pulegium | V. cholerae | [56] |
| Syzygium aromaticum | V. parahaemolyticus | [57] |
| Mentha longifolia; M. pulegium; Eugenia caryophyllata; Rosmarinus officinalis and Thymus vulgaris | V. alginolyticus, V. parahaemolyticus, V. fluvialis, V. vulnificus | [58] |
| C. cyminum | V. cholerae, V. parahaemolyticus, V. alginolyticus, V. vulnificus, V. harveyi, V. proteolyticus, V. furnisii, V. mimicus, V. furnisii, V. natrigens, V. carhiaccae, V. fluvialis | [47] |
| Ocimum basilicum | V. parahaemolyticus, V. mimicus | [59] |
| Satureja bachtiarica Bunge, Zataria multiflora | V. parahaemolyticus, V. harveyi. | [60] |
| Cymbopogon nardus | V. damsela, Vibrio spp. | [61] |
| Lippia berlandieri | V. cholerae, V. parahaemolyticus, V. vulnificus | [62] |
| Cordia globosa | V. cholerae | [63] |
| Eucalyptus globulus | V. cholerae | [64] |
| V. harveyi, V. ichthyoenteri | [65] | |
| Mentha piperita | V. parahaemolyticus, V. cholerae, V. vulnificus, V. alginolyticus, V. mimicus, V. damsela, V. campbellii, V. harveyi, V. logei | [66] |
| Elettaria cardamomum, Mentha spicata, Petroselinum crispum, Ocimum basilicum | V. cholerae, V. vulnificus, V. parahaemolyticus, V. alginolyticus, V. furnisii, V. cincinnatiensis, V. proteolyticus, V. natrigens, V. mimicus, V. fluvialis, V. anguillarum, V. carrichariae, V. harveyii, V. diazotrophicus, V. tapetis, V. splendidus. | [67,68,69] |
| Nigella sativa | V. parahaemolyticus | [70] |
| Origanum majorana | V. parahaemolyticus, V. alginolyticus | [21] |
| Artemisia absinthium, Zataria multiflora, Pulicaria gnaphalodes, Trachyspermum ammi, Cuminum cyminum | V. parahaemolyticus | [71] |
| Alpinia galanga, Zingiber officinale | V. cholerae | [72] |
| Origannum majorana, Cinnamomum verum | V. parahaemolyticus, V. cholerae | [73] |
| Protium heptaphyllum | V. parahaemolyticus | [74] |
| Abies alba, Apium graveolens, Artemisia dracunculus, A. herba alba, Cinnamomum camphora, C. cassia, C. zeylanicum, Citrus sinensis, C. cyminum, Curcuma longa, Cymbopogon martini, E. citriodora, E. dives, Laurus nobilis, Litsea citrata, Melaleuca alternifolia, Mentha × piperita, M. pulegium, P. crispum, Pogostemon cablin, Thymus zygis, Zingiber officinalis. | V. campbellii, V. parahaemolyticus | [75] |
| Clove, thyme, garlic | V. parahaemolyticus | [76] |
| Carum carvi, Coriandrum sativum L. | V. parahaemolyticus, V. alginolyticus, V. proteolyticus, V. furnisii, V. mimicus, V. natrigens, V. carhiaccae, V. fluvialis | [77] |
| Carum carvi | V. cholerae, V. vulnificus, V. parahaemolyticus, V. alginolyticus, V. furnisii, V. cincinnatiensis, V. proteolyticus, V. natrigens, V. mimicus, V. fluvialis, V. anguillarum, V. carrichariae, V. harveyii, V. diazotrophicus, V. tapetis, V. splendidus. | [53] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghannay, S.; Aouadi, K.; Kadri, A.; Snoussi, M. In Vitro and In Silico Screening of Anti-Vibrio spp., Antibiofilm, Antioxidant and Anti-Quorum Sensing Activities of Cuminum cyminum L. Volatile Oil. Plants 2022, 11, 2236. https://doi.org/10.3390/plants11172236
Ghannay S, Aouadi K, Kadri A, Snoussi M. In Vitro and In Silico Screening of Anti-Vibrio spp., Antibiofilm, Antioxidant and Anti-Quorum Sensing Activities of Cuminum cyminum L. Volatile Oil. Plants. 2022; 11(17):2236. https://doi.org/10.3390/plants11172236
Chicago/Turabian StyleGhannay, Siwar, Kaïss Aouadi, Adel Kadri, and Mejdi Snoussi. 2022. "In Vitro and In Silico Screening of Anti-Vibrio spp., Antibiofilm, Antioxidant and Anti-Quorum Sensing Activities of Cuminum cyminum L. Volatile Oil" Plants 11, no. 17: 2236. https://doi.org/10.3390/plants11172236