
Integrated Analysis of Transcriptome and Metabolome Reveals New Insights into the Formation of Purple Leaf Veins and Leaf Edge Cracks in Brassica juncea

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Comparison of Morphological Phenotypes among Mustards
2.2. Identification of Differential Expression of Genes with RNA-Seq
2.3. K-Means Cluster and Enrichment Analyses of DEGs Associated with the Formation of Different Phenotypes
2.4. Differential Accumulation of Metabolites in Four Mustards
2.5. Conjoint Analysis of Genes and Metabolites Related to Purple Leaf Veins and Notched Leaves
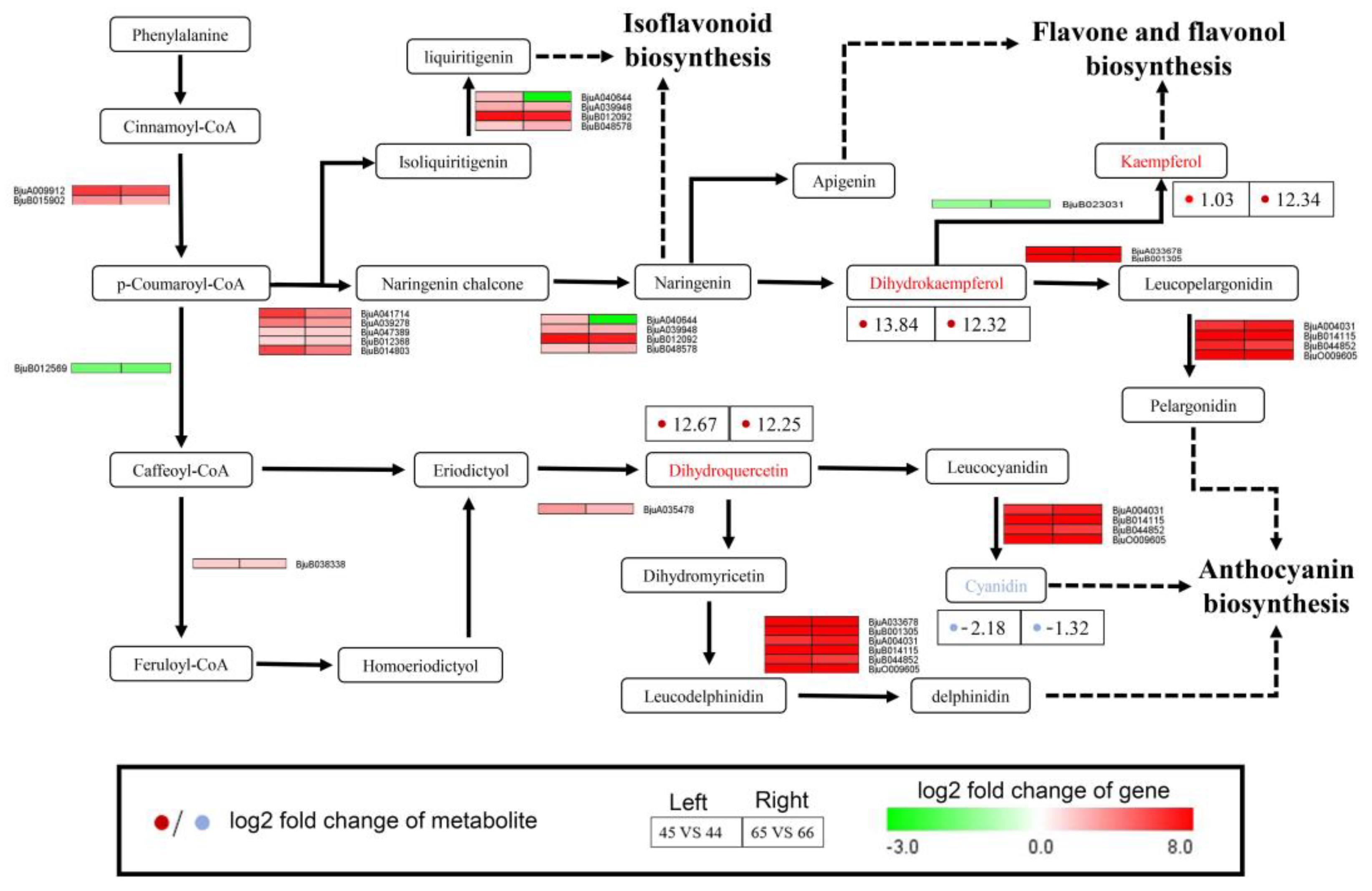
2.6. Integrating Related Genes and Metabolites in the Flavonoid Biosynthesis Pathway Provides Insights into Purple Vein Formation
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. RNA Sequencing and Data Analysis
4.3. Metabolic Analysis
4.4. Combined Analysis of DEGs and DAMs
4.5. qRT-PCR Validation of RNA-Seq Results
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fadhil, A.B.; Saleh, L.A.; Altamer, D.H. Production of biodiesel from non-edible oil, wild mustard (Brassica Juncea L.) seed oil through cleaner routes. Energy Sources Part A Recovery Util. Environ. Eff. 2020, 42, 1831–1843. [Google Scholar] [CrossRef]
- Voss-Fels, K.; Frisch, M.; Qian, L.; Kontowski, S.; Friedt, W.; Gottwald, S.; Snowdon, R.J. Subgenomic diversity patterns caused by directional selection in bread wheat gene pools. Plant Genome 2015, 8, 0013. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.; Li, S.; Dang, L.; Chai, W.T.; Li, P.L.; Wang, N.N. PL1 fusion gene: A novel visual selectable marker gene that confers tolerance to multiple abiotic stresses in transgenic tomato. Transgenic Res. 2012, 21, 1057–1070. [Google Scholar] [CrossRef] [PubMed]
- Heng, S.; Cheng, Q.; Zhang, T.; Liu, X.J.; Huang, H.; Yao, P.J.; Liu, Z.X.; Wan, Z.G.; Fu, T.D. Fine-mapping of the BjPur gene for purple leaf color in Brassica juncea. Theor. Appl. Genet. 2020, 133, 2989–3000. [Google Scholar] [CrossRef]
- Neill, S.O.; Gould, K.S.; Kilmartin, P.A.; Mitchell, K.A.; Markham, K.R. Antioxidant activities of red versus green leaves in Elatostema rugosum. Plant Cell Environ. 2002, 25, 539–547. [Google Scholar] [CrossRef]
- Oh, J.E.; Kim, Y.H.; Kim, J.H.; Kwon, Y.R.; Lee, H. Enhanced level of anthocyanin leads to increased salt tolerance in Arabidopsis PAP1-D plants upon sucrose treatment. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 79–88. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Qi, T.; Song, S.; Ren, Q.; Ren, G.C.; Wu, D.W.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.M.; et al. The Jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate Jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef]
- Carey, C.C.; Strahle, J.T.; Selinger, D.A.; Chandler, V.L. Mutations in the pale aleurone color1 regulatory gene of the Zea mays anthocyanin pathway have distinct phenotypes relative to the functionally similar TRANSPARENT TESTA GLABRA1 gene in Arabidopsis thaliana. Plant Cell 2004, 16, 450–464. [Google Scholar] [CrossRef]
- Hu, W.; Zhou, T.; Han, Z.; Tan, C.; Xing, Y.Z. Dominant complementary interaction between OsC1 and two tightly linked genes, Rb1 and Rb2, controls the purple leaf sheath in rice. Theor. Appl. Genet. 2020, 133, 2555–2566. [Google Scholar] [CrossRef]
- Chiu, L.W.; Zhou, X.; Burke, S.; Wu, X.L.; Prior, R.L.; Li, L. The purple cauliflower arises from activation of a MYB transcription factor. Plant Physiol. 2010, 154, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Song, H.Y.; Lee, M.; Han, C.T.; Lee, J.; Kim, H.; Park, J.I.; Nou, I.S.; Kim, S.J.; Hur, Y. Purple Brassica oleracea var. capitata F. rubra is due to the loss of BoMYBL2–1 expression. BMC Plant Biol. 2018, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Wu, J.; Zheng, S.; Cheng, F.; Liu, B.; Liang, J.L.; Cui, Y.; Wang, X.W. Anthocyanin profile characterization and quantitative trait locus mapping in zicaitai (Brassica rapa L. ssp. chinensis var. purpurea). Mol. Breed. 2015, 35, 113. [Google Scholar] [CrossRef]
- Tang, Q.; Tian, M.; An, G.; Zhang, W.Y.; Chen, J.J.; Yan, C.H. Rapid identification of the purple stem (Ps) gene of Chinese kale (Brassica oleracea var. alboglabra) in a segregation distortion population by bulked segregant analysis and RNA sequencing. Mol. Breed. 2017, 37, 153. [Google Scholar] [CrossRef]
- Runions, A.; Tsiantis, M.; Prusinkiewicz, P. A common developmental program can produce diverse leaf shapes. New Phytol. 2017, 216, 401–418. [Google Scholar] [CrossRef]
- Vogel, S. Leaves in the lowest and highest winds: Temperature, force and shape. New Phytol. 2009, 183, 13–26. [Google Scholar] [CrossRef]
- Sicard, A.; Thamm, A.; Marona, C.; Lee, Y.W.; Wahl, V.; Stinchcombe, J.R.; Wright, S.I.; Kappel, C.; Lenhard, M. Repeated evolutionary changes of leaf morphology caused by mutations to a homeobox gene. Curr. Biol. 2014, 24, 1880–1886. [Google Scholar] [CrossRef]
- Rowland, S.D.; Zumstein, K.; Nakayama, H.; Cheng, Z.Z.; Flores, A.M.; Chitwood, D.H.; Maloof, J.N.; Sinha, N.R. Leaf shape is a predictor of fruit quality and cultivar performance in tomato. New Phytol. 2020, 226, 851–865. [Google Scholar] [CrossRef]
- Li, Z.; Li, B.; Shen, W.H.; Huang, H.; Dong, A.W. TCP transcription factors interact with AS2 in the repression of class-I KNOX genes in Arabidopsis thaliana. Plant J. 2012, 71, 99–107. [Google Scholar] [CrossRef]
- Bar, M.; Ori, N. Compound leaf development in model plant species. Curr. Opin. Plant Biol. 2015, 23, 61–69. [Google Scholar] [CrossRef]
- Sarvepalli, K.; Gupta, M.D.; Challa, K.R.; Nath, U. Molecular cartography of leaf development-role of transcription factors. Curr. Opin. Plant Biol. 2019, 47, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.; Ori, N. Mechanisms of cross talk between gibberellin and other hormones. Plant Physiol. 2007, 144, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Sarvepalli, K.; Nath, U. Interaction of TCP4-mediated growth module with phytohormones. Plant Signal. Behav. 2011, 6, 1440–1443. [Google Scholar] [CrossRef]
- Hibara, K.; Karim, M.R.; Takada, S.; Taoka, K.I.; Furutani, M.; Aida, M.; Tasaka, M. Arabidopsis CUP-SHAPED COTYLEDON3 regulates postembryonic shoot meristem and organ boundary formation. Plant Cell 2006, 18, 2946–2957. [Google Scholar] [CrossRef] [PubMed]
- Nikovics, K.; Blein, T.; Peaucelle, A.; Ishida, T.; Morin, H.; Aida, M.; Laufs, P. The balance between the MIR164A and CUC2 genes controls leaf margin serration in Arabidopsis. Plant Cell 2006, 18, 2929–2945. [Google Scholar] [CrossRef]
- Hasson, A.; Plessis, A.; Blein, T.; Adroher, B.; Grigg, S.; Tsiantis, M.; Boudaoud, A.; Damerval, C.; Laufs, P. Evolution and diverse roles of the CUP-SHAPED COTYLEDON genes in Arabidopsis leaf development. Plant Cell 2011, 23, 54–68. [Google Scholar] [CrossRef]
- Saddic, L.A.; Huvermann, B.; Bezhani, S.; Su, Y.H.; Winter, C.M.; Kwon, C.S.; Collum, R.P.; Wagner, D. The LEAFY target LMI1 is a meristem identity regulator and acts together with LEAFY to regulate expression of CAULIFLOWER. Development 2006, 133, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Hajheidari, M.; Wang, Y.; Bhatia, N.; Vuolo, F.; Franco-Zorrilla, J.M.; Karady, M.; Mentink, R.A.; Wu, A.H.; Oluwatobi, B.R.; Muller, b.; et al. Autoregulation of RCO by low-affinity binding modulates cytokinin action and shapes leaf diversity. Curr. Biol. 2019, 29, 4183–4192. [Google Scholar] [CrossRef]
- Gupta, S.; Rosenthal, D.M.; Stinchcombe, J.R.; Baucom, R.S. The remarkable morphological diversity of leaf shape in sweet potato (Ipomoea batatas): The influence of genetics, environment, and G × E. New Phytol. 2020, 225, 2183–2195. [Google Scholar] [CrossRef]
- Ni, X.; Huang, J.; Ali, B.; Zhou, W.J.; Zhao, J.Y. Genetic analysis and fine mapping of the LOBED-LEAF 1 (BnLL1) gene in rapeseed (Brassica napus L.). Euphytica 2015, 204, 29–38. [Google Scholar] [CrossRef]
- Ni, X.; Liu, H.; Huang, J.; Zhao, J.Y. LMI1-like genes involved in leaf margin development of Brassica napus. Genetica 2017, 145, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Zhang, H.; Yang, Q.; Meng, Q.W.; Han, S.Q.; Nwafor, C.C.; Khan, M.H.U.; Fan, C.C.; Zhou, Y.M. Promoter variations in a homeobox gene, BnA10.LMI1, determine lobed leaves in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2018, 131, 2699–2708. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Fernie, A.R.; Luo, J. Exploring the diversity of plant metabolism. Trends Plant Sci. 2019, 24, 83–98. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, H.; Kambe, Y.; Ohshio, M.; Kunihiro, A.; Tanaka, Y.; Suzuki, T.; Nakamura, Y.; Morita, A.; Ikka, T. Integrated metabolome and transcriptome analyses reveal etiolation-induced metabolic changes leading to high amino acid contents in a light-sensitive Japanese albino tea cultivar. Front. Plant Sci. 2021, 11, 2194. [Google Scholar] [CrossRef] [PubMed]
- Szymański, J.; Bocobza, S.; Panda, S.; Sonawane, P.; Cárdenas, P.D.; Lashbrooke, J.; Kamble, A.; Shahaf, N.; Meir, S.; Bovy, A. Analysis of wild tomato introgression lines elucidates the genetic basis of transcriptome and metabolome variation underlying fruit traits and pathogen response. Nat. Genet. 2020, 52, 1111–1121. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, S. Biosynthesis and regulation of phenylpropanoids in plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Deng, Y.; Li, C.; Li, H.; Lu, S. Identification and characterization of flavonoid biosynthetic enzyme genes in Salvia miltiorrhiza (Lamiaceae). Molecules 2018, 23, 1467. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, C.; Duan, L.; Luan, Q.Q.; Li, J.L.; Yang, A.G.; Qi, X.Q.; Ren, Z.H. CsMYB60 is a key regulator of flavonols and proanthocyanidans that determine the colour of fruit spines in cucumber. J. Exp. Bot. 2019, 70, 69–84. [Google Scholar] [CrossRef]
- Peterson, J.J.; Dwyer, J.T.; Beecher, G.R.; Bhagwat, S.A.; Gebhardt, S.E.; Haytowitz, D.B.; Holden, J.M. Flavanones in oranges, tangerines (mandarins), tangors, and tangelos: A compilation and review of the data from the analytical literature. J. Food Compos. Anal. 2006, 19, S66–S73. [Google Scholar] [CrossRef]
- Goulas, V.; Manganaris, G.A. Exploring the phytochemical content and the antioxidant potential of Citrus fruits grown in Cyprus. Food Chem. 2012, 131, 39–47. [Google Scholar] [CrossRef]
- Shi, J.; Simal-Gandara, J.; Mei, J.; Ma, W.J.; Peng, Q.H.; Shi, Y.L.; Xu, Q.; Lin, Z.; Lv, H.P. Insight into the pigmented anthocyanins and the major potential co-pigmented flavonoids in purple-coloured leaf teas. Food Chem. 2021, 363, 130278. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Souza, M.A.; Peres, L.E.P.; Freschi, J.R.; Purgatto, E.; Lajolo, F.M.; Hassimotto, N.M. Changes in flavonoid and carotenoid profiles alter volatile organic compounds in purple and orange cherry tomatoes obtained by allele introgression. J. Sci. Food. Agric. 2020, 100, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pan, D.; Liang, M.; Abubakar, Y.S.; Li, J.; Lin, J.; Chen, S.; Chen, W. Regulation of anthocyanin biosynthesis in purple leaves of zijuan tea (Camellia sinensis var. kitamura). Int. J. Mol. Sci. 2017, 18, 833. [Google Scholar] [CrossRef] [PubMed]
- Chaves-Silva, S.; Dos Santos, A.L.; Chalfun-Júnior, A.; Zhao, J.; Peres, L.E.P.; Benedito, V.A. Understanding the genetic regulation of anthocyanin biosynthesis in plants–tools for breeding purple varieties of fruits and vegetables. Phytochemistry 2018, 153, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, P.; Zheng, Y.; Gu, M.Y.; Lin, X.Y.; Wang, S.Y.; Jin, S.; Ye, N.X. Comparison of metabolome and transcriptome of flavonoid biosynthesis pathway in a purple-leaf tea germplasm Jinmingzao and a green-leaf tea germplasm Huangdan reveals their relationship with genetic mechanisms of color formation. Int. J. Mol. Sci. 2020, 21, 4167. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.C.; Giusti, M.M. Anthocyanins. Adv. Nutr. 2015, 6, 620–622. [Google Scholar] [CrossRef]
- Giusti, M.M.; Rodríguez-Saona, L.E.; Wrolstad, R.E. Molar absorptivity and color characteristics of acylated and non-acylated pelargonidin-based anthocyanins. J. Agric. Food Chem. 1999, 47, 4631–4637. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Acylated anthocyanins from edible sources and their applications in food systems. Biochem. Eng. J. 2003, 14, 217–225. [Google Scholar] [CrossRef]
- Lai, Y.S.; Li, S.; Tang, Q.; Li, H.S.; Chen, S.X.; Li, P.W.; Xu, J.Y.; Xu, Y.; Guo, X. The dark-purple tea cultivar ‘Ziyan’ accumulates a large amount of delphinidin-related anthocyanins. J. Agric. Food Chem. 2016, 64, 2719–2726. [Google Scholar] [CrossRef]
- Noda, N.; Aida, R.; Kishimoto, S.; Ishiguro, K.; Fukuchi-Mizutani, M.; Tanaka, Y.; Ohmiya, A. Genetic engineering of novel bluer-colored chrysanthemums produced by accumulation of delphinidin-based anthocyanins. Plant Cell Physiol. 2013, 54, 1684–1695. [Google Scholar] [CrossRef] [Green Version]
- Chitwood, D.H.; Sinha, N.R. Evolutionary and environmental forces sculpting leaf development. Curr. Biol. 2016, 26, R297–R306. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, D.; Wang, X.; Ji, C.; Cheng, F.; Liu, B.; Hu, Z.; Chen, S.; Pental, D.; Ju, Y.; et al. The genome sequence of allopolyploid Brassica juncea and analysis of differential homoeolog gene expression influencing selection. Nat. Genet. 2016, 48, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Götz, S. Blast2GO: A comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genom. 2008, 2008, 619832. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirekawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36, D480–D484. [Google Scholar] [CrossRef]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [Green Version]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for visualization and analysis of biological networks. Humana Press 2011, 696, 291–303. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Raw Reads | Clean Reads | Q30 (%) | GC Content (%) | Unique Mapped |
|---|---|---|---|---|---|
| 44-1 | 52,764,138 | 51,838,932 | 90.47 | 47.65 | 41,712,627 (80.47%) |
| 44-2 | 57,898,070 | 56,684,510 | 92.36 | 47.71 | 46,074,374 (81.28%) |
| 44-3 | 48,686,530 | 47,839,794 | 92.77 | 47.59 | 38,895,911 (81.30%) |
| 45-1 | 59,411,784 | 58,048,268 | 92.23 | 47.75 | 47,126,857 (81.19%) |
| 45-2 | 59,141,800 | 57,661,630 | 92.14 | 47.56 | 45,972,841 (79.73%) |
| 45-3 | 52,856,618 | 51,892,220 | 92.01 | 47.64 | 41,742,258 (80.44%) |
| 65-1 | 59,045,222 | 57,784,616 | 92.21 | 47.37 | 46,641,623 (80.72%) |
| 65-2 | 52,647,580 | 51,702,172 | 91.46 | 47.61 | 41,756,075 (80.76%) |
| 65-3 | 57,451,232 | 56,518,720 | 92.24 | 47.45 | 45,650,258 (80.77%) |
| 66-1 | 54,027,590 | 53,162,134 | 92.64 | 47.86 | 42,963,193 (80.82%) |
| 66-2 | 56,428,334 | 55,492,792 | 92.28 | 47.8 | 44,569,467 (80.32%) |
| 66-3 | 51,296,644 | 50,462,792 | 92.09 | 47.62 | 40,610,737 (80.48%) |
| Metabolome ID | Compounds | Class I | Class II | Log2FC (45 vs. 44) | Log2FC (65 vs. 66) |
|---|---|---|---|---|---|
| CWJP002833 | Kaempferol-di-O-malonyl-glucoside-O-malonyl-diglucoside | Flavonoids | Flavonols | 13.84 | 1.64 |
| Hmgp006596 | Cyclobrassinin | Alkaloids | Plumerane | 1.128 | 1.64 |
| Hmyn007168 | LysoPG(16:0) | Lipids | Glycerol ester | 1.32 | 1.09 |
| Lmmp002995 | Quercetin glu-malonyl-glucoside2 | Flavonoids | Flavonols | 12.68 | 12.25 |
| Lmyn001269 | Kaempferol 3-O-β-D-sophoroside | Flavonoids | Flavonols | 1.03 | 12.34 |
| mws0126 | 1-Stearoyl-sn-glycero-3-phosphocholine | Lipids | LPC | 1.156 | 1.72 |
| mws0183 | Protocatechuic acid | Flavonoids | Flavanols | 1.63 | 1.70 |
| mws0639 | 2,3-Dihydroxybenzoic acid | Organic acids | Organic acids | 1.18 | 1.10 |
| mws1138 | Betanin | Alkaloids | Alkaloids | 14.19 | 13.17 |
| pmb0876 | LysoPE 16:0 | Lipids | LPE | 1.68 | 1.74 |
| pmb2936 | Disinapoyl hexoside | Phenolic acids | Phenolic acids | 1.76 | 12.55 |
| pmb2940 | 1-O-β-D-Glucopyranosyl sinapate | Phenolic acids | Phenolic acids | 1.66 | 1.97 |
| pmd0136 | LysoPC 18:0 | Lipids | LPC | 1.05 | 1.73 |
| pmn001409 | Plantainoside A | Phenolic acids | Phenolic acids | 1.32 | 1.06 |
| pmp001277 | 3-{(2-Aminoethoxy)(hydroxy)phosphoryl]oxy}-2-hydroxypropyl palmitate | Alkaloids | Alkaloids | 1.69 | 1.68 |
| pmp001286 | LysoPC(18:0) | Lipids | LPC | 1.15 | 1.67 |
| Hmpp002612 | Luteolin-7-O-β-D-gentiobioside | Flavonoids | Flavonoid | −2.19 | −1.31 |
| Lmbp002592 | Kaempferol-3,7-di-O-β-D-glucopyranoside | Flavonoids | Flavonols | −2.34 | −1.82 |
| mws0170 | Cyanidin chloride | Flavonoids | Anthocyanins | −2.26 | −2.32 |
| pme0256 | Xanthine | Nucleotides and derivatives | Nucleotides and derivatives | −1.01 | −1.09 |
| pme0516 | Inositol | Others | Saccharides and Alcohols | −1.66 | −2.42 |
| Metabolome ID | Compounds | Class I | Class II | Log2FC (45 vs. 44) | Log2FC (45 vs. 66) |
|---|---|---|---|---|---|
| Cmln000394 | 4-(Methylthio)-3-OH-butyl glucosinolate (glucoraphanin) | Others | Glucosinolates | 1.41 | 1.37 |
| CMLN000400 | 4-Methylsulfinyl-3-butenyl thioglucoside (glucoraphenin) | Others | Glucosinolates | 1.83 | 1.33 |
| GQ512003 | Fer-agmatine | Alkaloids | Phenolamine | 1.25 | 3.54 |
| Hmgp006596 | Cyclobrassinin | Alkaloids | Plumerane | 1.13 | 1.86 |
| mws0997 | Petunidin 3-O-glucoside | Flavonoids | Anthocyanins | 2.00 | 1.04 |
| pmb0489 | N-hexosyl-p-coumaroyl putrescine | Alkaloids | Phenolamine | 3.60 | 2.75 |
| pmb0490 | N-p-coumaroyl putrescine | Alkaloids | Phenolamine | 2.68 | 2.44 |
| pmb0494 | N-sinapoyl putrescine | Alkaloids | Phenolamine | 2.17 | 1.98 |
| pmb0496 | N-feruloyl agmatine | Alkaloids | Phenolamine | 1.15 | 3.72 |
| pmb0608 | Chrysoeriol O-malonylhexoside | Flavonoids | Flavonoid | 1.21 | 1.10 |
| pmb3081 | Glucarate O-phosphoric acid | Others | Saccharides and alcohols | 1.27 | 1.70 |
| pme0008 | L-citrulline | Amino acids and derivatives | Amino acids and derivatives | 1.18 | 1.27 |
| pme2596 | 4-Pyridoxic acid | Others | Vitamin | 1.26 | 1.30 |
| pme3388 | H-homoArg-OH | Amino acids and derivatives | Amino acids and derivatives | 2.30 | 1.84 |
| Rfmb319 | Pipecolic acid | Amino acids and derivatives | Amino acids and derivatives | 1.03 | 1.56 |
| Cmyn001733 | 1-Methylpropyl glucosinolate | Others | Glucosinolates | −4.29406 | −3.09 |
| CWJP002007 | Kaempferol-3-O-feruloyl-sophoroside-7-oglucoside | Flavonoids | Flavonols | −1.57628 | −1.29 |
| Hmbn002228 | kaeperferol-3-O-[2-O-(6-O-E-feruloyl)-β-D-glucopyranosyl]-β-galactopyranoside | Others | Others | −1.14448 | −2.39 |
| Lmhn002423 | vnilloylmalic acid | Phenolic acids | Phenolic acids | −4.00812 | −2.29 |
| Lnrp102522 | Kaempferol glc-glc-rha | Flavonoids | Flavonols | −3.30626 | −1.23 |
| mws1212 | Methyl ferulate | Phenolic acids | Phenolic acids | −1.32806 | −1.24 |
| mws1550 | S-allyl-L-cysteine | Amino acids and derivatives | Amino acids and derivatives | −1.65983 | −1.36 |
| pmb0382 | O-feruloyl 4-hydroxylcoumarin | Lignans and Coumarins | Coumarins | −2.10723 | −3.09 |
| pmb2620 | 3,4-Dimethoxycinnamic acid | Phenolic acids | Phenolic acids | −1.35293 | −1.09 |
| pmb3072 | 3-O-p-coumaroyl shikimic acid O-hexoside | Phenolic acids | Phenolic acids | −1.81168 | −2.06 |
| pme2914 | 3-Hydroxy-3-methylpentane-1,5-dioic acid | Amino acids and derivatives | Amino acids and derivatives | −1.70006 | −1.99 |
| pme3382 | N-acetylthreonine | Amino acids and derivatives | Amino acids and derivatives | −1.98759 | −1.27 |
| Zmhn001375 | Vanillic acid-C-glucoside | Phenolic acids | Phenolic acids | −1.91953 | −1.30 |
| Zmhn001883 | Vanillic acid glycoside | Phenolic acids | Phenolic acids | −1.64013 | −1.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Yang, D.; Hu, Y.; Njogu, M.K.; Qian, J.; Jia, L.; Yan, C.; Li, Z.; Wang, X.; Wang, L. Integrated Analysis of Transcriptome and Metabolome Reveals New Insights into the Formation of Purple Leaf Veins and Leaf Edge Cracks in Brassica juncea. Plants 2022, 11, 2229. https://doi.org/10.3390/plants11172229
Zhang K, Yang D, Hu Y, Njogu MK, Qian J, Jia L, Yan C, Li Z, Wang X, Wang L. Integrated Analysis of Transcriptome and Metabolome Reveals New Insights into the Formation of Purple Leaf Veins and Leaf Edge Cracks in Brassica juncea. Plants. 2022; 11(17):2229. https://doi.org/10.3390/plants11172229
Chicago/Turabian StyleZhang, Kaijing, Dekun Yang, Yuchao Hu, Martin Kagiki Njogu, Jingjing Qian, Li Jia, Congsheng Yan, Ziang Li, Xing Wang, and Liping Wang. 2022. "Integrated Analysis of Transcriptome and Metabolome Reveals New Insights into the Formation of Purple Leaf Veins and Leaf Edge Cracks in Brassica juncea" Plants 11, no. 17: 2229. https://doi.org/10.3390/plants11172229