
Physiological and Transcriptome Analyses of CaCl2 Treatment to Alleviate Chilling Injury in Pineapple

Abstract
:1. Introduction
2. Results
2.1. Physiological and Biochemical Results
2.1.1. IB Appearance and Electrical Conductivity
2.1.2. Membrane ROS Production and Membrane Lipid Peroxidation
2.1.3. Ascorbic Acid (ASA) and Glutathione (GSH) Contents
2.1.4. Phenolic Metabolism and Active Oxidase
2.1.5. Antioxidant Enzyme Activities
2.2. Transcriptome Results
2.2.1. Raw Data Quality Assessment
2.2.2. Overall Transcriptome Quality Assessment
2.2.3. Screening Results of DEGs
2.2.4. DEGs, GO, and KEGG Pathway Enrichment Analysis
2.2.5. Screening and Differential Expressions of Oxidation and Antioxidation-Related Genes
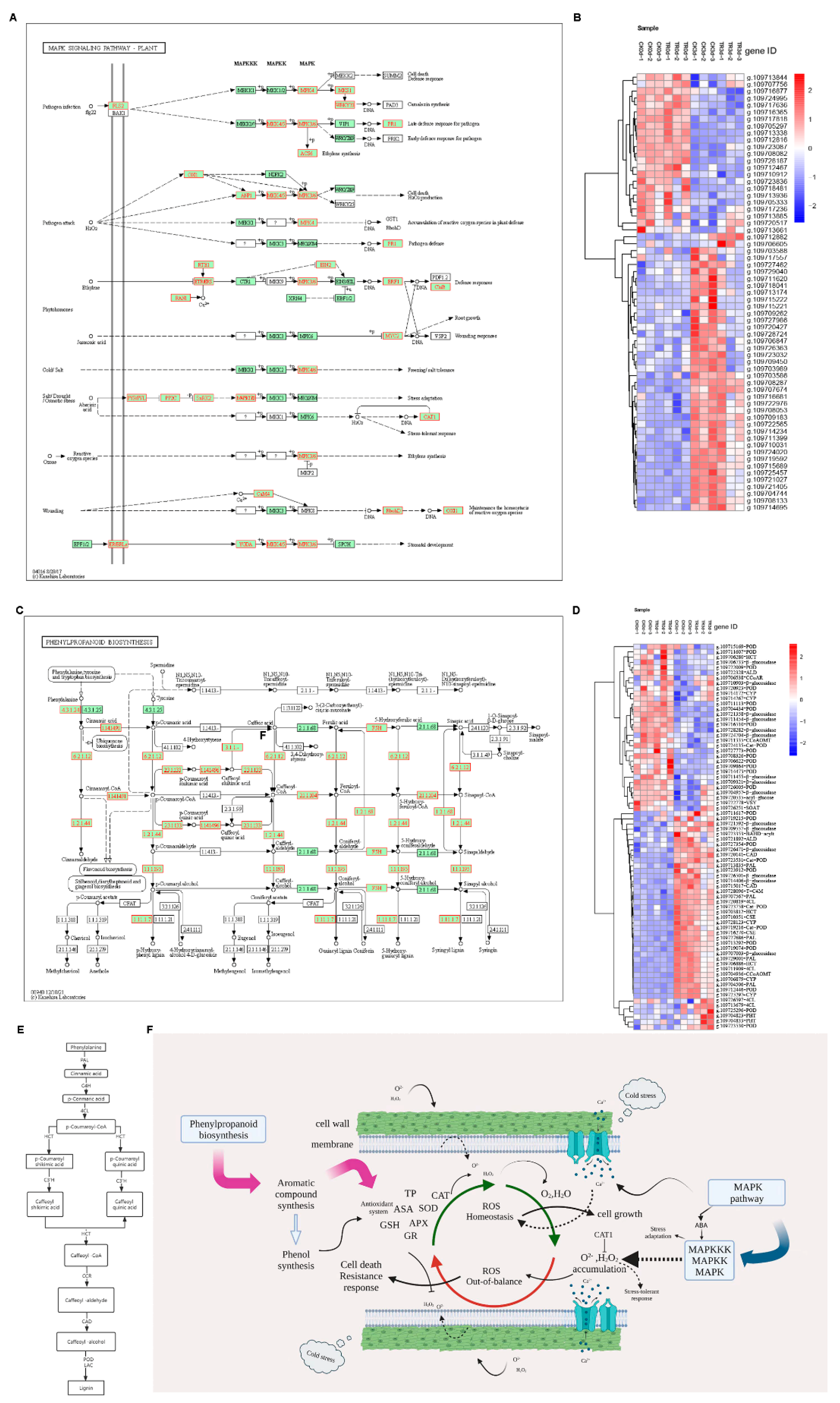
2.2.6. Phenylpropanoid Biosynthesis and MAPK Pathway
2.2.7. Heatmap Analysis of Related Pathways
2.2.8. Transcription Factor Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Optimum Concentration Obtained
4.2.1. IB Appearance
4.2.2. Electrical Conductivity Measurement
4.3. Determination of Physiological and Biochemical Indexes
4.3.1. Assays of ROS Production and Membrane Lipid Peroxidation MDA Contents
4.3.2. Determination of Total Phenolic Content
4.3.3. Assessments of Polyphenol Oxidase (PPO) and Peroxidase (POD) Activities
4.3.4. Assessments of Ascorbic Acid (ASA) and Glutathione (GSH) Contents
4.3.5. Assessment of Antioxidant Enzyme Activities
4.4. Transcriptome Data Analysis
4.4.1. Transcriptome Data Quality Assessment
4.4.2. GO and KEGG Enrichment Analysis
4.4.3. Screening of Related DEGs, Analysis of Related Pathways, and Transcription Factor Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hong, K.; Xu, H.; Wang, J.; Zhang, L.; Hu, H.; Jia, Z.; Gu, H.; He, Q.; Gong, D. Quality changes and internal browning developments of summer pineapple fruit during storage at different temperatures. Sci. Hortic. 2013, 151, 68–74. [Google Scholar] [CrossRef]
- Ali, M.M.; Hashim, N.; Abd Aziz, S.; Lasekan, O. Pineapple (Ananas comosus): A comprehensive review of nutritional values, volatile compounds, health benefits, and potential food products. Food Res. Int. 2020, 137, 109675. [Google Scholar]
- Zhao, J.; Quan, P.; Liu, H.; Li, L.; Qi, S.; Zhang, M.; Xing, L. Transcriptomic and metabolic analyses provide new insights into the apple fruit quality decline during long-term cold storage. J. Agric. Food Chem. 2020, 68, 4699–4716. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Tian, M.; Huang, W.; Luo, S.; Hu, H.; Zhang, Y.; Li, P. Physiological and transcriptomic analysis of ‘Whangkeumbae’ pear core browning during low-temperature storage. Gene Expr. Patterns 2020, 36, 119113. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, X.; Zhou, Q.; Cheng, S.; Wei, B.; Ji, S. Low temperature conditioning alleviates peel browning by modulating energy and lipid metabolisms of ‘Nanguo’ pears during shelf life after cold storage. Postharvest Biol. Technol. 2017, 131, 10–15. [Google Scholar] [CrossRef]
- Si, J.; Fan, Y.Y.; Liu, Z.L.; Wei, W.; Xiao, X.M.; Yang, Y.Y.; Chen, J.Y. Comparative transcriptomic analysis reveals the potential mechanism of hot water treatment alleviated-chilling injury in banana fruit. Food Res. Int. 2022, 157, 111296. [Google Scholar] [CrossRef]
- Rohrbach, K.G.; Johnson, M.W. Pests, diseases and weeds. In The Pineapple: Botany, Production and Uses; CABI Publishing: New York, NY, USA, 2003; pp. 203–251. [Google Scholar]
- Wang, A.; Li, J.; Al-Huqail, A.A.; Al-Harbi, M.S.; Ali, E.F.; Wang, J.; Eissa, M.A. Mechanisms of chitosan nanoparticles in the regulation of cold stress resistance in banana plants. Nanomaterials 2021, 11, 2670. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, W.; Cao, J.; Li, Y. Effect of chilling temperatures on physiological properties, phenolic metabolism and antioxidant level accompanying pulp browning of peach during cold storage. Sci. Hortic. 2019, 255, 175–182. [Google Scholar] [CrossRef]
- Liu, H.; He, H.; Liu, C.; Wang, C.; Qiao, Y.; Zhang, B. Changes of Sensory Quality, Flavor-Related Metabolites and Gene Expression in Peach Fruit Treated by Controlled Atmosphere (CA) under Cold Storage. Int. J. Mol. Sci. 2022, 23, 7141. [Google Scholar] [CrossRef]
- Zhao, Y.; Song, C.; Brummell, D.A.; Qi, S.; Lin, Q.; Bi, J.; Duan, Y. Salicylic acid treatment mitigates chilling injury in peach fruit by regulation of sucrose metabolism and soluble sugar content. Food Chem. 2021, 358, 129867. [Google Scholar] [CrossRef]
- Per, T.S.; Khan, N.A.; Reddy, P.S.; Masood, A.; Hasanuzzaman, M.; Khan, M.I.R.; Anjum, N.A. Approaches in modulating proline metabolism in plants for salt and drought stress tolerance: Phytohormones, mineral nutrients and transgenics. Plant Physiol. Biochem. 2017, 115, 126–140. [Google Scholar] [PubMed]
- Zhang, Q.; Liu, Y.; He, C.; Zhu, S. Postharvest exogenous application of abscisic acid reduces internal browning in pineapple. J. Agric. Food Chem. 2015, 63, 5313–5320. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Deng, W.; Hu, N.; Chen, N.; Li, Z. Metabolite and transcriptomic analysis reveals metabolic and regulatory features associated with Powell orange pulp deterioration during room temperature and cold storage. Postharvest Biol. Technol. 2016, 112, 75–86. [Google Scholar] [CrossRef]
- Zhu, A.; Li, W.; Ye, J.; Sun, X.; Ding, Y.; Cheng, Y.; Deng, X. Microarray expression profiling of postharvest Ponkan mandarin (Citrus reticulata) fruit under cold storage reveals regulatory gene candidates and implications on soluble sugars metabolism. J. Integr. Plant Biol. 2011, 53, 358–374. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yin, X.-R.; Zhang, B.; Grierson, D.; Xu, C.J.; Chen, K.S. Transcriptomic and Metabolic Analyses Provide New Insights into Chilling Injury in Peach Fruit. Plant Cell Environ. 2017, 40, 1531–1551. [Google Scholar] [CrossRef]
- Wang, J.; Dong, S.; Jiang, Y.; He, H.; Ji, S. Influence of long-term cold storage on phenylpropanoid and soluble sugar metabolisms accompanied with peel browning of ′nanguo′ pears during subsequent shelf life. Sci. Hortic. 2020, 260, 108888. [Google Scholar] [CrossRef]
- Sivankalyani, V.; Sela, N.; Feygenberg, O.; Zemach, H.; Maurer, D.; Alkan, N. Transcriptome dynamics in mango fruit peel reveals mechanisms of chilling stress. Front. Plant Sci. 2016, 7, 1579. [Google Scholar] [CrossRef]
- Yuan, P.; Yang, T.; Poovaiah, B.W. Calcium Signaling-Mediated Plant Response to Cold Stress. Int. J. Mol. Sci. 2018, 19, 3896. [Google Scholar] [CrossRef]
- Falchi, R.; D’Agostin, E.; Mattiello, A.; Coronica, L.; Spinelli, F.; Costa, G.; Vizzotto, G. ABA regulation of calcium-related genes and bitter pit in apple. Postharvest Biol. Technol. 2017, 132, 1–6. [Google Scholar] [CrossRef]
- Gilroy, S.; Blowers, D.; Trewavas, A. Calcium: A regulation system emerges in plant cells. Development 1987, 100, 181–184. [Google Scholar] [CrossRef]
- Wei, D.; Zhao, X.h. Calcium maintained higher quality and enhanced resistance against chilling stress by regulating enzymes in reactive oxygen and biofilm metabolism of Chinese winter jujube fruit. J. Food Biochem. 2020, 44, e13161. [Google Scholar] [CrossRef] [PubMed]
- Nor, S.M.; Ding, P. Trends and Advances in Edible Biopolymer Coating for Tropical Fruit: A Review. Food Res. Int. 2020, 134, 109208. [Google Scholar] [CrossRef] [PubMed]
- Nukuntornprakit, O.A.; Luengwilai, K.; Siriphanich, J. Chilling injury in pineapple fruit is related to mitochondrial antioxidative metabolism. Postharvest Biol. Technol. 2020, 170, 111330. [Google Scholar] [CrossRef]
- Shi, H.; Ye, T.; Zhong, B.; Liu, X.; Chan, Z. Comparative proteomic and metabolomic analyses reveal mechanisms of improved cold stress tolerance in bermudagrass (Cynodon dactylon (L.) Pers.) by exogenous calcium. J. Integr. Plant Biol. 2014, 56, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, I. Calcium in plant senescence and fruit ripening. Plant Cell Environ. 1984, 7, 477–489. [Google Scholar]
- Conway, W.S.; Sams, C.E.; Wang, C.Y.; Abbott, J.A. Additive effects of postharvest calcium and heat treatment on reducing decay and maintaining quality in apples. J. Am. Soc. Hortic. Sci. 1994, 119, 49–53. [Google Scholar] [CrossRef]
- Youryon, P.; Wongsaree, C.; Mcglasson, B.; Glahan, S.; Kanlayanarat, S. Alleviation of internal browning in pineapple fruit by peduncle infiltration with solutions of calcium chloride or strontium chloride under mild chilling storage. Int. Food Res. J. 2013, 20, 239–246. [Google Scholar]
- Youryon, P.; Supapvanich, S. Effects of Preharvest Boron, Calcium Sulfate Treatment and Postharvest Calcium Chloride Peduncle Infiltration on Chilling Injury Alleviation of Queen Pineapple cv. Sawi Fruit. Curr. Appl. Sci. Technol. 2011, 21, 456–466. [Google Scholar]
- Habibi, F.; Ramezanian, A.; Guillén, F.; Martínez-Romero, D.; Serrano, M.; Valero, D. Susceptibility of blood orange cultivars to chilling injury based on antioxidant system and physiological and biochemical responses at different storage temperatures. Foods 2020, 9, 1609. [Google Scholar] [CrossRef]
- Om-Arun, N.; Siriphanich, J. Hydrogen peroxide and ascorbic acid contents, superoxide dismutase and catalase activities in Smooth Cayenne and Queen pineapples during cold storage. V Int. Postharvest Symp. 2004, 682, 611–616. [Google Scholar] [CrossRef]
- Nukuntornprakit, O.-A.; Chanjirakul, K.; van Doorn, W.G.; Siriphanich, J. Chilling injury in pineapple fruit: Fatty acid composition and antioxidant metabolism. Postharvest Biol. Technol. 2015, 99, 20–26. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Y.; Zhang, J. Advances in the research on the AsA-GSH cycle in horticultural crops. Front. Agric. China 2010, 4, 84–90. [Google Scholar]
- Hou, Y.; Li, Z.; Zheng, Y.; Jin, P. Effects of CaCl2 treatment alleviates chilling injury of loquat fruit (Eribotrya japonica) by modulating ROS homeostasis. Foods 2021, 10, 1662. [Google Scholar] [CrossRef] [PubMed]
- Youryon, P.; Supapvanich, S.; Wongs-Aree, C. Internal browning alleviation of Queen pineapple cv. Sawi under cold storage using salicylic acid or abscisic acid peduncle infiltration. Hortic. Sci. Biotechnol. 2019, 94, 744–752. [Google Scholar] [CrossRef]
- Raimbault, A.K.; Marie Alphonsine, P.A.; Horry, J.P.; Francois Haugrin, M.; Romuald, K.; Soler, A. Polyphenol oxidase and peroxidase expression in four pineapple varieties (Ananas comosus L.) after a chilling injury. J. Agric. Food Chem. 2011, 59, 342–348. [Google Scholar] [CrossRef]
- You, J.; Chan, Z. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, M.; Ye, B.; Liu, L.; Ji, S. Calcium ion improves cold resistance of green peppers (Capsicum annuum L.) by regulating the activity of protective enzymes and membrane lipid composition. Sci. Hortic. 2021, 277, 109789. [Google Scholar]
- Chen, C.; Zhang, H.; Dong, C.; Ji, H.; Zhang, X.; Li, L.; Ban, Z.; Zhang, N.; Xue, W. Effect of ozone treatment on the phenylpropanoid biosynthesis of postharvest strawberries. RSC Adv. 2019, 9, 25429–25438. [Google Scholar] [CrossRef]
- Liu, Y.; He, C. A review of redox signaling and the control of MAP kinase pathway in plants. Redox Biol. 2017, 11, 192–204. [Google Scholar]
- Zhang, Z.; Huber, D.J.; Qu, H.; Yun, Z.; Wang, H.; Huang, Z.; Huang, H.; Jiang, Y. Enzymatic browning and antioxidant activities in harvested litchi fruit as influenced by apple polyphenols. Food Chem. 2015, 171, 191–199. [Google Scholar] [CrossRef]
- Li, W.; Liang, H.; Zhang, M.W.; Zhang, R.F.; Deng, Y.Y.; Wei, Z.C.; Tang, X.J. Phenolic profiles and antioxidant activity of litchi (Litchi Chinensis Sonn.) fruit pericarp from different commercially available cultivars. Molecules 2012, 17, 14954–14967. [Google Scholar] [CrossRef]
- Kumar, V.A.; Mohan, T.K.; Murugan, K. Purification and kinetic characterization of polyphenol oxidase from Barbados cherry (Malpighia glabra L.). Food Chem. 2008, 110, 328–333. [Google Scholar] [CrossRef]
- Zhang, Z.; Pang, X.; Duan, X.; Ji, Z.; Jiang, Y. Role of peroxidase in anthocyanin degradation in litchi fruit pericarp. Food Chem. 2005, 90, 47–52. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, H.; Jiang, H.; Xu, Y.; Cao, J.; Jiang, W. Multiple 1-MCP treatment more effectively alleviated postharvest nectarine chilling injury than conventional one-time 1-MCP treatment by regulating ROS and energy metabolism. Food Chem. 2020, 330, 127256. [Google Scholar] [CrossRef]
- Su, Z.; Hu, M.; Gao, Z.; Li, M.; Jiang, Y. Apple polyphenols delay senescence and maintain edible quality in litchi fruit during storage. Postharvest Biol. Technol. 2019, 157, 110976. [Google Scholar] [CrossRef]
- Smith, I.K.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5,5′-dithiobis (2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef]
- Liu, J.; Li, F.; Liang, L.; Jiang, Y.; Chen, J. Fibroin Delays Chilling Injury of Postharvest Banana Fruit via Enhanced Antioxidant Capability during Cold Storage. Metabolites 2019, 9, 152. [Google Scholar] [CrossRef] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Library | Raw Reads (bp) | Raw Bases(G) | Clean Reads (bp) | Clean Bases (G) | Er% | Q20% | Q30% | GC% |
|---|---|---|---|---|---|---|---|---|---|
| CK0d-1 | FRAS220060494-1r | 47,917,660 | 7.19 G | 46,984,726 | 7.05 G | 0.03 | 96.74 | 91.51 | 49.72 |
| CK0d-2 | FRAS220052691-2r | 41,150,130 | 6.17 G | 40,322,240 | 6.05 G | 0.03 | 96.54 | 91.25 | 49.05 |
| CK0d-3 | FRAS220052692-2r | 42,604,436 | 6.39 G | 41,244,352 | 6.19 G | 0.03 | 96.87 | 91.82 | 48.73 |
| TR0d-1 | FRAS220052693-2r | 45,285,588 | 6.79 G | 44,454,528 | 6.67 G | 0.03 | 96.74 | 91.55 | 48.64 |
| TR0d-2 | FRAS220052694-2r | 45,354,954 | 6.8 G | 44,426,710 | 6.66 G | 0.03 | 97.03 | 92.09 | 48.68 |
| TR0d-3 | FRAS220052695-2r | 46,247,152 | 6.94 G | 45,288,912 | 6.79 G | 0.03 | 96.95 | 91.96 | 47.58 |
| CK3d-1 | FRAS220052696-2r | 45,892,678 | 6.88 G | 45,465,842 | 6.82 G | 0.03 | 96.99 | 92.16 | 50.89 |
| CK3d-2 | FRAS220052697-2r | 46,614,824 | 6.99 G | 45,989,324 | 6.9 G | 0.03 | 96.8 | 91.62 | 49.54 |
| CK3d-3 | FRAS220052698-2r | 45,824,306 | 6.87 G | 44,867,480 | 6.73 G | 0.03 | 96.77 | 91.59 | 49.78 |
| TR3d-1 | FRAS220052699-2r | 55,346,256 | 8.3 G | 52,010,944 | 7.8 G | 0.03 | 96.85 | 91.85 | 50.09 |
| TR3d-2 | FRAS220052700-2r | 41,487,432 | 6.22 G | 37,793,338 | 5.67 G | 0.03 | 97.09 | 92.33 | 49.92 |
| TR3d-3 | FRAS220060495-1r | 45,300,832 | 6.8 G | 44,175,794 | 6.63 G | 0.03 | 96.4 | 90.89 | 51.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Zhang, Q.; Tian, C.; Liu, G.; Pan, Y.; Xu, X.; Shi, X.; Zhang, Z.; Meng, L. Physiological and Transcriptome Analyses of CaCl2 Treatment to Alleviate Chilling Injury in Pineapple. Plants 2022, 11, 2215. https://doi.org/10.3390/plants11172215
Zhang M, Zhang Q, Tian C, Liu G, Pan Y, Xu X, Shi X, Zhang Z, Meng L. Physiological and Transcriptome Analyses of CaCl2 Treatment to Alleviate Chilling Injury in Pineapple. Plants. 2022; 11(17):2215. https://doi.org/10.3390/plants11172215
Chicago/Turabian StyleZhang, Mengzhuo, Qiang Zhang, Cong Tian, Guangsen Liu, Yonggui Pan, Xiangbin Xu, Xuequn Shi, Zhengke Zhang, and Lanhuan Meng. 2022. "Physiological and Transcriptome Analyses of CaCl2 Treatment to Alleviate Chilling Injury in Pineapple" Plants 11, no. 17: 2215. https://doi.org/10.3390/plants11172215