
Inoculation with Arbuscular Mycorrhizal Fungi Alleviates the Adverse Effects of High Temperature in Soybean

 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect on Growth Parameters and Root Colonization
2.2. Effect on SPAD Value, Canopy Temperature and Stomatal Density
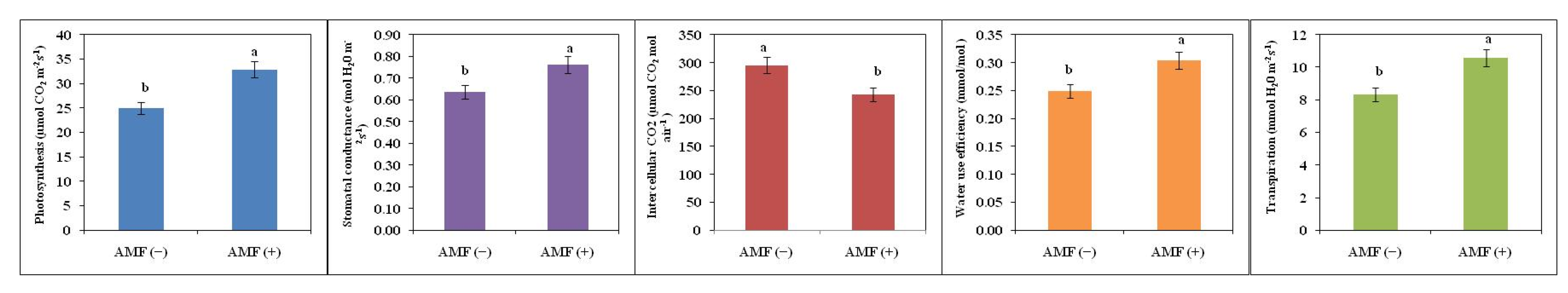
2.3. Effect on Gas Exchange Parameters
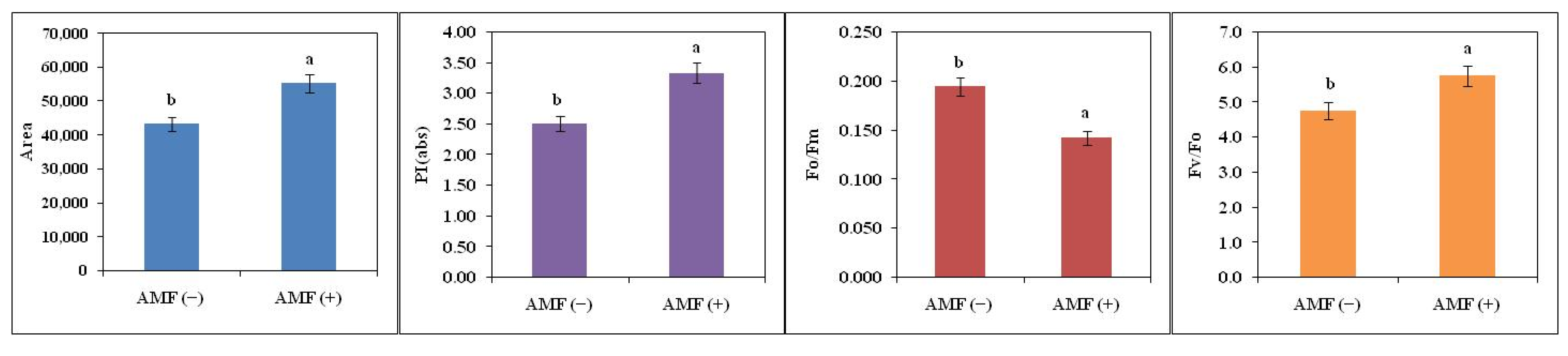
2.4. Effect on Chlorophyll a Fluorescence
2.5. Effect on Energy Pipeline Leaf Model
2.6. Effect on Seed Yield and Its Attributes
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. AM Fungi Inoculum
4.3. Growth Parameters and Assessment of AMF Root Colonization
4.4. Canopy Temperature
4.5. Chlorophyll Content
4.6. Stomatal Density
4.7. GasExchange Measurements
4.8. Chlorophyll a Fluorescence
4.9. Energy Pipeline Leaf Model
4.10. Seed Yield and Its Attributes
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ashraf, M.; Hafeez, M. Thermotolerance of pearl millet and maize at early growth stages: Growth and nutrient relations. Biol. Plant. 2004, 48, 81–86. [Google Scholar]
- Wahid, A.; Close, T.J. Expression of dehydrins under heat stress and their relationship with water relations of sugarcane leaves. Biol. Plant. 2007, 51, 104–109. [Google Scholar]
- Meehl, G.A.; Stocker, T.F.; Collins, W.D.; Gaye, A.J.; Gregory, J.M.; Kitoh, A.; Knutti, R.; Murphy, J.M.; Noda, A.; Raper, S.C.B.; et al. (Eds.) Global Climate Projections; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; pp. 749–844. [Google Scholar]
- Thomas, J.M.G.; Boote, K.J.; Allen, L.H., Jr.; Gallo Meagher, M.; Davis, J.M. Elevated temperature and carbon dioxide effects on soybean seed germination and transcript abundance. Crop Sci. 2003, 43, 1548–1557. [Google Scholar]
- Jumrani, K.; Bhatia, V.S. Impact of combined stress of high temperature and water deficit on growth and seed yield of soybean. Physiol. Mol. Biol. Plants 2018, 24, 37–50. [Google Scholar]
- Jumrani, K.; Bhatia, V.S. Identification of drought tolerant genotypes using physiological traits in soybean. Physiol. Mol. Biol. Plants 2019, 25, 667–681. [Google Scholar]
- Jumrani, K.; Bhatia, V.S. Combined effect of temperature and water stress on physiological and biochemical processes in soybean (Glycine max). Physiol. Mol. Biol. Plants 2019, 25, 697–711. [Google Scholar]
- Liu, X.B.; Jin, J.; Wang, G.H.; Herbert, S.J. Soybean yield physiology and development of high yielding practices in North east China. Field Crops Res. 2008, 105, 157–171. [Google Scholar]
- Tacarindua, C.R.P.; Shiraiwa, T.; Homma, K.; Kumagai, E.; Sameshima, R. The response of soybean seed growth characteristics to increased temperature under near-field conditions in a temperature gradient chamber. Field Crops Res. 2012, 131, 26–31. [Google Scholar]
- Jumrani, K.; Bhatia, V.S. Influence of different light intensities on specific leaf weight, stomatal density, photosynthesis and seed yield in soybean. Indian J. Plant Physiol. 2020, 25, 277–283. [Google Scholar]
- Jumrani, K.; Bhatia, V.S.; Pandey, G.P. Impact of elevated temperatures on specific leaf weight, stomatal density, photosynthesis and chlorophyll fluorescence in soybean. Photosynth Res. 2017, 131, 333–350. [Google Scholar]
- Gavito, M.E.; Rouhier, H.; Olsson, P.A.; Medina, P.A.; Jakobsen, I.; Bago, B.; Azcon, A.C. Temperature constraints on the growth and functioning of root organ cultures with arbuscular mycorrhizal fungi. N. Phytol. 2005, 168, 179–188. [Google Scholar]
- Meharg, A.A. Variation in arsenic accumulation—Hyper accumulation in ferns and their allies. N. Phytol. 2003, 157, 25–31. [Google Scholar]
- Baumann, K.; Schneider, B.U.; Marschner, P.; Huttl, R.F. Root distribution and nutrient status of mycorrhizal and non mycorrhizal Pinussylvestris, L. seedlings growing in a sandy substrate with lignite fragments. Plant Soil 2005, 276, 347–357. [Google Scholar]
- Smith, S.E.; Read, D.J. Arbuscular mycorrhizas, Mycorrhizal Symbiosis; Academic Press: San Diego, CA, USA, 2008; pp. 1–188. [Google Scholar]
- Andrade, G. Role of Functional Groups of Microorganisms on the Rhizosphere Microcosm Dynamics in Plant Surface Microbiology; Varma, A., Abbott, L., Werne, D., Hampp, R., Eds.; Springer: New York, NY, USA, 2004; pp. 51–68. [Google Scholar]
- Selosse, M.A.; Richard, F.; He, X.H.; Simard, S.W. Mycorrhizal networks: Desliaisons dangereuses? Trends Ecol. Evol. 2006, 21, 621–628. [Google Scholar]
- Gadkar, V.; David-Schwartz, R.; Kunik, T.; Kapilnik, Y. Arbuscular mycorrhizal fungal colonization: Factors involved in host recognition. Plant Physiol. 2001, 127, 1493–1499. [Google Scholar]
- Harrison, M.J. Signaling in the arbuscular mycorrhizal symbiosis. Annu. Rev. Microbiol. 2005, 59, 19–42. [Google Scholar]
- Auge, R.M.; Toler, H.D.; Moore, J.L.; Cho, K.; Saxton, A.M. Comparing contributions of soil versus root colonization to variations instomatal behavior and soil drying in mycorrhizal Sorghum bicolor and Cucurbita pepo. J. Plant Physiol. 2007, 164, 1289–1299. [Google Scholar]
- Maboko, M.M. Effect of arbuscular mycorrhiza and temperature control on plant growth, yield, and mineral content of tomato plants grown hydroponically. Hortic. Sci. 2013, 48, 1470–1477. [Google Scholar]
- Wezowicz, K.; Rozpądek, P.; Turnau, K. Interactions of arbuscular mycorrhizal and endophytic fungi improve seedling survival and grow thin post-mining waste. Mycorrhiza 2017, 27, 499–511. [Google Scholar]
- Barzana, G.; Aroca, R.; Bienert, G.P.; Chaumont, F.; Ruiz-Lozano, J.M. New insights into the regulation of aquaporins by the arbuscular mycorrhizal symbiosis in maize plants under drought stress and possible implications for plant performance. Mol. Plant Microbe Interact. 2014, 27, 349–363. [Google Scholar]
- Ruíz-Sánchez, M.; Aroca, R.; Munoz, Y.; Polon, R.; Ruíz-Lozano, J.M. The arbuscular mycorrhizal symbiosis enhances the photosynthetic efficiency and the antioxidative response of rice plants subjected to drought stress. J. Plant Physiol. 2010, 167, 862–869. [Google Scholar]
- Zhu, X.C.; Song, F.B.; Xu, H.W. Arbuscular mycorrhizae improves low temperature stress in maize via alterations in host water status and photosynthesis. Plant Soil 2010, 331, 129–137. [Google Scholar]
- Zhu, X.C.; Song, F.B.; Liu, S.Q.; Liu, T.D.; Zhou, X. Arbuscular mycorrhizae improves photosynthesis and water status of Zeamays, L. under drought stress. Plant Soil Environ. 2012, 58, 186–191. [Google Scholar]
- Habibzadeh, Y.; Pirzad, A.; Zardashti, M.R.; Jalilian, J.; Eini, O. Effects of arbuscular mycorrhizal fungi on seed and protein yield under water deficit stress in mungbean. Agron. J. 2013, 105, 79–84. [Google Scholar]
- Porcel, R.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal influence on leaf water potential, solute accumulation, and oxidative stress in soybean plants subjected to drought stress. J. Exp. Bot. 2004, 55, 1743–1750. [Google Scholar]
- Zhu, X.C.; Song, F.B.; Xu, H.W. Influence of arbuscular mycorrhiza on lipid peroxidation and antioxidant enzyme activity of maize plants under temperature stress. Mycorrhiza 2009, 20, 325–332. [Google Scholar]
- Wu, Q.S.; Zou, Y.N.; Liu, W.; Ye, X.F.; Zai, H.F.; Zhao, L.J. Alleviation of salt stress in citrus seedlings inoculated with mycorrhiza: Changes in leaf antioxidant defense systems. Plant Soil Environ. 2010, 56, 470–475. [Google Scholar]
- Lu, X.; Koide, R.T. The effects of mycorrhizal infection on components of plant growth and reproduction. N. Phytol. 1994, 128, 211–218. [Google Scholar]
- Reddy, K.R.; Hodges, H.F. Crop Ecosystem Responses to Global Climate Change: Cotton. In Climate Change and Global Productivity; Reddy, K.R., Hodges, H.F., Eds.; CAB International: Wallingford, UK, 2000; pp. 161–187. [Google Scholar]
- Al-Karaki, G.N. Growth of mycorrhizal tomato and mineral acquisition under salt stress. Mycorrhiza 2000, 10, 51–54. [Google Scholar]
- Auge, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhizac 2001, 11, 3–42. [Google Scholar]
- Hussain, H.A.; Qingwen, Z.; Hussain, S.; Hongbo, L.; Waqqas, A.; Li, Z. Effects of arbuscular mycorrhizal fungi on maize growth, root colonization, and root exudates varied with inoculums and application method. J. Soil Sci. Plant Nutr. 2021, 21, 2. [Google Scholar]
- Ruotsalainen, A.L.; Kytoiita, M.M. Mycorrhizal does not alter low temperature impact on Gnaphaliumnorvegicum. Oecologia 2004, 140, 226–233. [Google Scholar]
- Al Karaki, G.N. Nursery inoculation of tomato with arbuscular mycorrhizal fungi and subsequent performance under irrigation with saline water. Sci. Hortic. 2006, 109, 1–7. [Google Scholar]
- Kaya, C.; Ashraf, M.; Sonmez, O.; Aydemir, S.; Tuna, A.; Cullu, M.A. The influence of arbuscular mycorrhizal colonization on keygrowth parameters and fruit yield of pepper plants grown at high salinity. Sci. Hortic. 2009, 122, 1–6. [Google Scholar]
- Wu, Q.S.; Zou, Y.N. Beneficial roles of arbuscular mycorrhizas in citrus seedlings at temperature stress. Sci. Hortic. 2010, 125, 289–293. [Google Scholar]
- Wu, S.; Zhang, X.; Chen, B.; Wu, Z.; Li, T.; Hu, Y.; Sun, Y.; Wang, Y. Chromium immobilization by extra radical mycelium of arbuscularmycorrhiza contributes to plant chromium tolerance. Environ. Exp. Bot. 2016, 122, 10–18. [Google Scholar]
- Beltrano, J.; Ronco, M. Improved tolerance of wheat plants (Triticum aestivum L.) to drought stress and rewatering by the arbuscular mycorrhizal fungus Glomusclaroideum: Effect on growth and cell membrane stability. Braz. J. Plant Physiol. 2008, 20, 29–37. [Google Scholar]
- Birhane, E.; Sterck, F.J.; Fetene, M.; Bongers, F.; Kuyper, T.W. Arbuscular mycorrhizal fungi enhance photosynthesis, water use efficiency, and growth of frankincense seedlings under pulsed water availability conditions. Oecologia 2012, 169, 895–904. [Google Scholar]
- Maya, M.A.; Matsubara, Y.I. Tolerance to Fusarium wilt and anthracnose diseases and changes of antioxidative activity in mycorrhizal cyclamen. Crop Prot. 2013, 47, 41–48. [Google Scholar]
- Alotaibi, M.O.; Saleh, A.M.; Sobrinho, R.L.; Sheteiwy, M.S.; ElSawah, A.M.; Mohammed, A.E.; Elgawad, H.A. Arbuscular mycorrhizae mitigate aluminium toxicity and regulate proline metabolism in plants grown in acidic soil. J. Fungi 2021, 7, 531. [Google Scholar]
- Lin, A.J.; Zhang, X.H.; Wong, M.H.; Ye, Z.H.; Lou, L.Q.; Wang, Y.S. Increase of multimetal tolerance of three leguminous plants by arbuscular mycorrhizal fungi colonization. Environ. Geochem. Health 2007, 29, 473–481. [Google Scholar]
- De Novais, C.B.; Sbrana, C.; da Conceição, J.E.; Rouws, L.F.M.; Giovannetti, M.; Avio, L.; Siqueira, J.O.; Saggin, O.J., Jr.; Ribeiroda Silva, E.M.; de Faria, S.M. Mycorrhizal networks facilitate the colonization of legume roots by a symbiotic nitrogen-fixing bacterium. Mycorrhiza 2020, 30, 389–396. [Google Scholar]
- Scheublin, T.R.; Ridgway, K.P.; Young, J.P.W.; van der Heijden, M.G.A. Nonlegumes, legumes, and root nodules harbor different arbuscular mycorrhizal fungal communities. Appl. Environ. Microbiol. 2004, 70, 6240–6246. [Google Scholar]
- Ashraf, M. Some important physiological selection criteria for salt tolerance in plants. Flora 2004, 199, 361–376. [Google Scholar]
- Piao, S.; Ciais, P.; Friedlingstein, P.; Peylin, P.; Reichstein, M.; Luyssaert, S.; Margolis, H.; Fang, J.; Barr, A.; Chen, A.; et al. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature 2008, 451, 49–52. [Google Scholar]
- Egli, D.B. Soybean reproductive sink size and short-term reductions in photosynthesis during flowering and pod set. Crop Sci. 2010, 50, 1971–1977. [Google Scholar]
- Saibo, N.J.M.; Lourenço, T.; Oliveira, M.M. Transcription factors and regulation of photosynthetic and related metabolism under environmental stresses. Ann. Bot. 2009, 103, 609–623. [Google Scholar]
- Djanaguiraman, M.; Prasad, P.V.V. Ethylene production under high temperature stress causes premature leaf senescence in soybean. Funct. Plant Biol. 2010, 37, 1071–1084. [Google Scholar]
- Jagadish, S.V.K.; Way, D.A.; Sharkey, T.D. Plant heat stress: Concepts directing future research. Plant Cell Environ. 2021, 44, 1992–2005. [Google Scholar]
- Maggio, A.; Pascale, S.D.; Angelino, G.; Ruggiero, C.; Barbieri, G. Physiological response of tomato to saline irrigation in long terms alinized soils. Eur. J. Agron. 2004, 21, 149–159. [Google Scholar]
- Choe, K.; Toler, H.; Lee, J.; Ownley, B.; Stutz, J.; Moore, J.; Auge, R. Mycorrhizal symbiosis and responses of sorghum plants to combined drought and salinity stresses. J. Plant Physiol. 2006, 163, 517–528. [Google Scholar]
- Auge, R.M.; Toler, H.D.; Sams, C.E.; Nasim, G. Hydraulic conductance and water potential gradients in squash leaves showing mycorrhiza-induced increases in stomatal conductance. Mycorrhiza 2008, 18, 115–121. [Google Scholar]
- Sheng, M.; Tang, M.; Chan, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of arbuscular mycorrhizae on photosynthesis and water status of maize plants under salt stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar]
- Oukarroum, A.; Madidi, S.E.; Schansker, G.; Strasser, R.J. Probing the responses of barley cultivars (Hordeum vulgare L.) by chlorophyll a fluorescence OLKJIP under drought stress and rewatering. Environ. Exp. Bot. 2007, 60, 438–446. [Google Scholar]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypinski, P.; Samborska, I.A.; et al. Prompt chlorophyll fluorescence as a tool for crop phenotyping: An example of barley land races exposed to various abiotic stress factors. Photosynthetica 2018, 56, 953–961. [Google Scholar]
- Rastogi, A.; Stróżecki, M.; Kalaji, K.; Łuców, D.; Lamentowicz, M.; Juszczak, R. Impact of Warming and reduced precipitation on photosynthetic and remote sensing properties of peatland vegetation. Environ. Exp. Bot. 2019, 160, 71–80. [Google Scholar]
- Rastogi, A.; Zivcak, M.; Tripathi, D.K.; Yadav, S.; Kalaji, H.M.; Brestic, M. Phytotoxic effect of silver nanoparticles in Triticum aestivum: Improper regulation of photosystem I activity as the reason for oxidative damage in the chloroplast. Photosynthetica 2019, 57, 209–216. [Google Scholar]
- Rastogi, A.; Kovar, M.; He, X.; Zivcak, M.; Kataria, S.; Kalaji, H.M.; Skalicky, M.; Ibrahimova, U.F.; Hussain, S.; Mbarki, S.; et al. JIP-test asatool to identify salinity tolerance in sweet sorghum genotypes. Photosynthetica 2020, 58, 518–528. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar]
- Ruiz-Lozano, J.M.; Aroca, R.; Zamarreño, A.M.; Molina, S.; Andreo-Jiménez, B.; Porcel, R.; García-Mina, J.M.; Ruyter-Spira, C.; López-Ráez, J.A. Arbuscular mycorrhizal symbiosis induces strigolactone biosynthesis under drought and improves drought tolerance in lettuce and tomato. Plant Cell Environ. 2016, 39, 441–452. [Google Scholar]
- Meinzer, F.C. Functional convergence in plant responses to the environment. Oecologia 2003, 134, 1–11. [Google Scholar]
- Barbieri, G.; Vallone, S.; Orsini, F.; Paradiso, R.; De Pascale, S.; Negre-Zakharov, F.; Maggio, A. Stomatal density and metabolic determinants mediate salt stress adaptation and water use efficiency in basil (Ocimum basilicum L.). J. Plant Physiol. 2012, 169, 1737–1746. [Google Scholar]
- Drake, P.L.; Froend, R.H.; Franks, P.J. Smaller, faster stomata: Scaling of stomatal size, rate of response, and stomatal conductance. J. Exp. Bot. 2013, 64, 495–505. [Google Scholar]
- Abdel Latef, A.A.H.; Chaoxing, H. Does the inoculation with Glomus mosseae improve salt tolerance in pepper plants? J. Plant Growth Regul. 2014, 33, 644–653. [Google Scholar]
- Chitarra, W.; Pagliarani, C.; Maserti, B.; Lumini, E.; Siciliano, I.; Cascone, P.; Schubert, A.; Gambino, G.; Balestrini, R.; Guerrieri, E. Insights on the impact of arbuscular mycorrhizal symbiosis on tomato tolerance to water stress. Plant Physiol. 2016, 171, 1009–1023. [Google Scholar]
- Biermann, B.J.; Lindermann, R.Q. Quantifying vesicular-arbuscular mycorrhizae: Proposed method towards standardization. N. Phytol. 1981, 87, 63–67. [Google Scholar]
- Guendouz, A.; Guessoum, S.; Maamari, K.; Benidir, M.; Hafsi, M. Canopy temperature efficiency as indicators for drought tolerance in Durum Wheat (Triticum Durum Desf.) in semiarid conditions. J Agric Sustain. 2012, 1, 23–38. [Google Scholar]
- Markwell, J.; Osterman, J.; Mitchell, J. Calibration of the Minolta SPAD-502 leaf chlorophyll meter. Photosynth Res. 1995, 46, 467–472. [Google Scholar]
- Radoglou, K.M.; Jarvis, P.G. Effects of CO2 enrichment on four poplar clones. II. Leaf surface properties. Ann. Bot 1990, 65, 627–632. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jumrani, K.; Bhatia, V.S.; Kataria, S.; Alamri, S.A.; Siddiqui, M.H.; Rastogi, A. Inoculation with Arbuscular Mycorrhizal Fungi Alleviates the Adverse Effects of High Temperature in Soybean. Plants 2022, 11, 2210. https://doi.org/10.3390/plants11172210
Jumrani K, Bhatia VS, Kataria S, Alamri SA, Siddiqui MH, Rastogi A. Inoculation with Arbuscular Mycorrhizal Fungi Alleviates the Adverse Effects of High Temperature in Soybean. Plants. 2022; 11(17):2210. https://doi.org/10.3390/plants11172210
Chicago/Turabian StyleJumrani, Kanchan, Virender Singh Bhatia, Sunita Kataria, Saud A. Alamri, Manzer H. Siddiqui, and Anshu Rastogi. 2022. "Inoculation with Arbuscular Mycorrhizal Fungi Alleviates the Adverse Effects of High Temperature in Soybean" Plants 11, no. 17: 2210. https://doi.org/10.3390/plants11172210