
Tree Species Composition and Forest Community Types along Environmental Gradients in Htamanthi Wildlife Sanctuary, Myanmar: Implications for Action Prioritization in Conservation


Abstract
:1. Introduction
2. Results
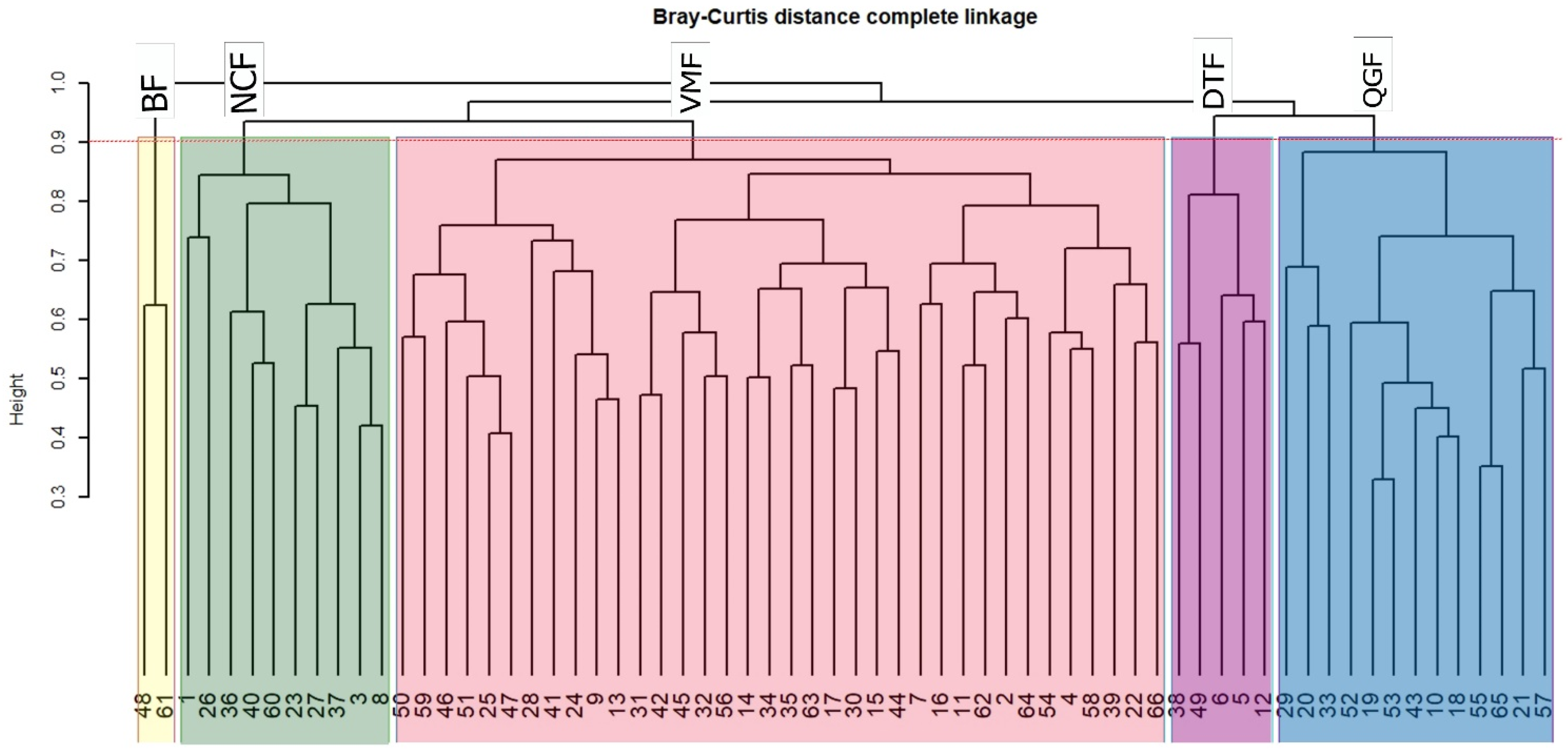
2.1. Forest Community Assemblages
2.2. Species Diversity and Importance Values among Forest Communities
2.3. Variations in Topography and Soil Characteristics among Forest Communities
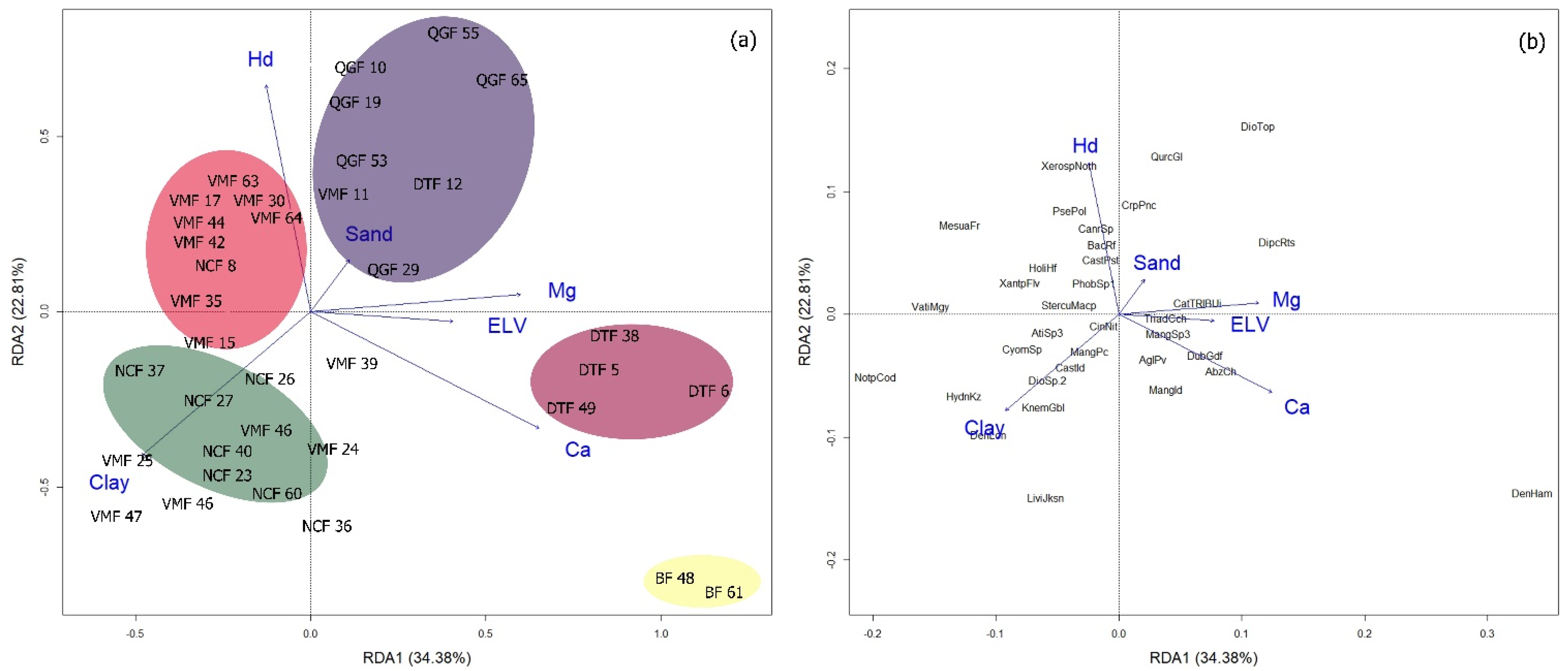
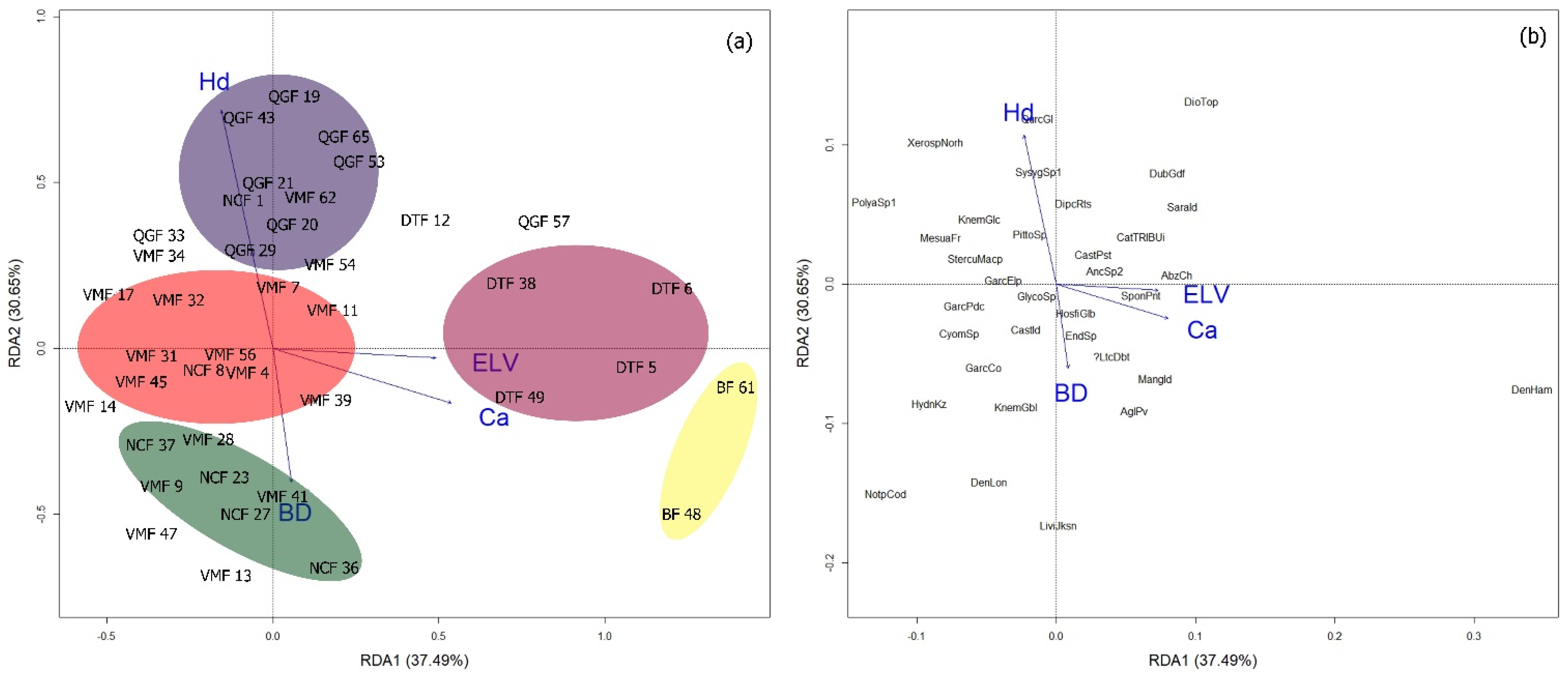
2.4. Redundancy Analysis (RDA) Biplot and Important Topographic and Edaphic Variables
3. Discussion
4. Materials and Methods
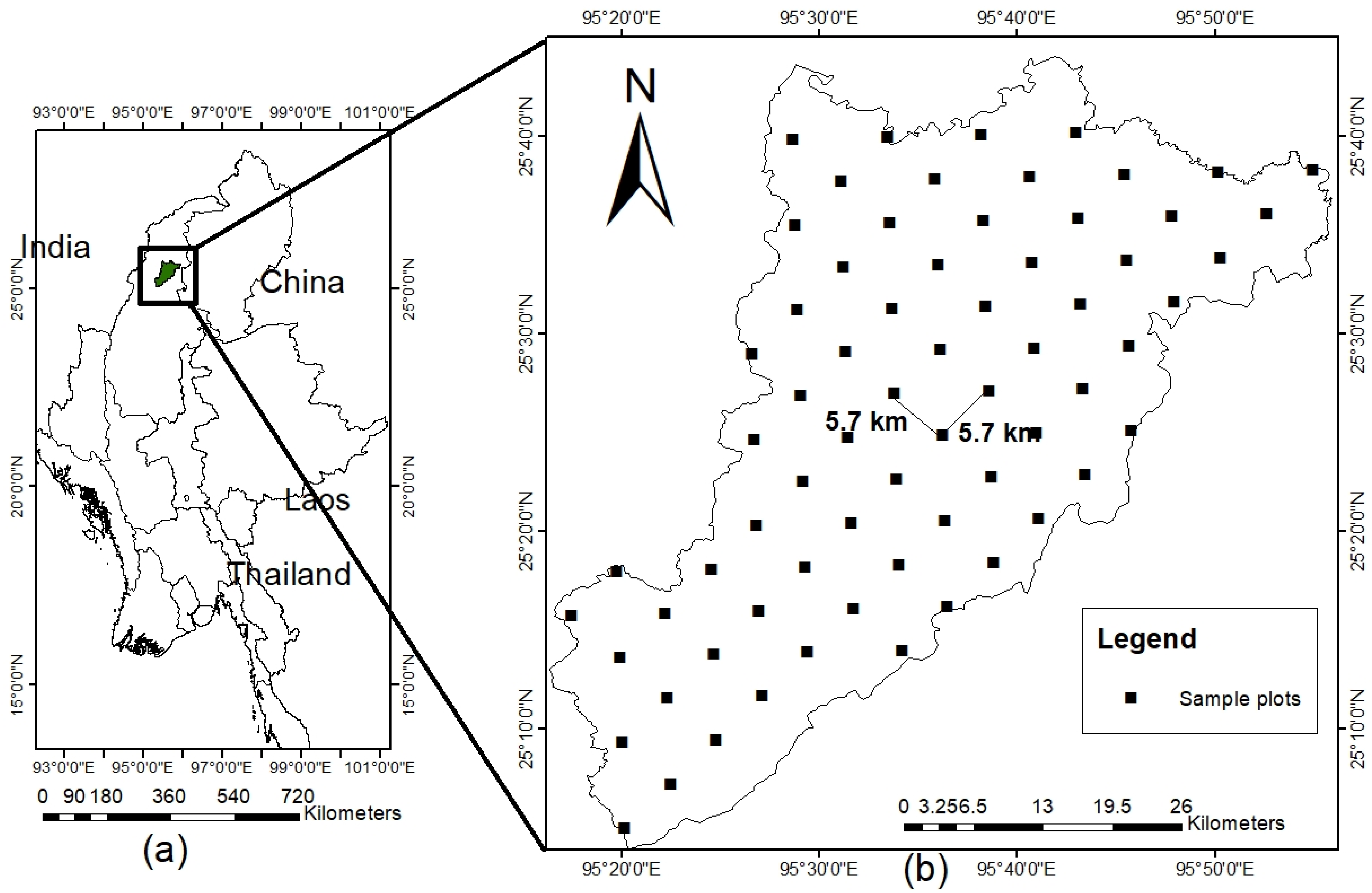
4.1. Study Site and Sampling Method
4.2. Determination of Topographic and Edaphic Variables
4.3. Data and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Saunders, S.C.; Crow, T.R.; Naiman, R.J.; Brosofske, K.D.; Mroz, G.D.; Brookshire, B.L.; Franklin, J.F. Microclimate in forest ecosystem and landscape ecology. BioScience 1999, 49, 288–297. [Google Scholar] [CrossRef] [Green Version]
- Duchicela, S.A.; Cuesta, F.; Tovar, C.; Muriel, P.; Jaramillo, R.; Salazar, E.; Pinto, E. Microclimatic warming leads to a decrease in species and growth form diversity: Insights from a tropical alpine grassland. Front. Ecol. Evol. 2021, 9, 673655. [Google Scholar] [CrossRef]
- Rahman, I.U.; Afzal, A.; Iqbal, Z.; Bussmann, R.W.; Alsamadany, H.; Calixto, E.S.; Shah, G.M.; Kausar, R.; Shah, M.; Ali, N.; et al. Ecological gradients hosting plant communities in Himalayan subalpine pastures: Application of multivariate approaches to identify indicator species. Ecol. Inform. 2020, 60, 101162. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.-F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Jianmeng, F. Biogeographical interpretation of elevational patterns of genus diversity of seed plants in Nepal. PLoS ONE 2015, 10, e0140992. [Google Scholar] [CrossRef]
- Nepali, B.R.; Skartveit, J.; Baniya, C.B. Impacts of slope aspects on altitudinal species richness and species composition of Narapani-Masina Landscape, Arghakhanchi, West Nepal. J. Asia-Pac. Biodivers. 2021, 14, 415–424. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Grubb, P.J. Regional and local patterns in plant species richness with respect to resource availability. Oikos 2003, 100, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Toro Manríquez, M.D.R.; Cellini, J.M.; Lencinas, M.V.; Peri, P.L.; Peña Rojas, K.A.; Martínez Pastur, G.J. Suitable conditions for natural regeneration in variable retention harvesting of southern Patagonian nothofagus pumilio forests. Ecol. Process. 2019, 8, 1–12. [Google Scholar] [CrossRef]
- Davis, E.L.; Hager, H.A.; Gedalof, Z. Soil properties as constraints to seedling regeneration beyond alpine treelines in the Canadian Rocky Mountains. Arct. Antarct. Alp. Res. 2018, 50, e1415625-1-15. [Google Scholar] [CrossRef] [Green Version]
- Wenk, E.H.; Dawson, T.E. Interspecific differences in seed germination, establishment, and early growth in relation to preferred soil type in an Alpine community. Arct. Antarct. Alp. Res. 2007, 39, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Palpurina, S.; Wagner, V.; Wehrden, H.V.; Hájek, M.; Horsák, M.; Brinkert, A.; Hölzel, N.; Wesche, K.; Kamp, J.; Hájková, P.; et al. The relationship between plant species richness and soil pH vanishes with increasing aridity across Eurasian dry grasslands. Glob. Ecol. Biogeogr. 2016, 26, 425–434. [Google Scholar] [CrossRef]
- Tyler, G. Some ecophysiological and historical approaches to species richness and calcicole/calcifuge behaviour—Contribution to a debate. Folia Geobot. 2003, 38, 419–428. [Google Scholar] [CrossRef]
- Currie, D.J.; Mittelbach, G.G.; Cornell, H.V.; Field, R.; Guegan, J.F.; Hawkins, B.A.; Kaufman, D.M.; Kerr, J.T.; Oberdorff, T.; O’Brien, E.; et al. Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 2004, 7, 1121–1134. [Google Scholar] [CrossRef]
- Naing, H.; Ross, J.; Burnham, D.; Htun, S.; Macdonald, D.W. Population density estimates and conservation concern for clouded leopards neofelis nebulosa, marbled cats pardofelis marmorata and tigers panthera tigris in htamanthi wildlife sanctuary, Sagaing, Myanmar. Oryx 2017, 53, 654–662. [Google Scholar] [CrossRef] [Green Version]
- Seddon, N.; Chausson, A.; Berry, P.; Girardin, C.A.; Smith, A.; Turner, B. Understanding the value and limits of nature-based solutions to climate change and other global challenges. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rodríguez, E.J.; Beltrán, J.F.; El Mouden, E.H.; Slimani, T.; Márquez, R.; Donaire-Barroso, D. Climate change challenges IUCN conservation priorities: A test with Western Mediterranean amphibians. SN Appl. Sci. 2020, 2, 216. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, J.O.; Maldia, L.S.J.; Park, B.B. Research trends and methodological approaches of the impacts of windstorms on forests in tropical, subtropical, and temperate zones: Where are we now and how should research move forward? Plants 2020, 9, 1709. [Google Scholar] [CrossRef]
- Dibaba, A.; Soromessa, T.; Warkineh, B. Plant community analysis along environmental gradients in moist afromontane forest of Gerba Dima, south-western Ethiopia. BMC Ecol. Evol. 2021, 22, 1–17. [Google Scholar] [CrossRef]
- Oliver, T.H.; Heard, M.S.; Isaac, N.J.B.; Roy, D.B.; Procter, D.; Eigenbrod, F.; Freckleton, R.; Hector, A.; Orme, C.D.; Petchey, O.L.; et al. Biodiversity and resilience of ecosystem functions. Trends Ecol. Evol. 2015, 30, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Davies, S.J.; Tan, S.; LaFrankie, J.V.; Potts, M.D. Soil-related floristic variation in the hyperdiverse dipterocarp forest in Lambir Hills, Sarawak. In Pollination Ecology and Rain Forest Diversity, Sarawak Studies; Roubik, D.W., Sakai, S., Hamid, A., Eds.; Springer: New York, NY, USA, 2005; pp. 22–34. [Google Scholar]
- Huszár, T.; Mika, J.; Lóczy, D.; Molnár, K.; Kertész, Á. Climate change and soil moisture: A case study. Phys. Chem. Earth Part A Solid Earth Geod. 1999, 24, 905–912. [Google Scholar] [CrossRef]
- Deepthy, R.; Balakrishnan, S. Climatic control on clay mineral formation: Evidence from weathering profiles developed on either side of the Western Ghats. J. Earth Syst. Sci. 2005, 114, 545–556. [Google Scholar] [CrossRef]
- Birkás, M. Tillage, impacts on soil and environment. In Encyclopedia of Agrophysics. Encyclopedia of Earth Sciences Series; Gliński, J., Horabik, J., Lipiec, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 903–906. [Google Scholar] [CrossRef]
- Ampoorter, E.; de Frenne, P.; Hermy, M.; Verheyen, K. Effects of soil compaction on growth and survival of tree saplings: A meta-analysis. Basic Appl. Ecol. 2011, 12, 394–402. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; He, D.; Shen, F.; Huang, J.; Zhang, R.; Liu, W.; Zhu, M.; Zhou, L.; Wang, L.; Zhou, Q. Effects of soil compaction on plant growth, nutrient absorption, and root respiration in soybean seedlings. Environ. Sci. Pollut. Res. 2019, 26, 22835–22845. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, J.O.; An, J.Y.; Combalicer, M.S.; Chun, J.P.; Oh, S.K.; Park, B.B. Morpho-anatomical traits and soluble sugar concentration largely explain the responses of three deciduous tree species to progressive water stress. Front. Plant Sci. 2021, 12, 738301. [Google Scholar] [CrossRef]
- Walter, J.; Hein, R.; Auge, H.; Beierkuhnlein, C.; Löffler, S.; Reifenrath, K.; Schädler, M.; Weber, M.; Jentsch, J. How do extreme drought and plant community composition affect host plant metabolites and herbivore performance? Arthropod-Plant Interact. 2012, 6, 15–25. [Google Scholar] [CrossRef]
- Ohdo, T.; Takahashi, K. Plant species richness and community assembly along gradients of elevation and soil nitrogen availability. AoB PLANTS 2020, 12, plaa014. [Google Scholar] [CrossRef] [Green Version]
- Drollinger, S.; Müller, M.; Kobl, T.; Schwab, N.; Böhner, J.; Schickhoff, U.; Scholten, T. Decreasing nutrient concentrations in soils and trees with increasing elevation across a treeline ecotone in Rolwaling Himal, Nepal. J. Mt. Sci. 2017, 14, 843–858. [Google Scholar] [CrossRef]
- Lalfakawma; Sahoo, U.K.; Roy, S.; Vanlalhriatpuia, K.; Vanalalhluna, P.C. Community composition and tree population structure in undisturbed and disturbed tropical semi-evergreen forest stands of north-east India. Appl. Ecol. Environ. Res. 2009, 7, 303–318. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Communities | Richness (S) | Shannon-Wiener Diversity Index (H) | Evenness (J′) | Dominant Species and Importance Values (IV) |
|---|---|---|---|---|
| BF | 10 | 0.086 | 0.037 | Aglaia perviridis (17.3%) |
| DTF | 51 | 2.659 | 0.676 | Diospyros toposia (25.2%) |
| NCF | 95 | 2.807 | 0.616 | Nothaphoebe condensa (26.2%) |
| QGF | 65 | 3.254 | 0.780 | Quercus glauca (30.9%) |
| VMF | 174 | 3.965 | 0.769 | Vatica maingayi (35.9%) |
| FC | n | Elevation (masl) | Slope (°) |
|---|---|---|---|
| DTF | 5 | 274 (57) a | 29 (19) a |
| NCF | 10 | 188 (32) ab | 8 (6) b |
| QGF | 13 | 179 (24) b | 16 (12) ab |
| VMF | 36 | 177 (31) b | 16 (14) ab |
| Soil Depth (cm) | Forest Community | n | Soil Hardness (kg/cm2) | Moisture Content (%) | Bulk Density (%) | Organic Matter (%) | Sand (%) | Silt (%) | Clay (%) |
|---|---|---|---|---|---|---|---|---|---|
| 0–15 | DTF | 5 | 2.50 (0.55) a | 10.30 (3.19) a | 1.18 (0.06) a | 6.00 (1.87) a | 54.2 (4.2) c | 29.0 (4.2) a | 16.2 (3.5) a |
| NCF | 10 | 2.79 (0.88) a | 20.10 (4.32) b | 1.18 (0.14) a | 7.30 (2.11) a | 39.4 (13.1) a | 30.3 (7.6) a | 29.2 (7.3) b | |
| QGF | 13 | 3.31 (0.72) a | 17.89 (8.04) ab | 1.10 (0.09) a | 6.69 (1.25) a | 50.0 (5.3) bc | 29.8 (3.7) a | 19.1 (3.8) a | |
| VMF | 36 | 3.00 (1.11) a | 20.10 (9.80) b | 1.19 (0.14) a | 6.00 (1.87) a | 46.6 (6.7) ab | 27.6 (5.0) a | 24.5 (6.1) b | |
| 15–30 | DTF | 5 | 3.06 (1.18) a | 12.00 (2.95) a | 1.13 (0.09) a | 6.20 (1.48) a | 46.8 (4.3) a | 27.0 (3.2) a | 22.6 (3.0) a |
| NCF | 10 | 2.89 (0.87) a | 21.80 (3.50) b | 1.16 (0.11) a | 7.70 (1.34) a | 42.1 (2.5) a | 28.9 (8.2) a | 32.6 (5.9) b | |
| QGF | 13 | 3.94 (0.48) b | 16.30 (2.52) a | 1.05 (0.10) a | 7.15 (1.14) a | 46.2 (2.2) a | 28.8 (3.7) a | 23.1 (4.3) a | |
| VMF | 36 | 3.05 (1.02) ab | 19.80 (4.63) b | 1.13 (0.09) a | 6.97 (1.13) a | 42.3 (1.3) a | 26.6 (4.8) a | 27.9 (6.8) ab |
| Soil Depth (cm) | Forest Community | n | pH | TN (g/kg) | AP (mg/kg) | K (mg/100 g) | Ca (mg/100 g) | Na (mg/100 g) | Mg (mg/100 g) |
|---|---|---|---|---|---|---|---|---|---|
| 0–15 | DTF | 5 | 5.12 (0.19) a | 0.590 (0.136) a | 80.0 (7.10) a | 8.40 (3.78) a | 63.80 (6.44) a | 0.30 (0.07) a | 154.40 (144.0) a |
| NCF | 10 | 4.94 (0.09) a | 0.657 (0.285) a | 47.0 (35.0) b | 5.20 (3.52) a | 10.20 (6.25) b | 0.32 (0.09) a | 21.60 (28.7) ab | |
| QGF | 13 | 4.85 (0.23) a | 0.611 (0.189) a | 55.4 (13.9) b | 4.15 (1.57) a | 5.15 (3.63) b | 0.22 (0.15) a | 18.77 (18.2) ab | |
| VMF | 36 | 4.89 (0.11) a | 0.663 (0.141) a | 50.0 (20.6) b | 5.14 (3.09) a | 7.36 (8.59) b | 0.28 (0.14) a | 12.19 (18.0) b | |
| 15–30 | DTF | 5 | 4.98 (0.12) a | 0.622 (0.955) a | 80.0 (21.5) a | 8.80 (4.32) a | 13.60 (12.60) a | 0.24 (0.11) a | 72.40 (49.0) a |
| NCF | 10 | 4.97 (0.09) a | 0.572 (0.174) a | 44.0 (32.4) b | 5.00 (3.56) a | 7.60 (3.92) ab | 0.34 (0.18) a | 19.80 (29.8) b | |
| QGF | 13 | 5.08 (0.84) a | 0.600 (0.129) a | 45.4 (32.4) b | 4.08 (1.93) a | 3.15 (2.61) b | 0.19 (0.13) a | 19.46 (21.6) b | |
| VMF | 36 | 4.94 (0.13) a | 0.618 (0.172) a | 42.8 (16.1) b | 5.28 (3.09) a | 6.39 (7.53) ab | 0.26 (0.16) a | 19.19 (54.7) b |
| Parameters | Description | Formula |
|---|---|---|
| Species density | The actual size or number of individuals of one species per unit area. | Den = No. of individuals of each species/AS |
| Relative density | The density of one species as a percent of the total density of all species. | RDen = (Den for a species/total density for all identified species) × 100 |
| Species frequency | The number of times a plant species is present in a given number of plots or quadrats. | Fre = No. of plots in which species occur/ total number of plots sampled |
| Relative frequency | The frequency of one species as a percent of the total frequency of all species. | RFre = (Fre for a species/total frequency for all identified species) × 100 |
| Dominance | A species that is most commonly found or dominant based on basal area or percent coverage. | Dom = BA of one species/AS |
| Relative dominance | The dominance of one species as a percent of the total dominance of all species. | RDom = (Dom for a species/total dominance for all identified species) × 100 |
| Importance values | The IV is a measure of how dominant a species is in a given forest area. | IV = RDen + RFre + RDom |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latt, M.M.; Park, B.B. Tree Species Composition and Forest Community Types along Environmental Gradients in Htamanthi Wildlife Sanctuary, Myanmar: Implications for Action Prioritization in Conservation. Plants 2022, 11, 2180. https://doi.org/10.3390/plants11162180
Latt MM, Park BB. Tree Species Composition and Forest Community Types along Environmental Gradients in Htamanthi Wildlife Sanctuary, Myanmar: Implications for Action Prioritization in Conservation. Plants. 2022; 11(16):2180. https://doi.org/10.3390/plants11162180
Chicago/Turabian StyleLatt, Myo Min, and Byung Bae Park. 2022. "Tree Species Composition and Forest Community Types along Environmental Gradients in Htamanthi Wildlife Sanctuary, Myanmar: Implications for Action Prioritization in Conservation" Plants 11, no. 16: 2180. https://doi.org/10.3390/plants11162180