
Description of an Arabica Coffee Ideotype for Agroforestry Cropping Systems: A Guideline for Breeding More Resilient New Varieties

 ,
,
Abstract
:1. Introduction
2. Agroforestry Is the Best Way to Secure Coffee Production in the Future
3. No Coffee Varieties Have Been Specifically Selected for Agroforestry Systems
4. Lessons Learned from the European BREEDCAFS Project
5. Select F1 Hybrids Rather Than Line Varieties
6. Definition of an Ideotype and Its Use in Breeding
7. Main Arabica Coffee Breeding Goals for AFS
8. Identification of Morpho-Physiological Traits to Achieve a Specific Goal
8.1. Trait 1: Productivity
8.1.1. Vigor and Tree Volume
8.1.2. Floral Induction
8.2. Trait 2: Quality
8.3. Trait 3: Plant Health
8.4. Trait 4: Low Fertilization
9. Coffea Arabica Ideotype for AFS Cropping Systems
10. Possible Ways to Select Targeted Traits within an Ideotype
11. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Steffen, W.; Richardson, K.; Rockström, J.; Cornell, S.E.; Fetzer, I.; Bennett, E.M.; Biggs, R.; Carpenter, S.R.; de Vries, W.; de Wit, C.A.; et al. Planetary boundaries: Guiding human development on a changing planet. Science 2015, 347, 1259855. [Google Scholar] [CrossRef] [PubMed]
- Beroya-Eitner, M.A. Ecological vulnerability indicators. Ecol. Indic. 2016, 60, 329–334. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Pérez Jimenez, J.G.; Montagnon, C.; Schilling, T. Multiclass classification of agro-ecological zones for Arabica coffee: An improved understanding of the impacts of climate change. PLoS ONE 2015, 10, e0140490. [Google Scholar] [CrossRef] [PubMed]
- Djerrab, D.; Bertrand, B.; Breitler, J.-C.; Léran, S.; Dechamp, E.; Campa, C.; Barrachina, C.; Conejero, G.; Etienne, H.; Sulpice, R. Photoperiod-dependent transcriptional modifications in key metabolic pathways in Coffea arabica. Tree Physiol. 2020, 41, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Kreft, S.; Eckstein, D.; Dorsch, L.; Fischer, L. Global Climate Risk Index 2016: Who Suffers Most from Extreme Weather Events? Weather-related Loss Events in 2014 and 1995 to 2014 (Bonn: Germanwatch e.v., 2016). 2016. Available online: http://germanwatch.org/ (accessed on 1 September 2021).
- Bellachew, B.; Sacko, J.C. Coffee genetic resources under severe threat from genetic erosion in the centres of origin and diversity: An urgent need for conservation measures. In Proceedings of the 22nd International Conference on Coffee Science, ASIC 2008, Campinas, SP, Brazil, 14–19 September 2008; Association Scientifique Internationale du Café (ASIC)p: Paris, France, 2009; pp. 1487–1496. [Google Scholar]
- Davis, A.P.; Gole, T.W.; Baena, S.; Moat, J. The Impact of Climate Change on Indigenous Arabica Coffee (Coffea arabica): Predicting Future Trends and Identifying Priorities. PLoS ONE 2012, 7, e47981. [Google Scholar] [CrossRef]
- Baca, M.; Läderach, P.; Haggar, J.; Schroth, G.; Ovalle, O. An Integrated Framework for Assessing Vulnerability to Climate Change and Developing Adaptation Strategies for Coffee Growing Families in Mesoamerica. PLoS ONE 2014, 2, e88463. [Google Scholar] [CrossRef]
- Bunn, C.; Läderach, P.; Ovalle Rivera, O.; Kirschke, D. A bitter cup: Climate change profile of global production of Arabica and Robusta coffee. Clim. Chang. 2015, 129, 89–101. [Google Scholar] [CrossRef]
- Grüter, R.; Trachsel, T.; Laube, P.; Jaisli, I. Expected global suitability of coffee, cashew and avocado due to climate change. PLoS ONE 2022, 17, e0261976. [Google Scholar] [CrossRef]
- Davis, A.P.; Chadburn, H.; Moat, J.; O’Sullivan, R.; Hargreaves, S.; Nic Lughadha, E. High extinction risk for wild coffee species and implications for coffee sector sustainability. Sci. Adv. 2019, 5, eaav3473. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Pais, I.P.; Leitão, A.E.; Guerra, M.; Reboredo, F.H.; Máguas, C.M.; Carvalho, M.L.; Scotti-Campos, P.; Ribeiro-Barros, A.I.; Lidon, F.J.C.; et al. Can Elevated Air [CO2] Conditions Mitigate the Predicted Warming Impact on the Quality of Coffee Bean? Front. Plant Sci. 2018, 9, 287. [Google Scholar] [CrossRef]
- Avila, R.T.; de Almeida, W.L.; Costa, L.C.; Machado, K.L.; Barbosa, M.L.; de Souza, R.P.; Martino, P.B.; Juárez, M.A.; Marçal, D.M.; Martins, S.C.; et al. Elevated air [CO2] improves photosynthetic performance and alters biomass T accumulation and partitioning in drought-stressed coffee plants. Environ. Exp. Bot. 2020, 177, 104137. [Google Scholar] [CrossRef]
- DaMatta, F.; Avila, R.; Cardoso, A.; Martins, S.; Ramalho, J.C. Physiological and Agronomic Performance of the Coffee Crop in the Context of Climate Change and Global Warming: A Review. J. Agric. Food Chem. 2018, 66, 5264–5274. [Google Scholar] [CrossRef] [PubMed]
- Mahon, N.; Crute, I.; Simmons, E.; Islam, M.M. Sustainable intensification—“oxymoron” or “third-way”? A systematic review. Ecol. Indic. 2017, 74, 73–97. [Google Scholar] [CrossRef]
- Struik, P.C.; Kuyper, T.W. Sustainable intensification in agriculture: The richer shade of green. A review. Agron. Sustain. Dev. 2017, 37, 39. [Google Scholar] [CrossRef]
- FAO. Save and Grow. A Policymaker’s Guide to the Sustainable Intensification of Smallholder Crop Production; FAO: Rome, Italy, 2017; Available online: http://www.fao.org/ag/save-and-grow/ (accessed on 1 November 2021).
- Bettles, J.; Battisti, D.; Cook-Patton, S.; Kroeger, T.; Spector, J.; Wolff, N.; Masuda, Y. Agroforestry and non-state actors: A review. For. Policy Econ. 2021, 130, 102538. [Google Scholar] [CrossRef]
- Lammerts van Bueren, E.T.; Struik, P.; van Eekeren, N.; Nuijten, E. Towards resilience through systems-based plant breeding. A review. Agron. Sustain. Dev. 2018, 38, 1–21. [Google Scholar] [CrossRef]
- Villarreyna, R.; Avelino, J.; Cerda, R. Ecosystem-based adaptation: Effect of shade trees on ecosystem services in coffee plantations. Agron. Mesoamer. 2020, 31, 499–516. [Google Scholar] [CrossRef]
- Zaro, G.C.; Caramori, P.H.; Junior, G.M.Y.; Sanquetta, C.R.; Filho, A.A.; Nunes, A.L.P.; Prete, C.E.C.; Voroney, P. Carbon sequestration in an agroforestry system of coffee with rubber trees compared to open-grown coffee in southern Brazil. Agrofor. Syst. 2020, 94, 799–809. [Google Scholar] [CrossRef]
- Jha, S.; Bacon, C.M.; Philpott, S.M.; Ernesto Méndez, V.; Läderach, P.; Rice, R.A. Shade coffee: Update on a disappearing refuge for biodiversity. BioScience 2014, 64, 416–428. [Google Scholar] [CrossRef]
- Lin, B.B. Agroforestry management as an adaptive strategy against potential microclimate extremes in coffee agriculture. Agric. For. Meteorol. 2007, 144, 85–94. [Google Scholar] [CrossRef]
- Padovan, M.D.; Brook, R.M.; Barrios, M.; Cruz-Castillo, J.B.; Vilchez-Mendoza, S.J.; Costa, A.N.; Rapidel, B. Water loss by transpiration and soil evaporation in coffee shaded by Tabebuia rosea Bertol. and Simarouba glauca dc. compared to unshaded coffee in sub-optimal environmental conditions. Agric. For. Meteorol. 2018, 248, 1–14. [Google Scholar] [CrossRef]
- Rigal, C.; Xu, J.; Hu, G.; Qiu, M.; Vaast, P. Coffee production during the transition period from monoculture to agroforestry systems in near optimal growing conditions, in Yunnan Province. Agric. Syst. 2019, 177, 102696. [Google Scholar] [CrossRef]
- Muñoz-Villers, L.E.; Geris, J.; Alvarado-Barrientos, M.S.; Holwerda, F.; Dawson, T. Coffee and shade trees show complementary use of soil water in a traditional agroforestry 1377 ecosystem. Hydrol. Earth Syst. Sci. 2020, 24, 1649–1668. [Google Scholar] [CrossRef]
- Koutouleas, A.; Sarzynski, T.; Bordeaux, M.; Bosselmann, A.S.; Campa, C.; Etienne, H.; Turreira-García, N.; Rigal, C.; Vaast, P.; Ramalho, J.C.; et al. Shaded-coffee: A nature-based strategy for coffee production under climate change? A review. Front. Sustain. Food Syst. 2022, 6. [Google Scholar] [CrossRef]
- Beer, H. Advantages, disadvantages and desirable characteristics of shade for coffee, cacao and tea. Agrofor. Syst. 1987, 5, 3–13. [Google Scholar] [CrossRef]
- Courbier, S.; Pierik, R. Canopy light quality modulates stress responses in plants. iScience 2019, 22, 441–452. [Google Scholar] [CrossRef]
- De Carvalho, A.F.; Fernandes-Filho, E.I.; Daher, M.; de Carvalho Gomes, L.; Cardoso, I.M.; Fernandes, R.; Schaefer, C.E.G.R. Microclimate and soil and water loss in shaded and unshaded agroforestry coffee systems. Agrofor. Syst. 2021, 95, 119–134. [Google Scholar] [CrossRef]
- Jiménez, A.E.; Golberg, D. Estudios ecologicos del agroecosistema cafetalero. III. Efecto de diferentes estructuras vegetales sobre el balance hidrico del cafetal. In Estudios Ecologicos en Agroecosistema Cafetalero; Jiménez, A.E., Gomez, P.A., Eds.; Editora Continental: Ciudad de Mexico, MX, USA, 1982; pp. 39–54. [Google Scholar]
- Muschler, R. Shade improves coffee quality in a sub-optimal coffee-zone of Costa Rica. Agrofor. Syst. 2001, 51, 131–139. [Google Scholar] [CrossRef]
- Bertrand, B.; Villegas Hincapié, A.M.; Marie, L.; Breitler, J.-C. Breeding for the main agricultural farming of arabica coffee. Front. Sustain. Food Syst. 2021, 5. [Google Scholar] [CrossRef]
- Jezeer, R.E.; Verweij, P.A.; Santos, M.; Boot, R.G. Shaded coffee and cocoa–double dividend for biodiversity and small-scale farmers. Ecol. Econ. 2017, 140, 136–145. [Google Scholar] [CrossRef]
- Choudhary, A.; Rijhwani, S. A review of multi-dimensional benefits of innovative agricultural practices with special reference to Agroforestry and agrosilvipastoral system. Eco. Env. Cons. 2021, 27. [Google Scholar]
- DaMatta, F. Ecophysiological constraints on the production of shaded and unshaded coffee: A review. Field Crop. Res. 2004, 86, 99–114. [Google Scholar] [CrossRef]
- Harvey, C.A.; Pritts, A.A.; Zwetsloot, M.J.; Jansen, K.; Pulleman, M.M.; Armbrecht, I.; Avelino, J.; Barrera, J.F.; Bunn, C.; García, J.H.; et al. Transformation of coffee-growing landscapes across Latin America. A review. Agron. Sustain. Dev. 2021, 41, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Anthony, F.; Combes, M.C.; Astorga, C.; Bertrand, B.; Graziosi, G.; Lashermes, P. The origin of cultivated Coffea arabica L. varieties revealed by AFLP and SSR markers. Theor. Appl. Genet. 2002, 104, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Scalabrin, S.; Toniutti, L.; Di Gaspero, G.; Scaglione, D.; Magris, G.; Vidotto, M.; Pinosio, S.; Cattonaro, F.; Magni, F.; Jurman, I.; et al. A single polyploidization event at the origin of the tetraploid genome of Coffea arabica is responsible for the extremely low genetic variation in wild and cultivated germplasm. Sci. Rep. 2020, 10, 4642. [Google Scholar] [CrossRef]
- Vaast, P.; Bertrand, B.; Perriot, J.; Guyot, B.; Génard, M. Fruit Thinning and Shade Improve Bean Characteristics and Beverage Quality of Coffee (Coffea arabica L.) under Optimal Conditions. J. Sci. Food Agric. 2006, 86, 197–204. [Google Scholar] [CrossRef]
- Van der Vossen, H.; Bertrand, B.; Charrier, A. Next generation variety development for sustainable production of arabica coffee (Coffea arabica L.): A review. Euphytica 2015, 204, 243–256. [Google Scholar] [CrossRef]
- Galiè, A.; Jiggins, J.; Struik, P.C.; Grando, S.; Ceccarelli, S. Women’s empowerment through seed improvement and seed governance: Evidence from participatory barley breeding in pre-war Syria. J. Life Sci. 2017, 81, 1–8. [Google Scholar] [CrossRef]
- Struik, P.C.; Kuyper, T.W. Editorial overview: Sustainable intensification to feed the world: Concepts, technologies and trade-offs. Curr. Opin. Environ. Sustain. 2014, 8, VI–VIII. [Google Scholar] [CrossRef]
- Suso, M.J.; Bebeli, P.J.; Christmann, S.; Mateus, C.; Negri, V.; Pinheiro de Carvalho, M.A.; Torricelli, R.; Veloso, M.M. Enhancing legume ecosystem services through an understanding of plant–pollinator interplay. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- De Deyn, G.B.; Cornelissen, J.H.; Bardgett, R.D. Plant functional traits and soil carbon sequestration in contrasting biomes. Ecol. Lett. 2008, 11, 516–531. [Google Scholar] [CrossRef] [PubMed]
- Deru, J.G.C.; Schilder, H.; van der Schoot, J.R.; van Eekere, N. Genetic differences in root mass of Lolium perenne varieties under field conditions. Euphytica 2014, 199, 223–232. [Google Scholar] [CrossRef]
- Perez, J.E.; Carrion, V.J.; Bosse, M.; Ferrão, L.F.V.; de Hollander, M.; Garcia, A.A.F.; Ramirez, C.A.; Mendes, R.; Raaijmakers, J.M. Linking rhizosphere microbiome composition of wild and domesticated Phaseolus vulgaris to genotypic and root phenotypic traits. ISME J. 2017, 11. [Google Scholar] [CrossRef]
- Van Bueren, E.T.L.; Struik, P.C. Diverse concepts for breeding for nitrogen use efficiency. A review. Agron. Sustain. Dev. 2017, 37, 50. [Google Scholar] [CrossRef]
- Gewin, V. Food: An underground revolution. Nature 2010, 466, 552–553. [Google Scholar] [CrossRef]
- Galván, G.A.; Kuyper, T.W.; Burger, K.; Keizer, L.C.P.; Hoekstra, R.F.; Kik, C.; Scholten, O.E. Genetic analysis of the interaction between Allium species and arbuscular mycorrhizal fungi. Theor. Appl. Genet. 2011, 122, 947–960. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Valladares, F.; Van Kleunen, M. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.; Monaco, L.C. The breeding of Arabica coffee. In Outlines of Perennial Crop Breeding in the Tropics. Misc. Pap. Agric. Univ. Wageningen; Fewerda, F.P., Wit, F., Eds.; Veenman: Wageningen, The Netherlands, 1969; pp. 198–216. [Google Scholar]
- Carvalho, A. Principles and practice of coffee plant breeding for productivity and quality factors: Coffea arabica. In Coffee. Vol. 4: Agronomy; Clarke, R.J., Macrae, R., Eds.; Elsevier: London, UK, 1985; pp. 129–166. [Google Scholar]
- Van der Vossen, H. Coffee selection and breeding. In Coffee Botany, Biochemistry and Production of Beans and Beverage; Clifford, C., Willson, J., Eds.; Croom Helm: London, UK, 1985; pp. 48–96. [Google Scholar]
- The Coffee Guide. 2021. Available online: https://www.thecoffeeguide.org/ (accessed on 15 February 2022).
- Voora, V.; Bermúdez, S.; Larrea, C.; Baliño, S. Sustainable Commodities Marketplace Series 2019; International Institute for Sustainable Development; Veenman: Wageningen, The Netherlands, 2019. [Google Scholar]
- Gallais, A. Hétérosis et Variétés Hybrides en Amélioration des Plantes; Editions Quae: Paris, France, 2009. [Google Scholar]
- Ameha, M. Heterosis and Arabica coffee breeding in Ethiopia. Plant Breed. Abstr. 1990, 60, 593–598. [Google Scholar]
- Walyaro, D.J. Considerations in Breeding for Improved Yield and Quality (Coffea arabica L.). Ph.D. Thesis, Agricultural University Wageningen, Wageningen, The Netherlands, 1983. [Google Scholar]
- Bellachew, B. Arabica coffee breeding in Ethiopia: A review. In Proceedings of the 17th ASIC International Conference on Coffee Science, Nairobi, Kenya, 20–25 July 1997; pp. 406–414. [Google Scholar]
- Bertrand, B.; Aguilar, G.; Santacreo, R.; Anthony, F.; Etienne, H.; Eskes, A.B.; Charrier, A. Comportement d’hybrides F1 de Coffea arabica pour la vigeur, la production et la fertilité en Amérique Centrale. In Proceedings of the 17th ASIC International Conference on Coffee Science, Nairobi, Kenya, 20–25 July 1997; pp. 415–423. [Google Scholar]
- Teri, J.; Kilambo, D.; Nyange, N.E.; Nzallawahe, T.S.; Chipun-gahelo, G.S.; Kipokola, T.P.; Kullaya, I.K. Improved arabica varieties for the benefit of Tanzanian coffee growers. In Proceedings of the 20th ASIC International Conference on Coffee Science, Bangalore, India, 11–15 October 2004; pp. 1187–1191. [Google Scholar]
- Bertrand, B.; Etienne, H.; Cilas, C.; Charrier, A.; Baradat, P. Coffea arabica hybrid performance for yield, fertility and bean weight. Euphytica 2005, 141, 255–262. [Google Scholar] [CrossRef]
- Bertrand, B.; Alpizar, E.; Llara, L.; SantaCreo, R.; Hidalgo, M.; Quijano, J.M.; Charmetant, P.; Montagnon, C.; Georget, F.; Etienne, H. Performance of Coffea arabica F1 hybrids in agro- forestry and full-sun cropping systems in comparison with American pure line varieties. Euphytica 2011, 181, 147–158. [Google Scholar] [CrossRef]
- Bertrand, B.; Vaast, P.; Alpizar, E.; Etienne, H.; Davrieux, F.; Charmetant, P. Comparison of bean biochemical composition and beverage quality of arabica hybrids involving Sudanese-Ethiopian origins with traditional varieties at various elevations in Central America. Tree Physiol. 2006, 26, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Etienne, H.; Bertrand, B.; Montagnon, C.; Bobadilla Landey, R.; Dechamp, E.; Jourdan, I.; Alpizar, E.; Malo, E.; Georget, F. Un exemple de transfert de technologie réussi dans le domaine de la micropropagation: La multiplication de Coffea arabica par embryogenèse somatique. Cah. Agric. 2012, 21, 115–124. [Google Scholar] [CrossRef]
- Bobadilla Landey, R.; Cenci, A.; Georget, F.; Bertrand, B.; Camayo, G.; Dechamp, E.; Simpson, J.; Herrera, J.C.; Santoni, S.; Lashermes, P.; et al. High genetic and epigenetic stability in Coffea arabica plants derived from embryogenic suspensions and secondary embryogenesis as revealed by AFLP, MSAP and the phenotypic variation rate. PLoS ONE 2013, 8, e56372. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Sugano, S.; Osakabe, K. Genome engineering of woody plants: Past, present and future. J. Wood Sci. 2016, 62, 217–225. [Google Scholar] [CrossRef]
- Eskes, A.; Leroy, T. Coffee selection and breeding. In Coffee: Growing, Processing, Sustainable Production: A Guidebook for Growers, Processors, Traders, and Researchers; WILEY-VCH Verlag GmbH & Co. KgaA: Weinheim, Germany, 2004. [Google Scholar] [CrossRef]
- Mishra, M.K.; Slater, A. Recent advances in the genetic transformation of coffee. Biotechnol. Res. Int. 2012, 10. [Google Scholar] [CrossRef] [PubMed]
- Marie, L.; Abdallah, C.; Campa, C.; Courtel, P.; Bordeaux, M.; Navarini, L.; Lonzarich, V.; Bosselmann, A.S.; Turreira-Garcia, N.; Alpizar, E.; et al. G × E Interactions on Yield and Quality in Coffea arabica: New F1 Hybrids Outperform American Cultivars. Euphytica 2020, 216, 1–17. [Google Scholar] [CrossRef]
- Turreira, N. Farmers’ perceptions and adoption of Coffea arabica F1 hybrids in Central America. World Dev. Sustain. 2022, 1, 100007. [Google Scholar] [CrossRef]
- Etienne, H.; Breton, D.; Breitler, J.-C.; Bertrand, B.; Déchamp, E.; Awada, R.; Marraccini, P.; Leran, S.; Alpizar, E.; Campa, C.; et al. Coffee somatic embryogenesis: How did research, experience gained and innovations promote the commercial propagation of elite clones from the two cultivated species? Front. Plant Sci. 2018, 9, 1630. [Google Scholar] [CrossRef]
- Georget, F.; Courtel, P.; Garcia, E.M.; Hidalgo, M.; Alpizar, E.; Breitler, J.-C.; Bertrand, B.; Etienne, H. Somatic embryogenesis-derived coffee plantlets can be efficiently propagated by horticultural rooted mini-cuttings: A boost for somatic embryogenesis. Sci. Hortic. 2017, 21, 177–185. [Google Scholar] [CrossRef]
- Georget, F.; Marie, L.; Alpizar, E.; Courtel, P.; Bordeaux, M.; Hidalgo, J.M.; Marraccini, P.; Breitler, J.-C.; Déchamp, E.; Poncon, C.; et al. Starmaya: The first Arabica F1 coffee hybrid produced using genetic male sterility. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Debaeke, P.; Quilot-Turion, B. Conception d’idéotypes de plantes pour une agriculture durable. In Collection École-Chercheurs; FormaSciences: Paris, France, 2014. [Google Scholar]
- Donald, C. The breeding of crop ideotypes. Euphytica 1968, 17, 385–403. [Google Scholar] [CrossRef]
- Rasmusson, D. An Evaluation of Ideotype Breeding. Crop Sci. 1987, 27, 1140–1146. [Google Scholar] [CrossRef]
- Hamblin, J. The Ideotype Concept: Useful or Outdated? In Book Series: ASA, CSSA, and SSSA Books; Buxton, D.R., Shibles, R., Forsberg, R.A., Blad, B.L., Asay, K.H., Paulsen, G.M., Wilson, R.F., Eds.; Crop Science Society of America: Madison, WI, USA, 1993. [Google Scholar] [CrossRef]
- Gauffreteau, A. Using ideotypes to support selection and recommendation of varieties. OCL 2018, 25, D602. [Google Scholar] [CrossRef]
- Semenov, M.; Stratonovitch, P. Designing high-yielding wheat ideotypes for a changing climate. Food Energy Secur. 2013, 2, 185–196. [Google Scholar] [CrossRef]
- Andrivon, D.; Giorgetti, C.; Baranger, A.; Calonnec, A.; Cartolaro, P.; Faivre, R.; Guyader, S.; Lauri, P.E.; Lescourret, F.; Parisi, L.; et al. Defining and designing plant architectural ideotypes to control epidemics? Eur. J. Plant Pathol. 2013, 135, 611–617. [Google Scholar] [CrossRef]
- Martre, P.; Quilot-Turion, B.; Luquet, D.; Ould-Sidi Memmah, M.M.; Chenu, K.; Debaeke, P. Model-Assisted Phenotyping and Ideotype Design. Crop Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2015. [Google Scholar] [CrossRef]
- Avelino, J.; Cristancho, M.; Georgiou, S.; Imbach, P.; Aguilar, L.; Bornemann, G.; Läderach, P.; Anzueto, F.; Hruska, A.; Morales, C. The coffee rust crises in Colombia and Central America (2008–2013): Impacts, plausible causes and proposed solutions. Food Secur. 2015, 7, 303–321. [Google Scholar] [CrossRef]
- Toniutti, L.; Breitler, J.-C.; Etienne, H.; Campa, C.; Doulbeau, S.; Urban, L.; Lambot, C.; Pinilla, J.-C.H.; Bertrand, B. Influence of environmental conditions and genetic background of Arabica Coffee (C. arabica L.) on Leaf Rust (Hemileia vastatrix) pathogenesis. Front. Plant Sci. 2017, 8, 2025. [Google Scholar] [CrossRef]
- Rhineya, K.; Guidob, Z.; Knudsonc, C.; Avelino, J.; Bacong, C.; Leclerc, G.; Aime, C.; Bebber, D. Epidemics and the future of coffee production. Proc. Natl. Acad. Sci. USA 2021, 118, e2023212118. [Google Scholar] [CrossRef]
- Carvalho, A. Advances in Coffee Production Technology Recent Advances in Our Knowledge of Coffee Trees. Coffee Tea Ind. 1958, 81, 37–41. [Google Scholar]
- Elias, A.A.; Busov, V.B.; Kosola, K.R.; Ma, C.; Etherington, E.; Shevchenko, O.; Gandhi, H.; Pearce, D.W.; Rood, S.; Strauss, S.H. Green revolution trees: Semidwarfism transgenes modify gibberellins, promote root growth, enhance morphological diversity, and reduce competitiveness in hybrid poplar. Plant Physiol. 2012, 160, 1130–1144. [Google Scholar] [CrossRef]
- Battistini, A.; Battistini, G. VICTOR®: A semi-dwarfing cherry rootstock for dry conditions. Acta. Hortic. Acta Hortic. 2005, 667, 189–190. [Google Scholar] [CrossRef]
- Moncada, P.; Cortina, H.; Alarcón, R. Cup quality and yield evaluation of the Ethiopian germplasm collection of Coffea arabica L. J. Agric. Rural. Res. 2019, 3, 100–126. [Google Scholar]
- Mbebi, A.J.; Breitler, J.-C.; Bordeaux, M.; Sulpice, R.; McHale, M.; Castillo, J.A.; Toniutti, L.; Bertrand, B.; Nikoloski, Z. A comparative analysis of genomic and phenomic predictions of growth-related traits in three-way coffee hybrids. Genes Genomes Genet. 2022. [Google Scholar]
- Stirbet, A.; Lazár, D.; Kromdijk, J.; Govindjee. Chlorophyll a Fluorescence Induction: Can Just a One-Second Measurement Be Used to Quantify Abiotic Stress Responses? Photosynthetica 2018, 56, 86–104. [Google Scholar] [CrossRef]
- Toniutti, L.; Breitler, J.-C.; Guittin, C.; Doulbeau, S.; Etienne, H.; Campa, C.; Lambot, C.; Herrera Pinilla, J.-C.; Bertrand, B. An Altered Circadian Clock Coupled with a Higher Photosynthesis Efficiency Could Explain the Better Agronomic Performance of a New Coffee Clone When Compared with a Standard Variety. Int. J. Mol. Sci. 2019, 20, 736. [Google Scholar] [CrossRef]
- Castillo, Z.; Lopez, A. Nota sobre el efecto de la intensidad de la luz en la floracion del café. Cenicafé 1966, 17, 51–60. [Google Scholar]
- Cannell, M. Crop physiological aspects of coffee bean yield—A review. Kenya Coffee 1976, 41. [Google Scholar]
- Cannell, M. Physiology of coffee crop. In Coffee, Botany, Biochemistry and Production of Beans and Beverage; Clifford, M.N., Willson, K.C., Eds.; Croom Helm: London, UK, 1985; pp. 108–134. [Google Scholar]
- Drinnan, J.; Menzel, C. Temperature affects vegetative growth and flowering of coffee (Coffea arabica L.). J. Hortic. Sci. 1995, 70, 25–34. [Google Scholar] [CrossRef]
- Montagnon, C.; Marraccini, P.; Bertrand, B. Breeding for coffee quality. In Specialty Coffee: Managing Quality. International Plant Nutrition Institute, Southeast Asia Program; IPNI (The International Plant Nutrition Institute): Norcross, GA, USA, 2012; pp. 93–122. [Google Scholar]
- De Rosso, M.; Lonzarich, V.; Navarini, L.; Flamini, R. Identification of new glycosidic terpenols and norisoprenoids (aroma precursors) in C. arabica L. green coffee by using a high-resolution mass spectrometry database developed in grape metabolomics. Curr. Res. Food Sci. 2022, 5, 336–344. [Google Scholar] [CrossRef]
- Rhinehart, R. What Is Specialty Coffee? 2009. Available online: http://www.scaa.org/?page=RicArtp1 (accessed on 15 March 2022).
- Bacon, C.M.; Ernesto Méndez, V.; Gómez, M.E.F.; Stuart, D.; Flores, S.R.D. Are sustainable coffee certifications enough to secure farmer livelihoods? The millenium development goals and Nicaragua’s Fair Trade cooperatives. Globalizations 2008, 5, 259–274. [Google Scholar] [CrossRef]
- Bertrand, B.; Guyot, B.; Anthony, F.; Lashermes, P. Impact of the Coffea canephora gene introgression on beverage quality of C. arabica. Theor. Appl. Genet. 2003, 107, 387–394. [Google Scholar] [CrossRef] [PubMed]
- McCook, S.; Vandermeer, J. The big rust and the red queen: Long- term perspectives on coffee rust research. Phytopathology 2015, 105, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Marie, L.; Breitler, J.-C.; Bamogo, P.; Bordeaux, M.; Lacombe, S.; Rios, M.; Lebrun, M.; Boulanger, R.; Lefort, E.; Nakamura, S.; et al. Terpene synthase promotes limonene accumulation at high elevation in Geisha, the Queen of the varieties for ‘Specialty coffee’ market. Plant Cell Environ. 2022; submitted. [Google Scholar]
- DaMatta, F.; Cavatte, P.; Martins, S. Coffee physiology: Growth, yield and quality. In Specialty Coffee: Managing Quality; Laderach, P., Pohlan, J., Cock, J.H., Eds.; International Plant Nutrition Institute: Norcross, GA, USA, 2012; pp. 75–91. [Google Scholar]
- Deivasigamani, S.; Swaminathan, C. Evaluation of seed test weight on major field crops. Int. J. Res. Stud. Agric. Sci. 2018, 4, 8–11. [Google Scholar] [CrossRef]
- Kathurima, C.W.; Gichimu, B.M.; Kenji, G.M.; Muhoho, S.M.; Boulanger, R. Evaluation of beverage quality and green bean physical characteristics of selected Arabica coffee genotypes in Kenya. Afr. J. Food Sci. 2009, 3. [Google Scholar] [CrossRef]
- Kathurima, C.W.; Kenji, G.M.; Muhoho, S.M.; Boulanger, R.; Davrieux, F. Discrimination of Coffea arabica hybrids of the composite cultivar ruiru 11 by sensorial evaluation and biochemical characterization. Adv. J. Food Sci. Technol. 2010, 2, 148–154. [Google Scholar]
- Ribeiro, J.S.; Ferreira, M.M.C.; Salva, T.J.G. Chemometric models for the quantitative descriptive sensory analysis of Arabica coffee beverages using near infrared spectroscopy. Talanta 2011, 83, 1352–1358. [Google Scholar] [CrossRef]
- Viapiana, A.; Maggi, F.; Kaszuba, M.; Konieczynski, P.; Wesolowski, M. Quality assessment of Coffea arabica commercial samples. Nat. Prod. Res. 2019, 34, 3154–3157. [Google Scholar] [CrossRef]
- Eskes, A.B. Natural enemies and biological control. In Coffee Rust: Epidemiology, Resistance and Management; Kushalappa, A.C., Eskes, A.B., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1989; pp. 161–169. [Google Scholar]
- Rozo, Y.; Escobar, C.; Gaitán, A.; Cristancho, M. Aggressiveness and genetic diversity of Hemileia vastatrix during an epidemic in Colombia. J. Phytopathol. 2012, 160, 732–740. [Google Scholar] [CrossRef]
- Silva, M.C.; Varzea, V.; Guerra-Guimaraes, L.; Gil Azinheira, H.; Fernandez, D.; Petitot, A.S.; Bertrand, B.; Lashermes, P.; Nicole, M. Coffee resistance to the main diseases: Leaf rust and coffee berry disease. Braz. J. Plant Physiol. 2006, 18, 119–147. [Google Scholar] [CrossRef]
- Herrera, P.J.C.; Alvarado, A.G.; Hernando, A.; Cortina, G.; Combes, M.C.; Romer, G.G.; Lashermes, P. Genetic analysis of partial resistance to coffee leaf rust (Hemileia vastatrix Berk & Br.) introgressed into the cultivated Coffea arabica L. from the diploid C. canephora species. Euphytica 2008, 167, 57–67. [Google Scholar] [CrossRef]
- Bertrand, B.; Marraccini, P.; Villain, L.; Breitler, J.C.; Etienne, H. Healthy tropical plants to mitigate the impact of climate change—As exemplified in coffee. In Climate Change and Agriculture Worldwide; Torquebiau, E., Manley, D., Cowan, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 83–95. [Google Scholar]
- Dordas, C. Role of nutrients in controlling plant diseases in sustainable agriculture. A review. Agron. Sustain. Dev. 2008, 28, 33–46. [Google Scholar] [CrossRef]
- Echeverria-Beirute, F.; Murray, S.C.; Klein, P.; Kerth, C.; Miller, R.; Bertrand, B. Rust and thinning management effect on cup quality and plant performance for two cultivars of Coffea arabica L. J. Agric. Food Chem. 2018, 66, 5281–5292. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Kim, E.D.; Ha, M.; Lackey, E.; Liu, J.; Zhang, Y.; Sun, Q.; Chen, Z. Altered circadian rhythms regulate growth vigour in hybrids and allopolyploids. Nature 2009, 457, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.; Hu, W.; Zhang, B.; Xing, Y. The regulatory network mediated by circadian clock genes is related to heterosis in rice. J. Integr. Plant Biol. 2015, 57, 300–312. [Google Scholar] [CrossRef]
- Ko, D.K.; Rohozinski, D.; Song, Q.; Taylor, S.H.; Juenger, T.E.; Harmon, F.G.; Chen, Z. Temporal Shift of Circadian-Mediated Gene Expression and Carbon Fixation Contributes to Biomass Heterosis in Maize Hybrids. PLoS Genet. 2016, 12, e1006197. [Google Scholar] [CrossRef]
- Avelino, J.; Zelaya, H.; Merlo, A.; Pineda, A.; Ordóñez, M.; Savary, S. The intensity of a coffee rust epidemic is dependent on production situations. Ecol. Model. 2006, 197, 431–447. [Google Scholar] [CrossRef]
- Carvajal, J. Cafeto: Cultivo y Fertilizacion; Instituto de la Potasa: Berna, Swtizerland, 1984. [Google Scholar]
- Salamanca-Jimenez, A.; Doane, T.; Horwath, W. Nitrogen Use Efficiency of Coffee at the Vegetative Stage as Influenced by Fertilizer Application Method. Front. Plant Sci. 2017, 8, 223. [Google Scholar] [CrossRef]
- Bornemisza, E. Nitrogen Cycling in Coffee Plantations. Plant Soil 1982, 67, 241–246. [Google Scholar] [CrossRef]
- Aranguren, J.; Escalante, G.; Herrera, R. Nitrogen Cycle of Tropical Perennial Crops Under Shade Trees: Coffee. Plant Soil 1982, 67, 148–154. [Google Scholar] [CrossRef]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. N uptake and distribution in crops: An agronomical and ecophysiological perspective. J. Exp. Bot. 2002, 53, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Thorup-Kristensen, K. Effect of deep and shallow root systems on the dynamics of soil inorganic N during 3-year crop rotations. Plant Soil 2006, 288, 233–248. [Google Scholar] [CrossRef]
- Philippot, L.; Sport, A.; Hénault, C.; Bru, D.; Bizouard, F.; Jones, C.M.; Sarr, A.; Maron, P.-A. Loss in microbial diversity affects nitrogen cycling in soil. ISME J. 2013, 7, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, N.; Belec, C. Adapting nitrogen fertilization to unpredictable seasonal conditions with the least impact on the environment. HortTechnology 2006, 16, 408–412. [Google Scholar] [CrossRef]
- Dresbøll, D.B.; Thorup-Kristensen, K. Will breeding for nitrogen use efficient crops lead to nitrogen use efficient cropping systems?: A simulation study of G× E× M interactions. Euphytica 2014, 199, 97–117. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K.; Dresboll, D.B.; Kristensen, H.L. Crop yield, root growth, and nutrient dynamics in a conventional and three organic cropping systems with different levels of external inputs and N re-cycling through fertility building crops. Eur. J. Agron. 2012, 37, 66–82. [Google Scholar] [CrossRef]
- Chan-Navarrete, R.; Dolstra, O.; van Kaauwen, M.; Lammerts van Bueren, E.T.; van der Linden, C.G. Genetic map construction and QTL analysis of nitrogen use efficiency in spinach (Spinacia oleracea L.). Euphytica 2016, 208, 621–636. [Google Scholar] [CrossRef]
- Zhang, D.; Vega, F.E.; Solano, W.; Su, F.; Infante, F.; Meinhardt, L.W. Selecting a core set of nuclear SNP markers for molecular characterization of Arabica coffee (Coffea arabica L.) genetic resources. Conserv. Genet. Resour. 2021, 13, 329–335. [Google Scholar] [CrossRef]
- Montagnon, C.; Mahyoub, A.; Solano, W.; Sheibani, F. Unveiling a unique genetic diversity of cultivated Coffea arabica L. in its main domestication center: Yemen. Genet. Resour. Crop Evol. 2021, 68, 2411–2422. [Google Scholar] [CrossRef]
- Bertrand, B.; Breitler, J.-C.; Georget, F.; Penot, E.; Bordeaux, M.; Marraccini, P.; Leran, S.; Campa, C.; Bonato, O.; Villain, L.; et al. New varieties for innovative agroforestry coffee systems. In The Agroecological Transition of Agricultural Systems in the Global South; Côte, F.-X., Poirier-Magona, E., Perret, S., Roudier, P., Bruno, R., Thirion, M.-C., Eds.; Ed Quae: Versailles, France, 2019; pp. 161–176. [Google Scholar]
- Rebolledo, M.; Luquet, D.; Courtois, B.; Henry, A.; Soulié, J.-C.; Rouan, L.; Dingkuhn, M. Can early vigour occur in combination with drought tolerance and efficient water use in rice genotypes? Funct. Plant Biol. 2013, 40, 582–594. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
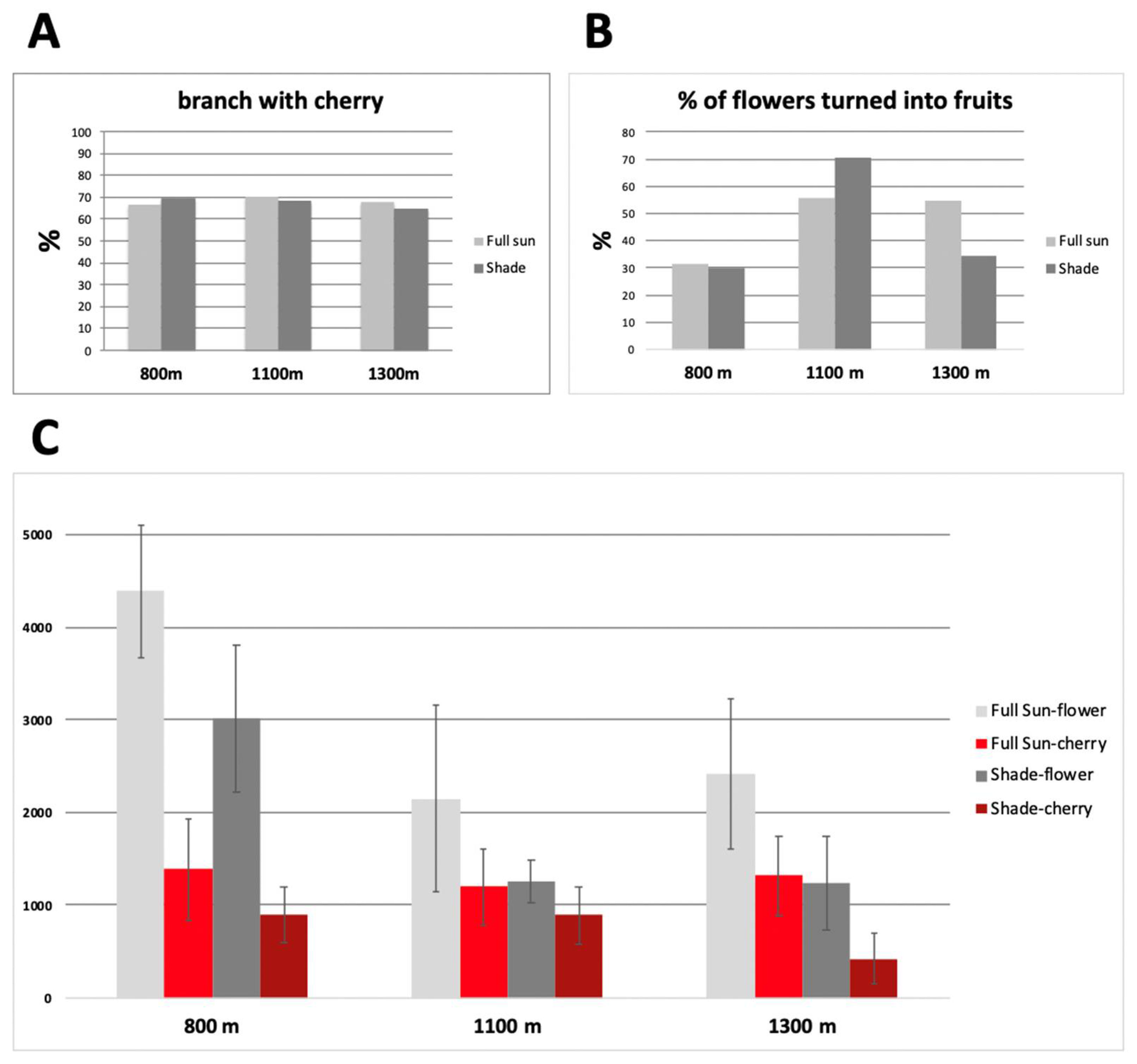{kind=link}
{kind=link}
| Cropping System | Agroforestry | |
|---|---|---|
| Crop management | Organic or «passive organic» | |
| Trait ranking | Traits | Phenotypic goal based on |
| Trait 1 | Productivity | yield (green coffee kg/plant) |
| Trait 2 | Quality | Cupping score, bean size, W100 * |
| Trait 3 | Plant health | vigor, high PI ** |
| Trait 4 | Low fertilization | NUE, P and K |
| Individual Traits | Phenotypic Goal |
|---|---|
| Productivity | |
| Yield (Kg of green coffee/plant) | >0.5 |
| Floriation intensity | >4000 flowers/plant |
| Flowers to fruit conversion | >75% |
| Tree volume (m3) | 0.5–1 |
| Collar diameter (6 months old) | 18–20 mm |
| Crop density | 4000–5000 plants/ha |
| Plant height | 180–250 cm |
| Precocity | 2 years |
| Quality | |
| Bean size (16–20) | >85% |
| 100-grain weight (g) | 20–22 |
| Cupping score | >85 |
| Plant health | |
| Chlorophyll content (mg/gFW) | >2 |
| Performance Index | highest |
| Rust sensitivity | <1(sporulating lesions reaching 1 to 5% of the total leaf area [110]) |
| Organic production | |
| Nitrogen use efficiency (NUE) | 25–50 kg N ha−1 annually |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Breitler, J.-C.; Etienne, H.; Léran, S.; Marie, L.; Bertrand, B. Description of an Arabica Coffee Ideotype for Agroforestry Cropping Systems: A Guideline for Breeding More Resilient New Varieties. Plants 2022, 11, 2133. https://doi.org/10.3390/plants11162133
Breitler J-C, Etienne H, Léran S, Marie L, Bertrand B. Description of an Arabica Coffee Ideotype for Agroforestry Cropping Systems: A Guideline for Breeding More Resilient New Varieties. Plants. 2022; 11(16):2133. https://doi.org/10.3390/plants11162133
Chicago/Turabian StyleBreitler, Jean-Christophe, Hervé Etienne, Sophie Léran, Lison Marie, and Benoit Bertrand. 2022. "Description of an Arabica Coffee Ideotype for Agroforestry Cropping Systems: A Guideline for Breeding More Resilient New Varieties" Plants 11, no. 16: 2133. https://doi.org/10.3390/plants11162133