
Centromere-Specific Single-Copy Sequences of Secale Species

Abstract
:1. Introduction
2. Results
2.1. Cloning of Single-Copy Sequences
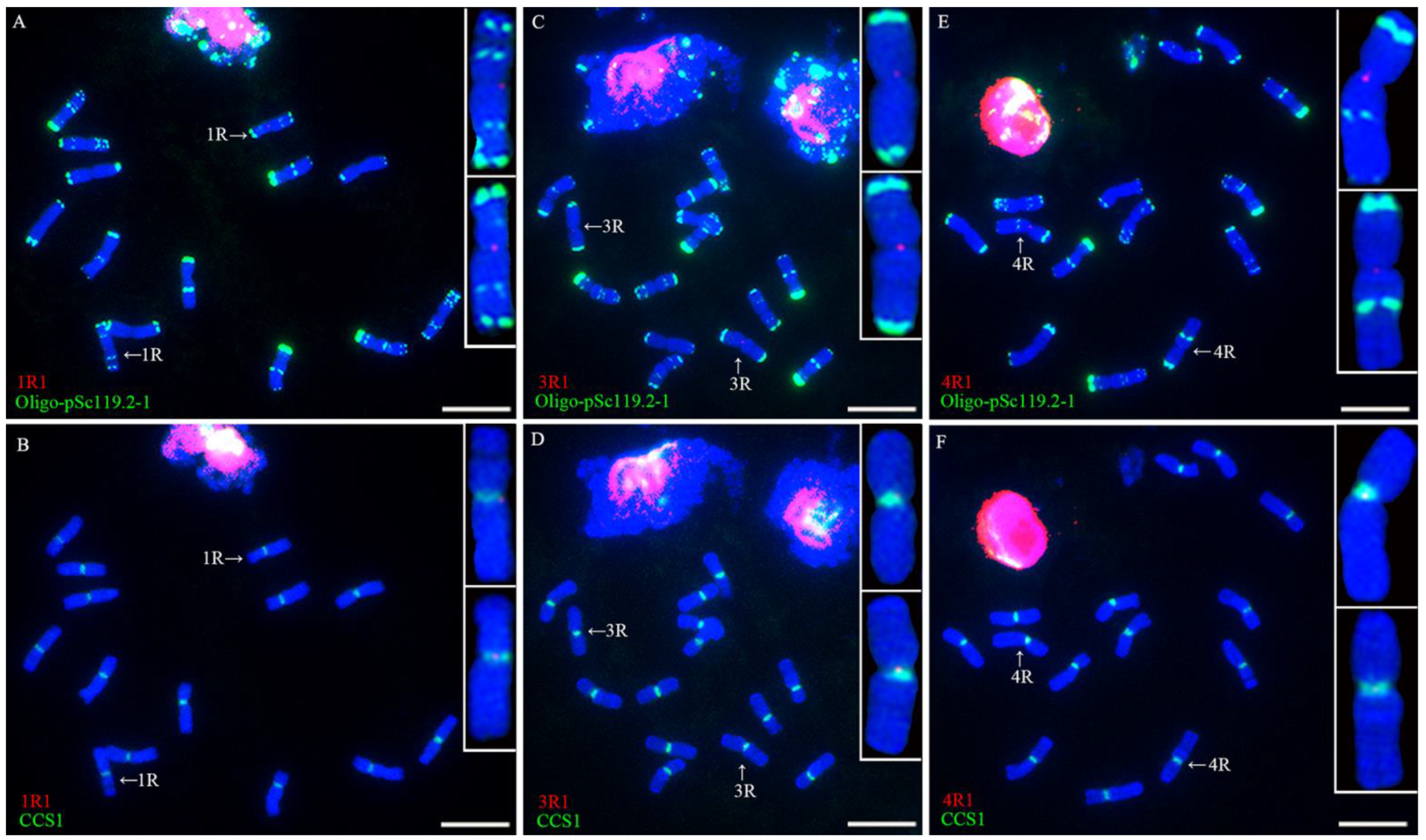
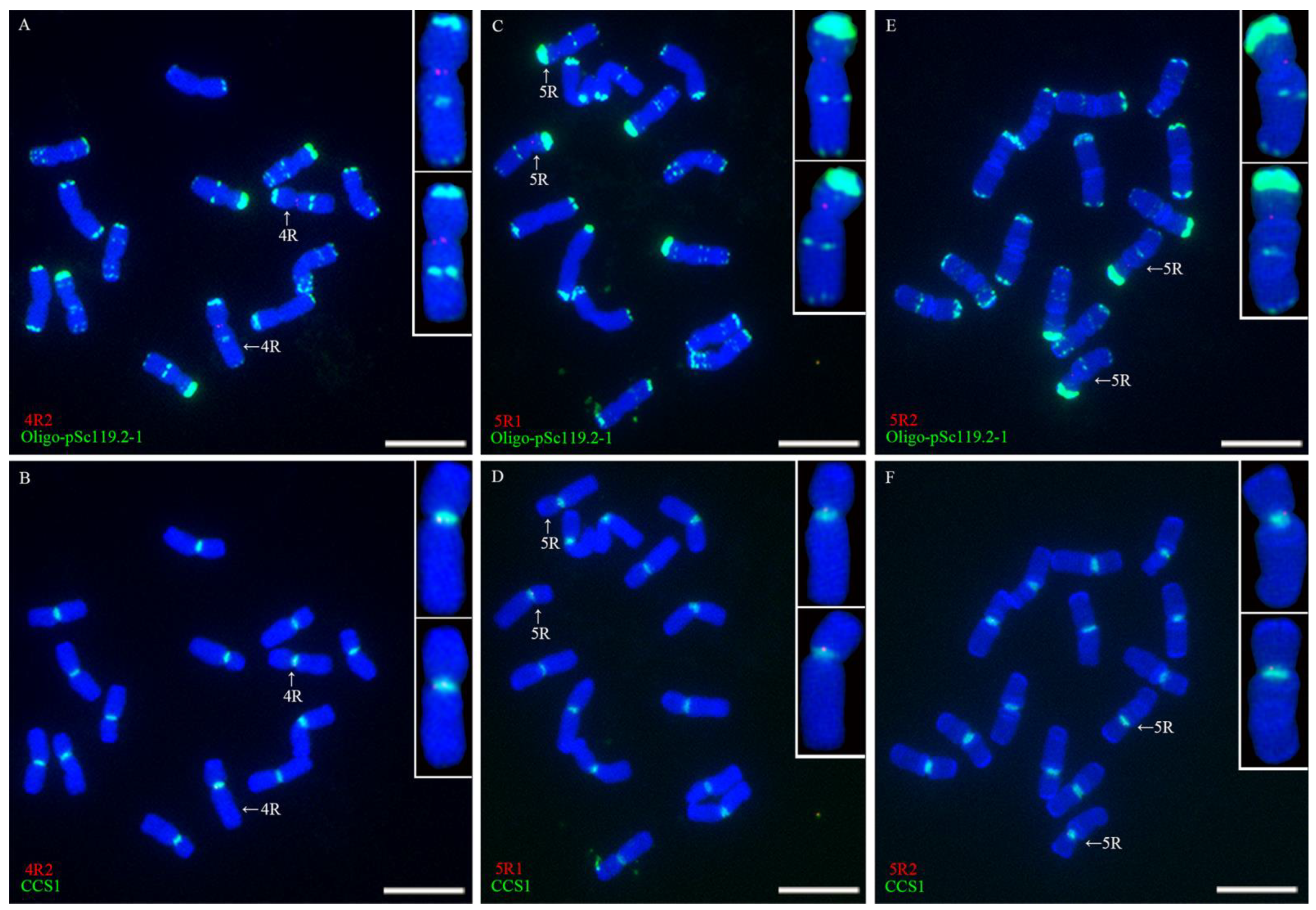
2.2. Location of Single-Copy Sequences on Rye Chromosomes
2.3. Different Centromeric Structure of Wheat-Rye 1BL.1RS Translocations
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Selection of Single-Copy Sequence
4.3. PCR Amplification and Sequence Cloning
4.4. Single-Copy FISH Analysis
4.5. Non-Denaturing FISH (ND-FISH)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borowska-Zuchowska, N.; Senderowicz, M.; Trunova, D.; Kolano, B. Tracing the evolution of the angiosperm genome from the cytogenetic point of view. Plants 2022, 11, 784. [Google Scholar] [CrossRef]
- Jiang, J. Fluorescence in situ hybridization in plants: Recent developments and future applications. Chromosome Res. 2019, 27, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bi, Y.; Wang, X.; Wang, Y.; Yang, S.; Zhang, Z.; Chen, J.; Lou, Q. Chromosome identification in Cucumis anguria revealed by cross-species single-copy gene FISH. Genome 2018, 61, 397–404. [Google Scholar] [CrossRef]
- Danilova, T.V.; Friebs, B.; Gill, B.S. Single-copy gene fluorescence in situ hybridization and genome analysis: Acc-2 loci mark evolutionary chromosomal rearrangements in wheat. Chromosoma 2012, 121, 597–611. [Google Scholar] [CrossRef] [PubMed]
- Danilova, T.V.; Friebs, B.; Gill, B.S. Development of a wheat single gene FISH map for analyzing homoeologous relationship and chromosomal rearrangements within the Triticeae. Theor. Appl. Genet. 2014, 127, 715–730. [Google Scholar] [CrossRef]
- Aliyeva-Schnorr, L.; Stein, N.; Houben, A. Collinearity of homoeologous group 3 chromosomes in the genus Hordeum and Secale cereale as revealed by 3H-derived FISH analysis. Chromosome Res. 2016, 24, 231–242. [Google Scholar] [CrossRef]
- Danilova, T.V.; Akhunova, A.R.; Akhunov, E.D.; Friebe, B.; Gill, B.S. Major structural genomic alterations can be associated with hybrid speciation in Aegilops markgrafii (Triticeae). Plant J. 2017, 92, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Said, M.; Hřibová, E.; Danilova, T.V.; Karafiátová, M.; Čížková, J.; Friebe, B.; Doležel, J.; Gill, B.S.; Vrána, J. The Agropyron cristatum karyotype, chromosome structure and cross-genome homoeology as revealed by fluorescence in situ hybridization with tandem repeats and wheat single-gene probes. Theor. Appl. Genet. 2018, 131, 2213–2227. [Google Scholar] [CrossRef]
- Said, M.; Hřibová, E.; Farkas, A.; Ivanizs, L.; Gaál, E.; Cápal, P.; Abrouk, M.; Martis-Thiele, M.M.; Kalapos, B.; Bartoš, J.; et al. Development of DNA markers from physically mapped loci in Aegilops comosa and Aegilops umbellulata using single-gene FISH and chromosome sequences. Front. Plant Sci. 2021, 12, 689031. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Liu, B.; Dou, Q. The Kengyiliahirsuta karyotype polymorphisms as revealed by FISH with tandem repeats and single-gene probes. Comp. Cytogenet. 2021, 15, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Nagaki, K.; Talbert, P.B.; Zhong, C.X.; Dawe, R.K.; Henikoff, S.; Jiang, J. Chromatin immunoprecipitation reveals that the 180-bp satellite repeat is the key functional DNA element of Arabidopsis thaliana centromeres. Genetics 2003, 163, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Francki, M.G. Identification of Bilby, a diverged centromeric Ty1-copia retrotransposon family from cereal rye (Secale cereale L.). Genome 2001, 44, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Dong, F.; Langdon, T.; Ouyang, S.; Buell, C.R.; Gu, M.; Blattner, F.R.; Jiang, J. Functional rice centromeres are marked by a satellite repeat and a centromere-specific retrotransposon. Plant Cell 2002, 14, 1691–1704. [Google Scholar] [CrossRef]
- Ananiev, E.V.; Phillips, R.L.; Rines, H.W. Chromosome-specific molecular organization of maize (Zea mays L.) centromeric regions. Proc. Natl. Acad. Sci. USA 1998, 95, 13073–13078. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Nasuda, S.; Dong, F.; Scherrer, C.W.; Woo, S.S.; Wing, R.A.; Gill, B.S.; Ward, D.C. A conserved repetitive DNA element located in the centromeres of cereal chromosomes. Proc. Natl. Acad. Sci. USA 1996, 93, 14210–14213. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Friebe, B.; Lukaszewski, A.J.; Gill, B.S. The centromere structure in Robertsonian wheat-rye translocation chromosomes indicates that centric breakage-fusion can occur at different positions within the primary constriction. Chromosoma 2001, 110, 335–344. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Y.; Su, H.; Guo, X.; Han, F. Centromere structure and function analysis in wheat–rye translocation lines. Plant J. 2017, 91, 199–207. [Google Scholar] [CrossRef]
- Karimi-Ashtiyani, R.; Schubert, V.; Houben, A. Only the rye derived part of the 1BL/1RS hybrid centromere incorporates CENH3 of wheat. Front. Plant Sci. 2021, 12, 802222. [Google Scholar] [CrossRef] [PubMed]
- Moskal, K.; Kowalik, S.; Podyma, W.; Łapi´nski, B.; Boczkowska, M. The pros and cons of rye chromatin introgression into wheat genome. Agronomy 2021, 11, 456. [Google Scholar] [CrossRef]
- Henikoff, S.; Ahmad, K.; Malik, H.S. The centromere paradox: Stable inheritance with rapidly evolving DNA. Science 2001, 293, 1098–1102. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.C.; Torres, G.A. Plant centromeres: Genetics, epigenetics and evolution. Mol. Biol. Rep. 2018, 45, 1491–1497. [Google Scholar] [CrossRef]
- Su, H.; Liu, Y.; Liu, C.; Shi, Q.; Huang, Y.; Han, F. Centromere satellite repeats have undergone rapid changes in polyploid wheat subgenomes. Plant Cell 2019, 31, 2035–2051. [Google Scholar] [CrossRef]
- Gong, Z.; Wu, Y.; Koblízková, A.; Torres, G.A.; Wang, K.; Iovene, M.; Neumann, P.; Zhang, W.; Novák, P.; Buell, C.R.; et al. Repeatless and repeat-based centromeres in potato: Implications for centromere evolution. Plant Cell 2012, 24, 3559–3574. [Google Scholar] [CrossRef]
- Lukaszewski, A.J. Behavior of centromeres in univalents and centric misdivision in wheat. Cytogenet. Genome Res. 2010, 129, 97–109. [Google Scholar]
- Kopecky, D.; Lukaszewski, A.J. Misdivision of telocentrics and isochromosomes in wheat. Cytogenet. Genome Res. 2019, 157, 179–188. [Google Scholar]
- Tang, H.J.; Yin, G.H.; Xia, X.C.; Feng, J.J.; Qu, Y.Y.; He, Z.H. Evalution of molecular markers specific for 1BL.1RS translocation and characterization of 1RS chromosome in wheat varieties from different origins. Acta Agron. Sin. 2009, 35, 2107–2115. [Google Scholar] [CrossRef]
- Ren, T.H.; Yang, Z.J.; Yan, B.J.; Zhang, H.Q.; Fu, S.L.; Ren, Z.L. Development and characterization of a new 1BL.1RS translocation line with resistance to stripe rust and powdery mildew of wheat. Euphytica 2009, 169, 207–213. [Google Scholar] [CrossRef]
- Jia, J.; Xie, Y.; Cheng, J.; Kong, C.; Wang, M.; Gao, L.; Zhao, F.; Guo, J.; Wang, K.; Li, G.; et al. Homology-mediated inter-chromosomal interactions in hexaploid wheat lead to specific subgenome territories following polyploidization and introgression. Genome Biol. 2021, 22, 26. [Google Scholar] [CrossRef]
- Fu, S.L.; Yang, M.Y.; Ren, Z.L.; Yan, B.J.; Tang, Z.X. Abnormal mitosis induced by wheat-rye 1R monosomic addition lines. Genome 2014, 57, 21–28. [Google Scholar] [CrossRef]
- Aragón-Alcaide, L.; Miller, T.; Schwarzacher, T.; Reader, S.; Moore, G. A cereal centromeric sequence. Chromosoma 1996, 105, 261–268. [Google Scholar] [CrossRef]
- Li, G.; Wang, L.; Yang, J.; He, H.; Jin, H.; Li, X.; Ren, T.; Ren, Z.; Li, F.; Han, X.; et al. A high-quality genome assembly highlights rye genomic characteristics and agronomically important genes. Nature Genet. 2021, 53, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Wan, L.; Luo, J.; Tang, Z.; Fu, S. FISH landmarks reflecting meiotic recombination and structural alterations of chromosomes in wheat (Triticum aestivum L.). BMC Plant Biol. 2021, 21, 167. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Li, G.; Wang, H.; Yu, Z.; Chen, Q.; Yang, E.; Fu, S.; Tang, Z.; Yang, Z. Physical location of tandem repeats in the wheat genome and application for chromosome identification. Planta 2019, 249, 663–675. [Google Scholar] [CrossRef]
- Han, F.P.; Lamb, J.C.; Birchler, A. High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize. Proc. Natl. Acad. Sci. USA 2006, 103, 3238–3243. [Google Scholar] [CrossRef]
- Tang, Z.; Yang, Z.; Fu, S. Oligonucleotides replacing the roles of repetitive sequences pAs1, pSc119.2, pTa-535, pTa71, CCS1, and pAWRC.1 for FISH analysis. J. Appl. Genet. 2014, 55, 313–318. [Google Scholar] [CrossRef]
- Fu, S.; Chen, L.; Wang, Y.; Li, M.; Yang, Z.; Qiu, L.; Yan, B.; Ren, Z.; Tang, Z. Oligonucleotide probes for ND-FISH analysis to identify rye and wheat chromosomes. Sci. Rep. 2015, 5, 10552. [Google Scholar] [CrossRef]
- Xiao, Z.; Tang, S.; Qiu, L.; Tang, Z.; Fu, S. Oligonucleotides and ND-FISH displaying different arrangements of tandem repeats and identification of Dasypyrum villosum chromosomes in wheat backgrounds. Molecules 2017, 22, 973. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Primer | Sequence of Primer (5′-3′) | Target Region of Amplification (bp) * |
|---|---|---|
| Primer-1R1 | F: GGGTGATTGGCATATCTCGT R: GCAGCCACAGGTATCTCGTT | 1R: 349,498,361 to 349,501,266 |
| Primer-3R1 | F: ACATTGGGTCTCTGCGTCAG R: AGAACATCTCAATTCGTGGTGC | 3R: 517,650,456 to 517,652,830 |
| Primer-4R1 | F: CAGACAGGCGAGCAGGATAG R: CTCTTCCGCAACAGCTACCT | 4R: 409,233,133 to 409,235,918 |
| Primer-4R2 | F: GCTCGCTCTCTTACCCTTCC R: TGTTCTCACAGGCTTTTTCTCA | 4R: 414,933,891 to 414,936,603 |
| Primer-5R1 | F: CTAGGCAGCTGGGTAATGCG R: CTCCTCTCCCCTAACCCTCC | 5R: 237,421,693 to 237,423,703 |
| Primer-5R2 | F: CGCGATCCCCATCTCTGTTT R: GTCGCCTCACCTTACGCTTT | 5R: 245,442,755 to 245,445,320 |
| Primer-6R2 | F: TGCCCAGACCAGCTAGACTA R: TACAACAGCAACCCGAGCAA | 6R: 309,045,272 to 309,048,228 |
| Primer-6R3 | F: GTCTTGCAAATTGGTTCATGAGA R: CACTACAAGATACCAACTCCA | 6R: 322,669,622 to 322,671,571 |
| Primer-7R1 | F: AGTGAAGTTCCCGTTGGTCA R: TCCAGCTGTTGAACCATCCA | 7R: 468,542,502 to 468,545,143 |
| Name of Single-Copy Sequences | Length of Sequence (bp) | Source of Sequence | Location on Chromosome | GenBank Accession Number | Similarity with Its Corresponding Sequence in Weining (%) |
|---|---|---|---|---|---|
| 1R1 | 2906 | Kustro | centromere of 1R | ON557265 | 100.00 |
| SL-1R1 | 2900 | Secale sylvestre | ON557267 | 99.21 | |
| SD-1R1 | 2914 | Secale strictum | ON557268 | 99.86 | |
| 3R1 | 2373 | Kustro | centromere of 3R | ON557271 | 99.92 |
| SL-3R1 | 2382 | Secale sylvestre | ON557272 | 99.16 | |
| SD-3R1 | 2374 | Secale strictum | ON557273 | 99.96 | |
| 4R1 | 2787 | Kustro | centromere of 4R | ON557274 | 99.86 |
| SD-4R1 | 2786 | Secale strictum | ON557275 | 99.93 | |
| 4R2 | 2713 | Kustro | ON557276 | 99.78 | |
| 5R1 | 2011 | Kustro | centromere of 5R | ON557277 | 99.65 |
| 5R2 | 2566 | Kustro | ON557278 | 99.84 | |
| 6R2 | 2957 | Kustro | centromere of 6R | ON557279 | 99.80 |
| 6R3 | 1950 | Kustro | ON557280 | 99.69 | |
| 7R1 | 2642 | Kustro | centromere of 7R | ON557281 | 99.85 |
| SD-7R1 | 2642 | Secale strictum | ON557282 | 99.89 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, Z.; Luo, J.; Tang, Z.; Fu, S. Centromere-Specific Single-Copy Sequences of Secale Species. Plants 2022, 11, 2117. https://doi.org/10.3390/plants11162117
Pan Z, Luo J, Tang Z, Fu S. Centromere-Specific Single-Copy Sequences of Secale Species. Plants. 2022; 11(16):2117. https://doi.org/10.3390/plants11162117
Chicago/Turabian StylePan, Zijin, Jie Luo, Zongxiang Tang, and Shulan Fu. 2022. "Centromere-Specific Single-Copy Sequences of Secale Species" Plants 11, no. 16: 2117. https://doi.org/10.3390/plants11162117