
Effects of Coir-Based Growing Medium with Municipal Solid Waste Compost or Biochar on Plant Growth, Mineral Nutrition, and Accumulation of Phytochemicals in Spinach

 , ,
, ,
Abstract
:1. Introduction
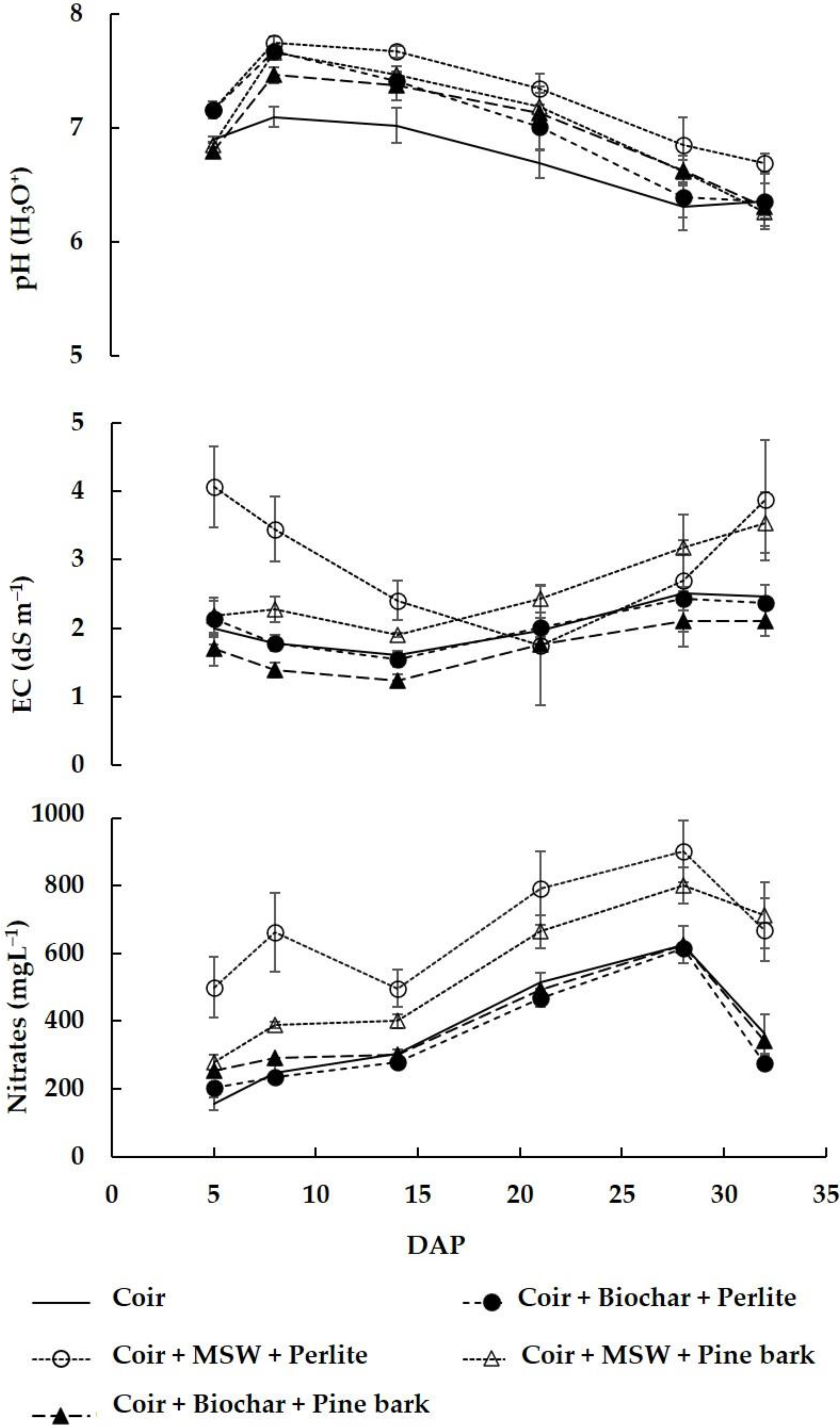
2. Results and Discussion
2.1. Initial Physical and Chemical Properties of the Media
2.2. Drainage Water
2.3. Shoot Nutrient Concentration and Uptake
2.4. Photosynthetic Pigments
2.5. Plant Growth
2.6. Phytochemical Accumulation and Antioxidant Activity
2.6.1. Total Phenols and Antioxidant Activities
2.6.2. Ascorbate-Glutathione Cycle
3. Conclusions
4. Material and Methods
4.1. Growth Conditions and Substrates
4.2. Measurements
4.3. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gruda, N.S.; Bragg, N. Developments in alternative organic materials for growing media in soilless culture systems. In Advances in Horticultural Soilless Culture; Burleigh Dodds Science Publishing: Cambridge, UK, 2021; pp. 73–106. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Alves-Pereira, I.; Ferreira, R.; Gruda, N.S. Coir, an Alternative to Peat—Effects on Plant Growth, Phyto-chemical Accumulation, and Antioxidant Power of Spinach. Horticulturae 2021, 7, 127. [Google Scholar] [CrossRef]
- Barcelos, C.; Machado, R.M.; Alves Pereira, I.; Ferreira, R.; Bryla, D.R. Effects of substrate type on plant growth and nitrogen and nitrate concentration in spinach. Int. J. Plant Biol. 2016, 7, 44–47. [Google Scholar] [CrossRef] [Green Version]
- Michel, J.C. The physical properties of peat: A key factor for modern growing media. Mires Peat 2010, 6. Available online: https://hal-agrocampus-ouest.archives-ouvertes.fr/hal-00729716 (accessed on 8 May 2022).
- Fascella, G. Growing substrates alternative to peat for ornamental plants. In Soilless Culture-Use of Substrates for the Production of Quality Horticultural Crops; Asaduzzaman, M., Ed.; IntechOpen: London, UK, 2015; pp. 47–67. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N.; Caron, J.; Prasad, M.; Maher, M. Growing media. Encyclopedia of Soil Science; CRC Press: Boca Raton, FL, USA, 2016; pp. 1053–1058. [Google Scholar]
- Sani, M.N.H.; Yong, J.W. Harnessing Synergistic Biostimulatory Processes: A Plausible Approach for Enhanced Crop Growth and Resilience in Organic Farming. Biology 2021, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, S.F.; Kenar, J.A.; Thompson, A.R.; Peterson, S.C. Comparison of biochars derived from wood pellets and pelletized wheat straw as replacements for peat in potting substrates. Ind. Crops Prod. 2013, 51, 437–443. [Google Scholar] [CrossRef]
- Graber, E.R.; Harel, Y.M.; Kolton, M.; Cytryn, E.; Silber, A.; David, D.R.; Tsechansky, L.; Borenshtein, M.; Elad, Y. Biochar impact on development and productivity of pepper and tomato grown in fertigated soilless media. Plant Soil 2010, 337, 481–496. [Google Scholar] [CrossRef]
- Gaskin, J.W.; Steiner, C.; Harris, K.; Das, K.C.; Bibens, B. Effect of low-temperature pyrolysis conditions on biochar for agricultural use. Trans. ASABE 2008, 51, 2061–2069. Available online: https://www.researchgate.net/publication/237079730 (accessed on 4 March 2022).
- Dispenza, V.; De Pasquale, C.; Fascella, G.; Mammano, M.M.; Alonzo, G. Use of biochar as peat substitute for growing substrates of Euphorbia × lomi potted plants. Span. J. Agric. Res. 2016, 14, e0908. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.M.A.; Bui, H.H. Physicochemical characteristics of substrates of the mixture of coir with municipal solid waste compost and with biochar. VIII South-East. Eur. Symp. Veg. Potatoes 2021, 1320, 399–404. [Google Scholar] [CrossRef]
- Altland, J.E.; Jeong, K.Y. Dolomitic lime amendment affects pine bark substrate pH, nutrient availability, and plant growth: A review. HortTechnology 2016, 26, 565–573. [Google Scholar] [CrossRef]
- Bilderback, T.E.; Warren, S.L.; Owen, J.S., Jr.; Albano, J.P. Healthy substrates need physicals too! HortTechnology 2005, 15, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Martínez, P.F.; Roca, D. Sustratos para el cultivo sin suelo. Materiales, propiedades y manejo. In Sustratos, Manejo del Clima, Automatización y Control en Sistemas de Cultivo sin Suelo; Flórez, R.V.J., Ed.; Editorial Universidad Nacional de Colombia: Bogotá, Colombia, 2011; pp. 37–77. [Google Scholar]
- Lemaire, F.; Dartigues, A.; Riviere, L.M. Properties of substrates with ground pine bark. Acta Hortic. 1980, 99, 67–80. [Google Scholar] [CrossRef]
- Fisher, P.R.; Dickson, R.W.; Mohammad-Pour, G.S.; Huang, J. Effect of solution electrical conductivity (EC) and pre-plant nutrient form on the pH of a peat-perlite substrate. In International Symposium on Growing Media and Soilless Cultivation; ISHS: Leuven, Belgium, 2013; Volume 1034, pp. 249–254. [Google Scholar] [CrossRef]
- Carlile, W.; Cattivello, C.; Zaccheo, P. Organic growing media: Constituents and properties. Vadose Zone J. 2015, 14, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.B. Growing Plants Hydroponically; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar] [CrossRef]
- Saleh, H.A.-R.; El-Nashar, Y.I.; Serag-El-Din, M.F.; Dewir, Y.H. Plant growth, yield and bioactive compounds of two culinary herbs as affected by substrate type. Sci. Hortic. 2019, 243, 464–471. [Google Scholar] [CrossRef]
- Nerlich, A.; Dannehl, D. Soilless cultivation: Dynamically changing chemical properties and physical conditions of organic substrates influence the plant phenotype of lettuce. Front. Plant Sci. 2021, 11, 601455. [Google Scholar] [CrossRef]
- Bailey, D.A.; Nelson, P.V.; Fonteno, W.C. Substrates pH and Water Quality; North Carolina State University: Raleigh, NC, USA, 2000. [Google Scholar]
- Baudoin, W.; Grafiadellis, M.; Jiménez, R.; La Malfa, G.; Marínez-García, P.; Nisen, A.; Garnaud, J. El Cultivo Protegido en Clima Mediterráneo: Estudio FAO Producción y Protección Vegetal; FAO: Rome, Italy, 2002; p. 90. [Google Scholar]
- Soria, C.B.; Oloverti, J.M. Cultivo sin suelo de hortalizas. Aspectos Prácticos y Experiencias. Serie de divulgación técnica; Generalitat Valenciana, Consellería de Agricultura, Pesca y Alimentación: Valencia, Spain, 2002; p. 110. [Google Scholar]
- Pardossi, A.; Carmassi, G.; Diara, C.; Incrocci, L.; Maggini, R.; Massa, D. Fertigation and Substrate Management in Closed Soilless Culture; University of Pisa: Pisa, Italy, 2011. [Google Scholar]
- Fonteno, W.C.; Harden, C.T. Procedures for determining physical properties of horticultural substrates using the NCS-Porometer. Horticultural Substrates Laboratory; North Carolina State University: Raleigh, NC, USA, 2003; Available online: https://www.ncsu.edu/project/hortsublab/pdf/porometer_manual.pdf (accessed on 21 March 2022).
- Lee, J.; Lee, B.; Kang, J.; Bae, J.; Ku, Y.; Gorinstein, S.; Lee, J. Effect of root zone aeration on the growth and bioactivity of cucumber plants cultured in perlite substrate. Biologia 2014, 69, 610–617. [Google Scholar] [CrossRef]
- Kingston, P.H.; Scagel, C.F.; Bryla, D.R.; Strik, B.C. Influence of perlite in peat-and coir-based media on vegetative growth and mineral nutrition of highbush blueberry. HortScience 2020, 55, 658–663. [Google Scholar] [CrossRef]
- Prasad, M.; Chrysargyris, A.; McDaniel, N.; Kavanagh, A.; Gruda, N.S.; Tzortzakis, N. Plant nutrient availability and pH of biochars and their fractions, with the possible use as a component in a growing media. Agronomy 2020, 10, 10. [Google Scholar] [CrossRef] [Green Version]
- Kulczycki, G.; Magnucka, E.G.; Oksińska, M.P.; Kucińska, J.; Kobyłecki, R.; Pawęska, K.; Zarzycki, R.; Kacprzak, A.; Pietr, S.J. The Effect of Various Types of Biochar Mixed with Mineral Fertilization on the Development and Ionome of Winter Wheat (Triticum aestivum L.) Seedlings and Soil Properties in a Pot Experiment. Agronomy 2020, 10, 1903. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Candan, N.; Tarhan, L. Effects of calcium, stress on contents of chlorophyll and carotenoid, LPO levels, and antioxidant enzyme activities in Mentha. J. Plant Nutr. 2005, 28, 127–139. [Google Scholar] [CrossRef]
- Borowski, E.; Michalek, S. The effect of foliar nutrition of spinach (Spinacia oleracea L.) with magnesium salts and urea on gas exange, leaf yield and quality. Acta Agrobot 2010, 63, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.M.A.; Alves-Pereira, I.; Lourenço, D.; Ferreira, R.M.A. Effect of organic compost and inorganic nitrogen fertigation on spinach growth, phytochemical accumulation, and antioxidant activity. Heliyon 2020, 6, e05085. [Google Scholar] [CrossRef]
- Schlering, C.; Zinkernagel, J.; Dietrich, H.; Frisch, M.; Schweiggert, R. Alterations in the chemical composition of spinach (Spinacia oleracea L.) as provoked by season and moderately limited water supply in open field cultivation. Horticulturae 2020, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Mou, B. Responses of spinach to salinity and nutrient deficiency in growth, physiology, and nutritional value. J. Am. Soc. Hortic. Sci. 2016, 141, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Sonneveld, C.; Baas, R.; Nijssen, H.M.C.; De Hoog, J. Salt tolerance of flower crops grown in soilless culture. J. Plant Nutr. 1999, 22, 1033–1048. [Google Scholar] [CrossRef]
- Ferreira, J.F.; Sandhu, D.; Liu, X.; Halvorson, J.J. Spinach (Spinacea oleracea L.) response to salinity: Nutritional value, physiological parameters, antioxidant capacity, and gene expression. Agriculture 2018, 8, 163. [Google Scholar] [CrossRef] [Green Version]
- Machado, R.M.A.; Serralheiro, R.P. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Casierra-Posada, F.; Ulrichs, C.; Pérez, C. Growth of spinach plants (Spinacia oleracea L.) exposed to excess zinc and manganese. Agron. Colomb. 2012, 30, 345–350. [Google Scholar]
- Proteggente, A.R.; Pannala, A.S.; Paganga, G.; van Buren, L.; Wagner, E.; Wiseman, S.; van de Put, F.; Dacombe, C.; Rice-Evans, C.A. The antioxidant activity of regularly consumed fruit and vegetables reflects their phenolic and vitamin C composition. Free Radic. Res. 2002, 36, 217–223. [Google Scholar] [CrossRef]
- Tavarini, S.; Cardelli, R.; Saviozzi, A.; Degl’Innocenti, E.; Guidi, L. Effects of green sompost on Soil biochemical characteristics and nutritive quality of leafy vegetables. Compost. Sci. Util. 2011, 19, 114–122. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Alves-Pereira, I.; Ferreira, R.M.A. Plant growth, phytochemical accumulation and antioxidant activity of substrate-grown spinach. Heliyon 2018, 4, e00751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galani, J.H.Y.; Patel, J.S.; Patel, N.J.; Talati, J.G. Storage of Fruits and Vegetables in Refrigerator Increases their Phenolic acids but decreases the total phenolics, anthocyanins and vitamin c with Subsequent Loss of their Antioxidant Capacity. Antioxidants 2017, 6, 59. [Google Scholar] [CrossRef] [Green Version]
- Apack, R.; Güçlü, K.; Demirata, B.; Özyürek, M.; Çelik, S.; Bektasoğlü, B.; Berker, K.; Özyurt, D. Comparative evaluation of various total antioxidant capacity assays applied to phenolic compounds with the CUPRAC assay. Molecules 2007, 12, 1496–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzucotelli, C.A.; Gonzalez-Aguilar, G.A.; Villegas-Ochoa, M.A.; Domínguez-Avila, A.J.; Ansorena, M.R.; Di Scala, K.C. Chemical characterization and functional properties of selected leafy vegetables for innovative mixed salads. J. Food Biochem. 2018, 42, e12461. [Google Scholar] [CrossRef] [Green Version]
- Stefanelli, D.; Winkler, S.; Jones, R. Reduced nitrogen availability during growth improves quality in red oak lettuce leaves by minimizing nitrate content and increasing antioxidant capacity and leaf mineral content. Agric. Sci. 2011, 2, 477–486. [Google Scholar] [CrossRef]
- Anjum, N.A.; Umar, S.; Chan, M.T. Ascorbate-Glutathione Pathway and Stress Tolerance in Plants; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Orsini, F.; Maggio, A.; Rouphael, Y.; De Pascale, S. Physiological Quality of organically grown vegetables. Sci. Hortic. 2016, 208, 131–139. [Google Scholar] [CrossRef]
- Tewari, R.K.; Kumar, P.; Tewari, N.; Srivastava, S.; Sharma, P.N. Macronutrient deficiencies and differential antioxidant responses—influence on the activity and expression of superoxide dismutase in maize. Plant Sci. 2004, 166, 687–694. [Google Scholar] [CrossRef]
- Cakmak, I. Activity of ascorbate-dependent H2O2-scavenging enzymes and leaf chlorosis are enhanced in magnesium- and potassium-deficient leaves, but not in phosphorus-deficient leaves. J. Exp. Botany 1994, 45, 1259–1266. Available online: https//www.jstor.org/stable/23694597 (accessed on 7 April 2022). [CrossRef]
- Hippler, F.W.R.; Boaretto, R.M.; Dovis, V.L.; Quaggio, J.A.; Azevedo, R.A.; Mattos, D., Jr. Oxidative stress induced by Cu nutritional disorders in Citrus depends on nitrogen and calcium availability. Sci. Rep. 2018, 8, 1641. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.K.; Yadav, N.; Gupta, R.; Kumar, P. Oxidative stress under macronutrient deficiency in plants. J. Soil Sci. Plant Nutri. 2021, 21, 832–859. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Cross-talk between calcium and reactive oxygen species originated from NADPH oxidase in abscisic acid-induced antioxidant defence in leaves of maize seedlings. Plant Cell Environ. 2003, 26, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Ryu, S.H.; Jang, I.C.; Kwon, S.Y.; Kim, J.G.; Kwak, S.S. Molecular cloning of a cytosolic ascorbate peroxidase cDNA from cell cultures of sweet potato and its expression in response to stress. Mol. Genet. Genom. 2004, 271, 339–346. [Google Scholar] [CrossRef]
- Ntagkas, N.; Woltering, E.J.; Marcelis, L.F.M. Light regulates ascorbate in plants: An integrated view on physiology and biochemistry. Environ. Exp. Bot. 2018, 147, 271–280. [Google Scholar] [CrossRef]
- Shin, R.; Berg, R.H.; Schachtman, D.P. Reactive oxygen species and root hairs in Arabidopsis root response to nitrogen, phosphorus and potassium deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef]
- Amlinger, F.; Favoino, E.; Pollak, M.; Peyr, S.; Centemero, M.; Caima, V. Heavy Metals and Organic Compounds from Wastes Used as Organic Fertilisers; Study on Behalf of the European Commission, Directorate-General Environment: Hochbergstr, Austria, 2004; pp. 168–210. [Google Scholar]
- Prazeres, A.O. Comparação de Metodologias Laboratoriais para Determinação de Azoto Nítrico e Amoniacal em Solos e Águas; Programa e Livro de Resumos do 1° Congresso Nacional e Rega e Drenagem: Beja, Portugal, 2005; pp. 59–60. [Google Scholar]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Lake, B. Preparation and characterization of microsomal fractions for studies of xenobiotic metabolism. In Biochemical Toxicology: A Practical Approach; Snell, K., Mullock, B., Eds.; IRL Press: Oxford, UK, 1987; pp. 183–215. [Google Scholar]
- Inoue, Y.; Matsuda, T.; Sugiyama, K.; Izawa, S.; Kimura, A. Genetic Analysis of glutathione peroxidase in oxidative stress response of Saccharomyces Cerevisiae. J. Biol. Chem. 1999, 17, 27002–27009. [Google Scholar] [CrossRef] [Green Version]
- Janda, T.; Szalai, G.; Tari, I.; Páldi, E. Hydroponic treatment with salicylic acid decreases the effects of chilling injury in maize (Zea mays L.) plants. Planta 1998, 208, 175–180. [Google Scholar] [CrossRef]


{kind=link}
| Substrate | Coir | Coir + Biochar + Perlite | Coir + MWS + Perlite | Coir + Biochar + Pine Bark | Coir + MSW + Pine Bark |
|---|---|---|---|---|---|
| pH * | 5.65 c† | 6.52 b | 7.10 a | 6.77 b | 7.00 a |
| EC (dS·m−1) * | 1.8 b | 1.51 c | 2.80 a | 1.12 c | 3.10 a |
| Total porosity (v/v. %) ** | 90.2 c | 97.0 a | 98.2 a | 95.7 b | 96.3 b |
| Moisture content (w/w, %) ** | 95.0 a | 81.1 b | 79.6 b | 78.9 b | 81.7 b |
| Mass wetness (g water/g substrate) ** | 7.90 a | 5.39 b | 5.01 b | 5.00 b | 4.94 b |
| Bulk density (g·cm−3) ** | 0.11 b | 0.17 a | 0.18 a | 0.17 a | 0.19 a |
| Substrate | Shoot Macronutrients (%) | Shoot Micronutrients (μg·g−1) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | Fe | B | Mn | Zn | Na 1 | |
| Coir | 5.87 a† | 0.56 a | 6.01 a | 0.51 d | 0.35 a | 141.3 a | 21.6 a | 85.6 b | 56.3 a | 0.84 a |
| Coir + biochar + perlite | 5.69 a | 0.54 a | 5.37 b | 0.72 bc | 0.35 a | 147.5 a | 26.4 a | 166.9 a | 60.0 a | 0.66 a |
| Coir + MWS + perlite | 4.68 b | 0.59 a | 5.67 ab | 1.07 a | 0.30 a | 160.6 a | 18.7 a | 61.87 c | 58.0 a | 0.91 a |
| Coir + biochar + pine bark | 5.75 a | 0.51 a | 5.28 b | 0.64 bc | 0.26 a | 138.7 a | 20.5 a | 148.1 a | 75.6 a | 0.80 a |
| Coir + MWS + pine bark | 5.40 a | 0.61 a | 5.70 ab | 0.84 b | 0.26 a | 133.8 a | 17.2 a | 60.6 c | 61.9 a | 0.88 a |
| Significance | * | NS | * | *** | NS | NS | NS | *** | NS | NS |
| Substrate | Shoot Macronutrients’ Uptake (mg/Plant) | Shoot Micronutrients’ Uptake (µg/Plant) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | Fe | B | Mn | Zn | Na 1 | |
| Coir | 135.0 a† | 2.91 b | 137.1 b | 11.5 c | 2.02 b | 326.6 b | 49.9 a | 196.4 c | 129.4 a | 1.9 a |
| Coir + biochar + perlite | 155.2 a | 4.15 ab | 146.9 ab | 19.8 b | 2.50 a | 400.9 ab | 71.2 a | 455.3 a | 163.9 a | 1.8 a |
| Coir + MWS + perlite | 143.8 a | 4.53 a | 173.5 a | 32.9 a | 2.88 a | 495.2 a | 57.4 a | 191.0 c | 179.8 a | 2.8 a |
| Coir + biochar + pine bark | 146.8 a | 2.22 c | 135.5 b | 16.4 bc | 1.70 c | 357.1 ab | 52.4 a | 379.6 b | 192.9 a | 2.0 a |
| Coir + MWS + pine bark | 151.3 a | 3.13 b | 159.5 ab | 23.7 b | 2.01 b | 377.2 ab | 48.2 a | 169.7 c | 171.8 a | 2.5 a |
| Significance | NS | *** | * | *** | ** | * | NS | *** | NS | NS |
| Substrate | Photosynthetic Pigments (mg/100 g FW) | ||||
|---|---|---|---|---|---|
| Total Chl | Chl a | Chl b | Cc | Chl a/Chl b Ratio | |
| Coir | 37.36 a† | 22.55 a | 14.81 ab | 54.59 ab | 1.51 a |
| Coir + biochar + perlite | 25.34 c | 12.70 bc | 12.63 b | 42.07 bc | 1.01 b |
| Coir + MSW + perlite | 31.44 b | 14.99 bc | 16.45 a | 54.76 a | 0.91 b |
| Coir + biochar + pine bark | 32.75 b | 17.24 ab | 15.51 ab | 48.07 abc | 1.14 b |
| Coir + MSW + pine bark | 24.03 c | 10.51 c | 13.52 b | 39.69 c | 0.78 b |
| Significance | ** | *** | * | * | *** |
| Substrate | Shoot Dry Weight | Leaf Area | Fresh Yield | |
|---|---|---|---|---|
| (g/Plant) | (%) | (cm2/Plant) | (kg/m2) | |
| Coir | 2.30 c† | 8.88 b | 510.9 a | 2.52 b |
| Coir + biochar + perlite | 2.73 ab | 9.50 a | 427.7 b | 2.75 b |
| Coir + MWS + perlite | 3.05 a | 9.05 a | 514.3 a | 3.25 a |
| Coir + biochar + pine bark | 2.56 bc | 9.40 a | 463.6 b | 2.61 b |
| Coir + MWS + pine bark | 2.86 ab | 9.39 a | 523.0 a | 2.87 ab |
| Significance | * | * | * | * |
| Substrate | TPC (mg GAE/100 g FW) | DPPH (mg GAE/100 g FW) | FRAP (mg Trolox/100 g FW) |
|---|---|---|---|
| Coir | 125.2 a | 67.0 a | 235.8 a |
| Coir + biochar + perlite | 122.9 a | 57.6 a | 161.2 c |
| Coir + MSW + perlite | 131.6 a | 53.6 a | 202.4 b |
| Coir + biochar + pine bark | 130.3 a | 64.6 a | 211.9 ab |
| Coir + MSW + pine bark | 102.4 a | 55.4 a | 172.6 c |
| Significance | NS | NS | *** |
| Substrate | APx a | GPx | GR | AsA | GSH |
|---|---|---|---|---|---|
| (nmol min−1/mg Protein) | (mg/100 g FW) | ||||
| Coir | 48.43 a | 59.6 b | 9.46 c | 10.77 b | 3.63 a |
| Coir + biochar + perlite | 24.79 b | 58.5 b | 17.67 a | 15.71 a | 2.76 b |
| Coir + MSW + perlite | 18.86 b | 73.8 b | 9.71 c | 14.41 a | 1.85 c |
| Coir + biochar + pine bark | 5.34 c | 107.6 a | 12.98 b | 14.38 a | 1.82 c |
| Coir + MSW + pine bark | 21.29 b | 68.8 b | 11.26 c | 9.06 b | 2.32 bc |
| Significance | *** | *** | *** | *** | *** |
| Substrate | Substrate Components (%, v/v) | ||||
|---|---|---|---|---|---|
| Coir | MSW | Biochar | Perlite | Pine Bark | |
| Coir | 100 | - | - | - | - |
| Coir + biochar + perlite | 78 | - | 12 | 10 | - |
| Coir + MSW + perlite | 78 | 12 | - | 10 | - |
| Coir + biochar + pine bark | 78 | - | 12 | - | 10 |
| Coir + MSW + pine bark | 78 | 12 | - | - | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado, R.M.A.; Alves-Pereira, I.; Morais, C.; Alemão, A.; Ferreira, R. Effects of Coir-Based Growing Medium with Municipal Solid Waste Compost or Biochar on Plant Growth, Mineral Nutrition, and Accumulation of Phytochemicals in Spinach. Plants 2022, 11, 1893. https://doi.org/10.3390/plants11141893
Machado RMA, Alves-Pereira I, Morais C, Alemão A, Ferreira R. Effects of Coir-Based Growing Medium with Municipal Solid Waste Compost or Biochar on Plant Growth, Mineral Nutrition, and Accumulation of Phytochemicals in Spinach. Plants. 2022; 11(14):1893. https://doi.org/10.3390/plants11141893
Chicago/Turabian StyleMachado, Rui M. A., Isabel Alves-Pereira, Carolina Morais, André Alemão, and Rui Ferreira. 2022. "Effects of Coir-Based Growing Medium with Municipal Solid Waste Compost or Biochar on Plant Growth, Mineral Nutrition, and Accumulation of Phytochemicals in Spinach" Plants 11, no. 14: 1893. https://doi.org/10.3390/plants11141893