
The Influence of Plant Stress Hormones and Biotic Elicitors on Cyclotide Production in Viola uliginosa Cell Suspension Cultures

 , and
, and 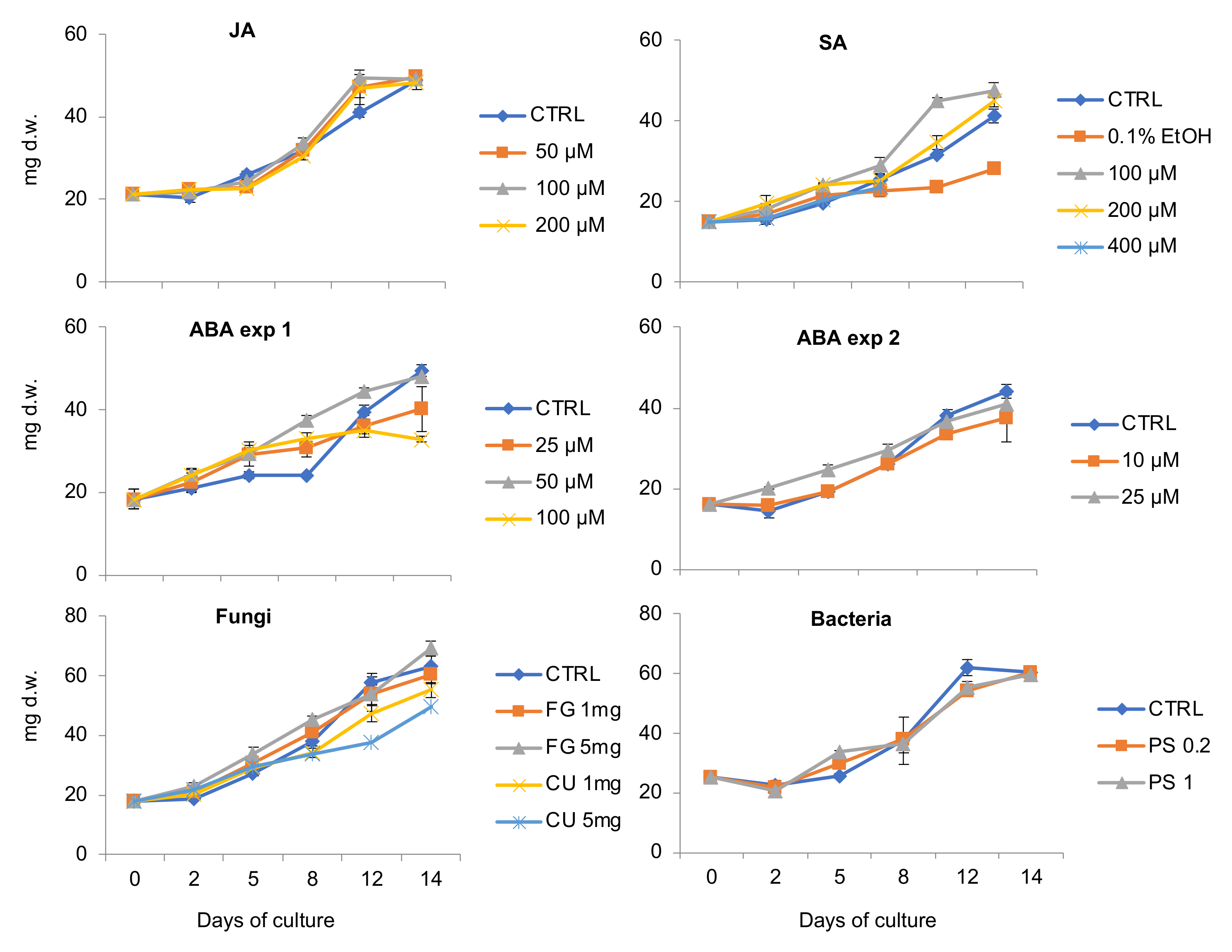{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Influence of Plant Stress Hormones and Elicitors on V. uliginosa Suspension Culture Growth
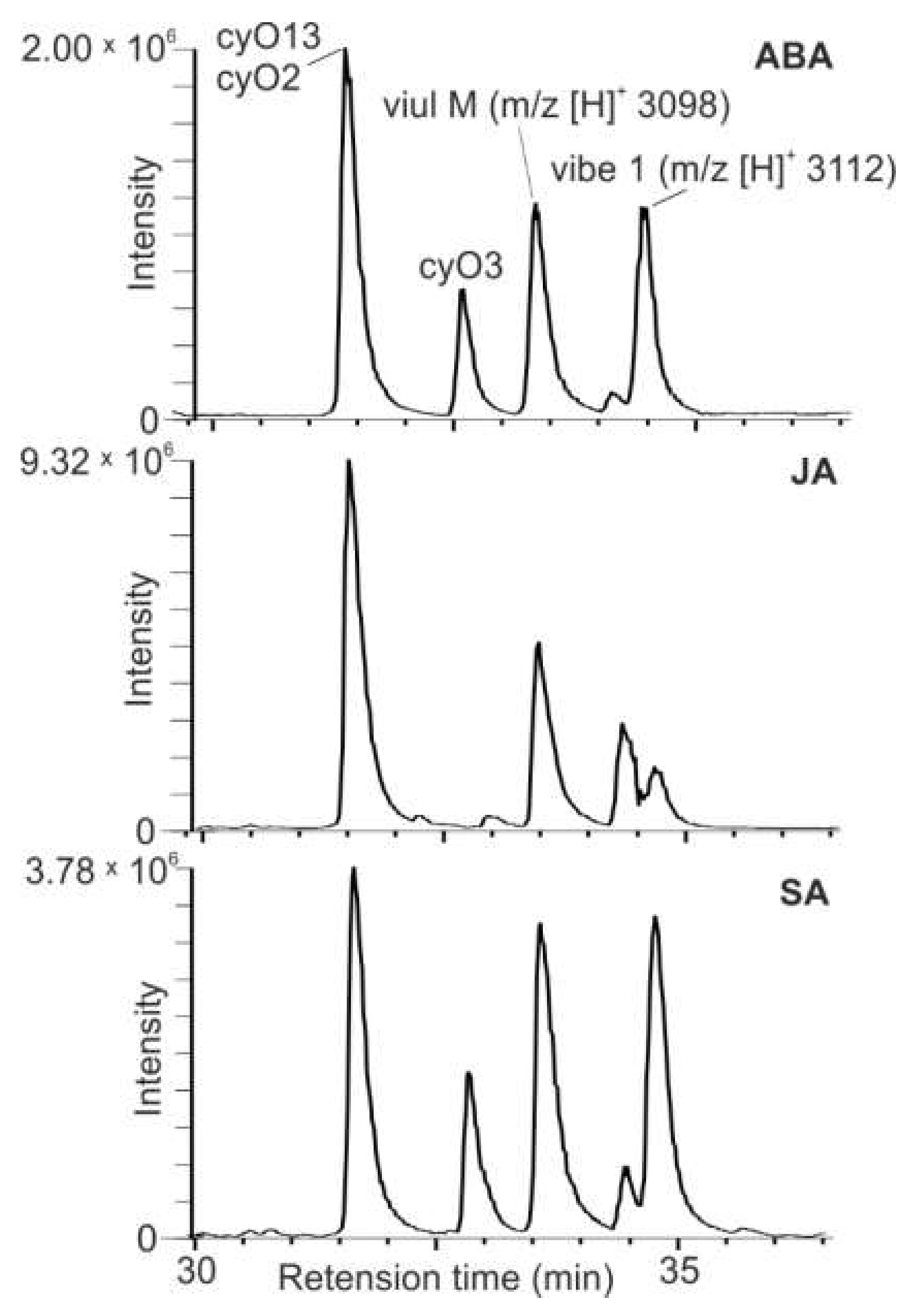
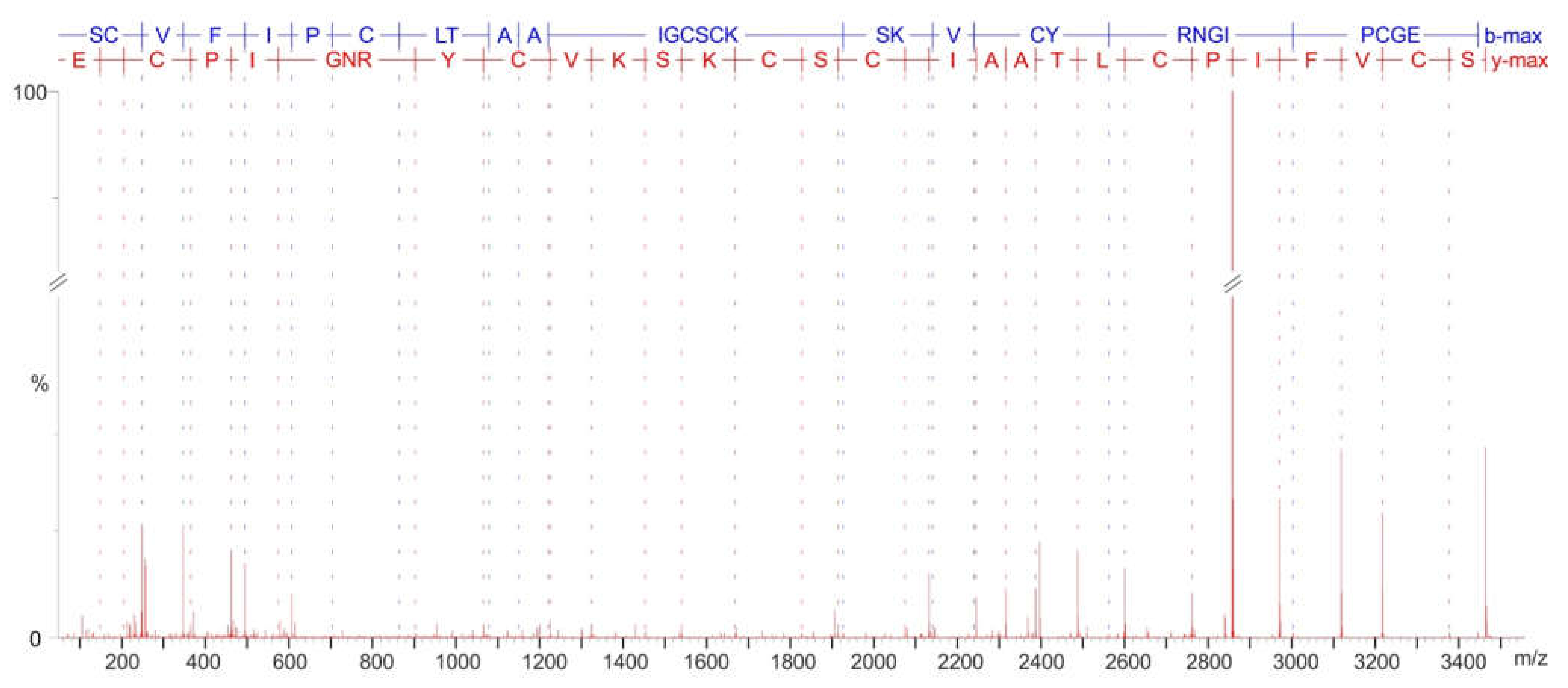
2.2. Major Cyclotides Produced by V. uliginosa Cell Suspensions and Discovery of a Novel Cyclotide
2.3. The Influence of Plant Stress Hormones and Elicitors on Relative Cyclotide Production in V. uliginosa Cell Cultures
3. Discussion
4. Materials and Methods
4.1. V. uliginosa Cell Suspension Initiation and Culture Conditions
4.2. Suspension Culture Treatments and Sample Preparation
4.3. Relative Quantitation of Cyclotides
4.4. Cyclotide MS/MS Sequencing
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poth, A.G.; Colgrave, M.L.; Philip, R.; Kerenga, B.; Daly, N.L.; Anderson, M.A.; Craik, D.J. Discovery of cyclotides in the Fabaceae plant family provides new insights into the cyclization, evolution, and distribution of circular proteins. ACS Chem. Biol. 2011, 6, 345–355. [Google Scholar] [CrossRef] [PubMed]
- Poth, A.G.; Mylne, J.S.; Grassl, J.; Lyons, R.E.; Millar, A.H.; Colgrave, M.L.; Craik, D.J. Cyclotides associate with leaf vasculature and are the products of a novel precursor in Petunia (Solanaceae). J. Biol. Chem. 2012, 287, 27033–27046. [Google Scholar] [CrossRef] [Green Version]
- Gran, L. On the Effect of a Polypeptide Isolated from “Kalata-Kalata” (Oldenlandia affinis DC) on the Oestrogen Dominated Uterus. Acta Pharmacol. Toxicol. 1973, 33, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Burman, R.; Yeshak, M.Y.; Larsson, S.; Craik, D.J.; Rosengren, K.J.; Göransson, U. Distribution of circular proteins in plants: Large-scale mapping of cyclotides in the Violaceae. Front. Plant Sci. 2015, 6, 855. [Google Scholar] [CrossRef] [Green Version]
- Hernandez, J.F.; Gagnon, J.; Chiche, L.; Nguyen, T.M.; Andrieu, J.P.; Heitz, A.; Hong, T.T.; Pham, T.T.C.; Le Nguyen, D. Squash trypsin inhibitors from Momordica cochinchinensis exhibit an atypical macrocyclic structure. Biochemistry 2000, 39, 5722–5730. [Google Scholar] [CrossRef]
- Nguyen, G.K.T.; Lian, Y.; Pang, E.W.H.; Nguyen, P.Q.T.; Tran, T.D.; Tam, J.P. Discovery of linear cyclotides in monocot plant Panicum laxum of Poaceae family provides new insights into evolution and distribution of cyclotides in plants. J. Biol. Chem. 2013, 288, 3370–3380. [Google Scholar] [CrossRef] [Green Version]
- Göransson, U.; Luijendijk, T.; Johansson, S.; Bohlin, L.; Claeson, P. Seven novel macrocyclic polypeptides from Viola arvensis. J. Nat. Prod. 1999, 62, 283–286. [Google Scholar] [CrossRef]
- Craik, D.J.; Daly, N.L.; Bond, T.; Waine, C. Plant cyclotides: A unique family of cyclic and knotted proteins that defines the cyclic cystine knot structural motif. J. Mol. Biol. 1999, 294, 1327–1336. [Google Scholar] [CrossRef]
- De Veer, S.J.; Kan, M.-W.; Craik, D.J. Cyclotides: From Structure to Function. Chem. Rev. 2019, 119, 12375–12421. [Google Scholar] [CrossRef] [PubMed]
- Burman, R.; Gunasekera, S.; Strömstedt, A.A.; Göransson, U. Chemistry and biology of cyclotides: Circular plant peptides outside the box. J. Nat. Prod. 2014, 77, 724–736. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, J.; Craik, D.J. Discovery, structure, function, and applications of cyclotides: Circular proteins from plants. J. Exp. Bot. 2016, 67, 4801–4812. [Google Scholar] [CrossRef] [Green Version]
- Jennings, C.; West, J.; Waine, C.; Craik, D.; Anderson, M.A. Biosynthesis and insecticidal properties of plant cyclotides: The cyclic knotted proteins from Oldenlandia affinis. Proc. Natl. Acad. Sci. USA 2001, 98, 10614–10619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pränting, M.; Lööv, C.; Burman, R.; Göransson, U.; Andersson, D.I. The cyclotide cycloviolacin O2 from Viola odorata has potent bactericidal activity against Gram-negative bacteria. J. Antimicrob. Chemother. 2010, 65, 1964–1971. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.K.L.; Colgrave, M.L.; Gustafson, K.R.; Ireland, D.C.; Goransson, U.; Craik, D.J. Anti-HIV cyclotides from the Chinese medicinal herb Viola yedoensis. J. Nat. Prod. 2008, 71, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Svangård, E.; Burman, R.; Gunasekera, S.; Lövborg, H.; Gullbo, J.; Göransson, U. Mechanism of action of cytotoxic cyclotides: Cycloviolacin O2 disrupts lipid membranes. J. Nat. Prod. 2007, 70, 643–647. [Google Scholar] [CrossRef]
- Dörnenburg, H. Cyclotide synthesis and supply: From plant to bioprocess. Biopolymers 2010, 94, 602–610. [Google Scholar] [CrossRef]
- Craik, D.J.; Conibear, A.C. The Chemistry of Cyclotides. J. Org. Chem. 2011, 76, 4805–4817. [Google Scholar] [CrossRef]
- Narayani, M.; Sai Varsha, M.K.N.; Potunuru, U.R.; Sofi Beaula, W.; Rayala, S.K.; Dixit, M.; Chadha, A.; Srivastava, S. Production of bioactive cyclotides in somatic embryos of Viola odorata. Phytochemistry 2018, 156, 135–141. [Google Scholar] [CrossRef]
- Slazak, B.; Jacobsson, E.; Kuta, E.; Göransson, U. Exogenous plant hormones and cyclotide expression in Viola uliginosa (Violaceae). Phytochemistry 2015, 117, 527–536. [Google Scholar] [CrossRef]
- Seydel, P.; Dörnenburg, H. Establishment of in vitro plants, cell and tissue cultures from Oldenlandia affinis for the production of cyclic peptides. Plant Cell. Tissue Organ Cult. 2006, 85, 247–255. [Google Scholar] [CrossRef]
- Dömenburg, H. Progress in kalata peptide production via plant cell bioprocessing. Biotechnol. J. 2009, 4, 632–645. [Google Scholar] [CrossRef]
- Slazak, B.; Kapusta, M.; Strömstedt, A.A.; Słomka, A.; Krychowiak, M.; Shariatgorji, M.; Andrén, P.E.; Bohdanowicz, J.; Kuta, E.; Göransson, U. How Does the Sweet Violet (Viola odorata L.) Fight Pathogens and Pests—Cyclotides as a Comprehensive Plant Host Defense System. Front. Plant Sci. 2018, 9, 1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylne, J.S.; Wang, C.K.; van der Weerden, N.L.; Craik, D.J. Cyclotides are a component of the innate defense of Oldenlandia affinis. Biopolymers 2010, 94, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Slazak, B.; Kaltenböck, K.; Steffen, K.; Rogala, M.; Rodríguez-Rodríguez, P.; Nilsson, A.; Shariatgorji, R.; Andrén, P.E.; Göransson, U. Cyclotide host-defense tailored for species and environments in violets from the Canary Islands. Sci. Rep. 2021, 11, 12452. [Google Scholar] [CrossRef]
- Simonsen, S.M.; Sando, L.; Ireland, D.C.; Colgrave, M.L.; Bharathi, R.; Göransson, U.; Craik, D.J. A continent of plant defense peptide diversity: Cyclotides in Australian Hybanthus (Violaceae). Plant Cell 2005, 17, 3176–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trabi, M.; Svangård, E.; Herrmann, A.; Göransson, U.; Claeson, P.; Craik, D.J.; Bohlin, L. Variations in cyclotide expression in Viola species. J. Nat. Prod. 2004, 67, 806–810. [Google Scholar] [CrossRef]
- Slazak, B.; Jędrzejska, A.; Badyra, B.; Sybilska, A.; Lewandowski, M.; Kozak, M.; Kapusta, M.; Shariatgorji, R.; Nilsson, A.; Andrén, P.E.; et al. The involvement of cyclotides in mutual interactions of violets and the two-spotted spider mite. Sci. Rep. 2022, 12, 1914. [Google Scholar] [CrossRef]
- Seydel, P.; Gruber, C.W.; Craik, D.J.; Dörnenburg, H. Formation of cyclotides and variations in cyclotide expression in Oldenlandia affinis suspension cultures. Appl. Microbiol. Biotechnol. 2007, 77, 275–284. [Google Scholar] [CrossRef]
- Narayani, M.; Chadha, A.; Srivastava, S. Callus and cell suspension culture of Viola odorata as in vitro production platforms of known and novel cyclotides. Plant Cell Tissue Organ Cult. 2017, 130, 289–299. [Google Scholar] [CrossRef]
- Slazak, B.; Haugmo, T.; Badyra, B.; Göransson, U. The life cycle of cyclotides: Biosynthesis and turnover in plant cells. Plant Cell Rep. 2020, 39, 1359–1367. [Google Scholar] [CrossRef]
- Rajendran, S.; Slazak, B.; Mohotti, S.; Strömstedt, A.A.; Göransson, U.; Hettiarachchi, C.M.; Gunasekera, S. Tropical vibes from Sri Lanka—Cyclotides from Viola betonicifolia by transcriptome and mass spectrometry analysis. Phytochemistry 2021, 187, 112749. [Google Scholar] [CrossRef] [PubMed]
- Gran, L.; Sandberg, F.; Sletten, K. Oldenlandia affinis (R and S) DC: A plant containing uteroactive peptides used in African traditional medicine. J. Ethnopharmacol. 2000, 70, 197–203. [Google Scholar] [CrossRef]
- Ireland, D.C.; Colgrave, M.L.; Craik, D.J. A novel suite of cyclotides from Viola odorata: Sequence variation and the implications for structure, function and stability. Biochem. J. 2006, 400, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.K.L.; Kaas, Q.; Chiche, L.; Craik, D.J. CyBase: A database of cyclic protein sequences and structures, with applications in protein discovery and engineering. Nucleic Acids Res. 2008, 36, 206–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Wang, J.; Qian, J.; Yao, L.; Lu, Y. Enhanced production of flavonoids by methyl jasmonate elicitation in cell suspension culture of Hypericum perforatum. Bioresour. Bioprocess. 2015, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [Green Version]
- Mylne, J.S.; Chan, L.Y.; Chanson, A.H.; Daly, N.L.; Schaefer, H.; Bailey, T.L.; Nguyencong, P.; Cascales, L.; Craik, D.J. Cyclic Peptides Arising by Evolutionary Parallelism via Asparaginyl-Endopeptidase-Mediated Biosynthesis. Plant Cell 2012, 24, 2765–2778. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Källback, P.; Nilsson, A.; Shariatgorji, M.; Andrén, P.E. msIQuant—Quantitation Software for Mass Spectrometry Imaging Enabling Fast Access, Visualization, and Analysis of Large Data Sets. Anal. Chem. 2016, 88, 4346–4353. [Google Scholar] [CrossRef]
- Broussalis, A.M.; Göransson, U.; Coussio, J.D.; Ferraro, G.; Martino, V.; Claeson, P. First cyclotide from Hybanthus (Violaceae). Phytochemistry 2001, 58, 47–51. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slazak, B.; Jędrzejska, A.; Badyra, B.; Shariatgorji, R.; Nilsson, A.; Andrén, P.E.; Göransson, U. The Influence of Plant Stress Hormones and Biotic Elicitors on Cyclotide Production in Viola uliginosa Cell Suspension Cultures. Plants 2022, 11, 1876. https://doi.org/10.3390/plants11141876
Slazak B, Jędrzejska A, Badyra B, Shariatgorji R, Nilsson A, Andrén PE, Göransson U. The Influence of Plant Stress Hormones and Biotic Elicitors on Cyclotide Production in Viola uliginosa Cell Suspension Cultures. Plants. 2022; 11(14):1876. https://doi.org/10.3390/plants11141876
Chicago/Turabian StyleSlazak, Blazej, Aleksandra Jędrzejska, Bogna Badyra, Reza Shariatgorji, Anna Nilsson, Per E. Andrén, and Ulf Göransson. 2022. "The Influence of Plant Stress Hormones and Biotic Elicitors on Cyclotide Production in Viola uliginosa Cell Suspension Cultures" Plants 11, no. 14: 1876. https://doi.org/10.3390/plants11141876