
Hirsutanone Isolated from the Bark of Alnus japonica Attenuates Melanogenesis via Dual Inhibition of Tyrosinase Activity and Expression of Melanogenic Proteins


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
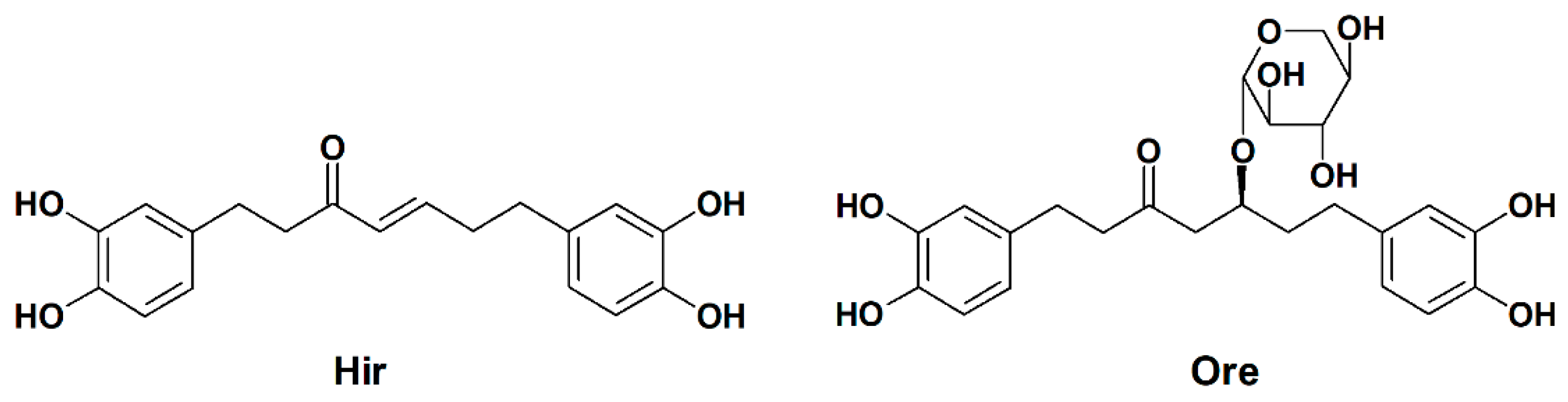
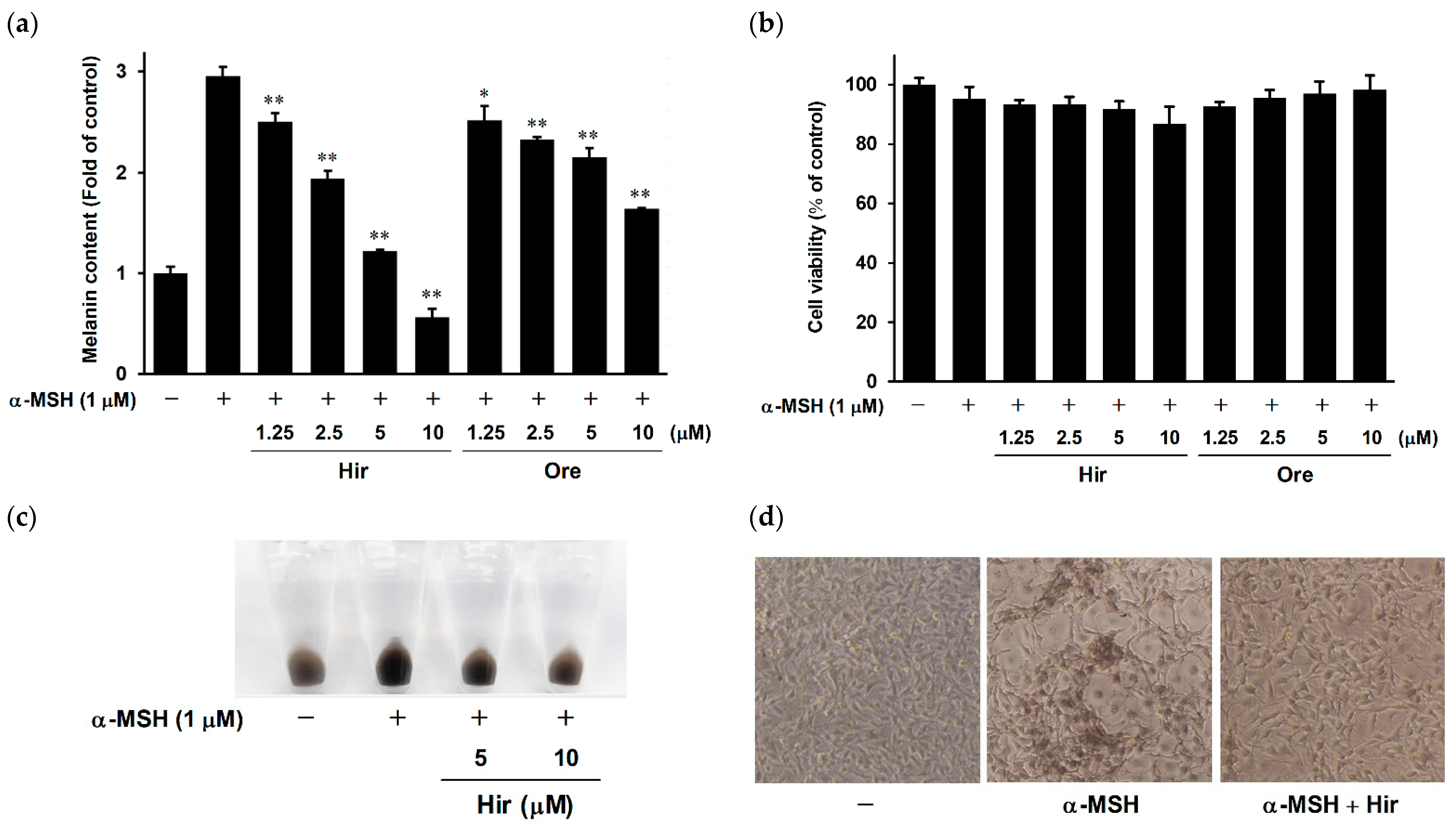
2.1. Effects of Hir and Ore on Melanin Biosynthesis and Cell Viability in B16-F1 Cells
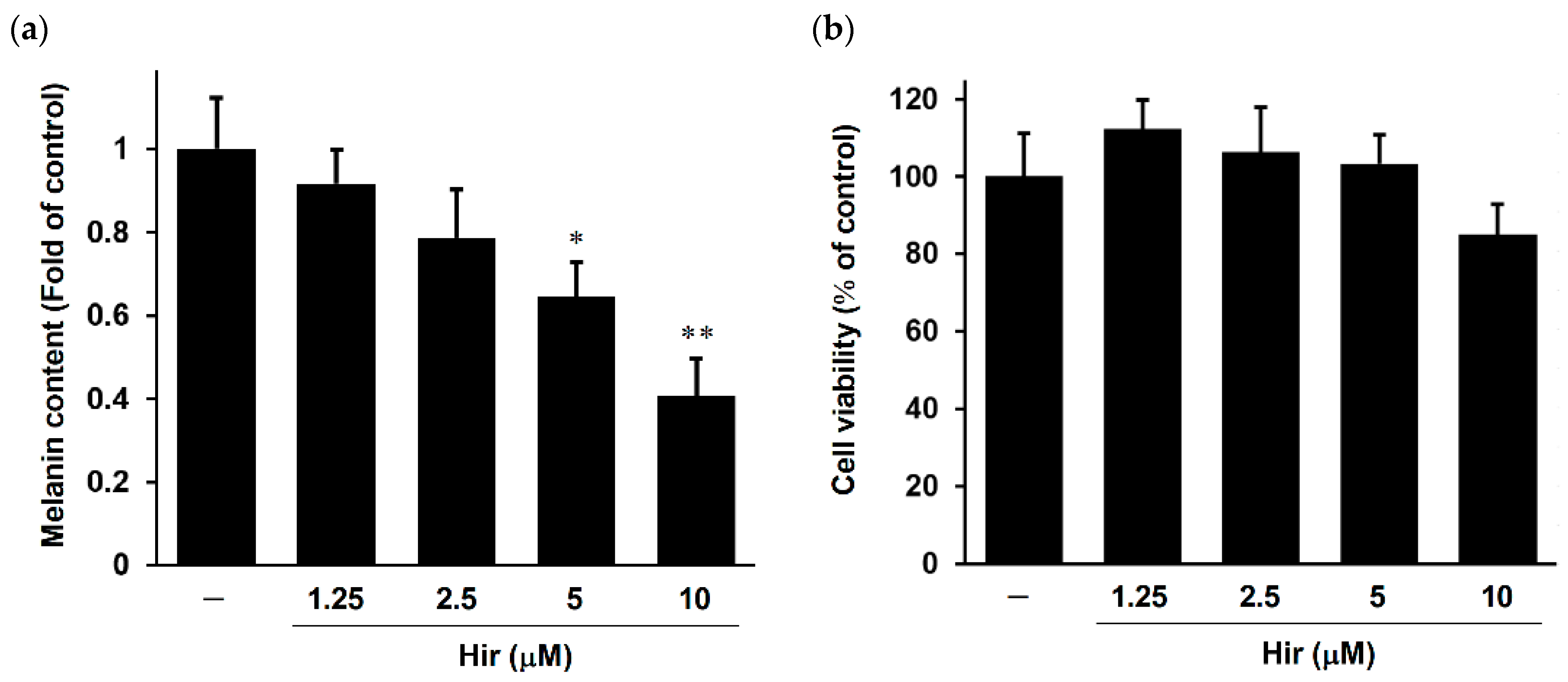
2.2. Effect of Hir on Melanin Biosynthesis and Cell Viability in Human Epidermal Melanocytes
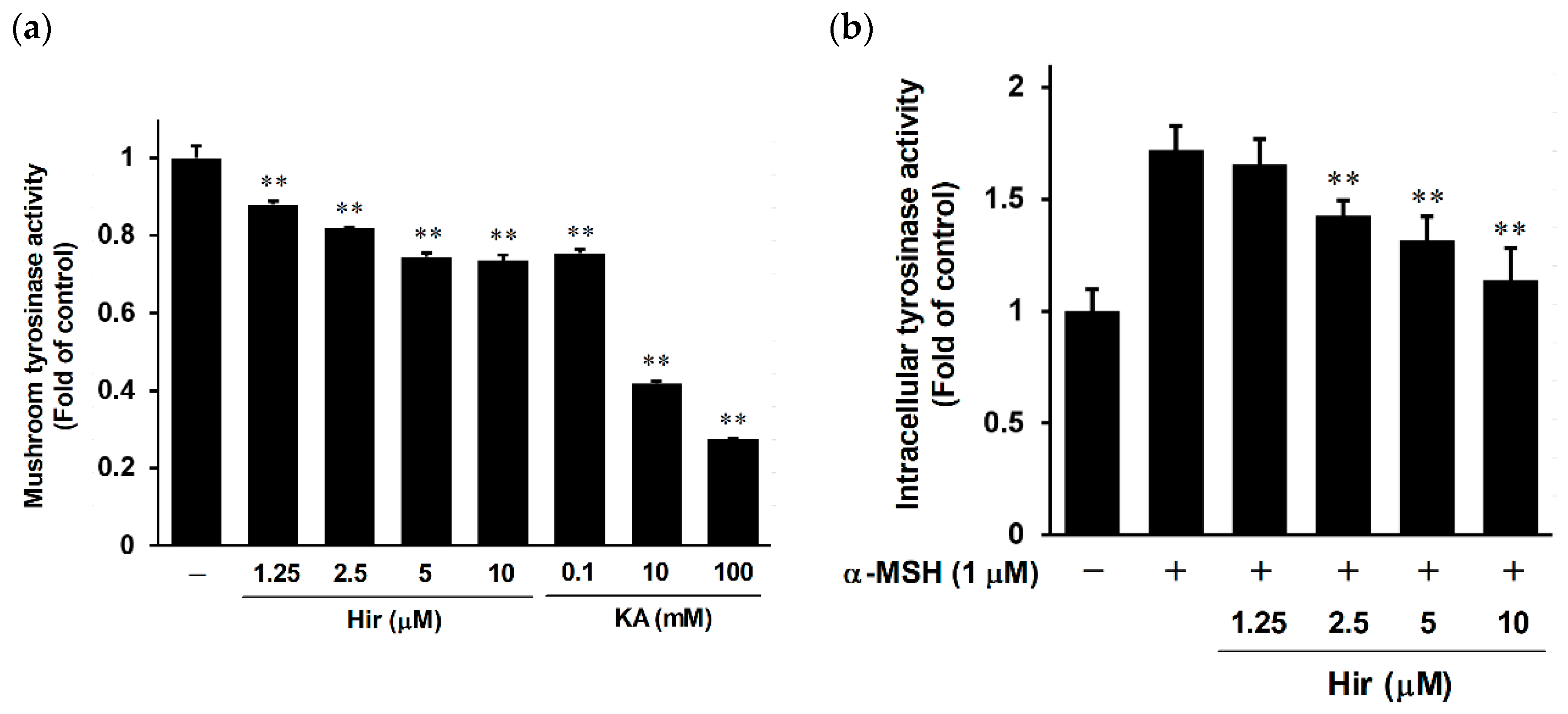
2.3. Effect of Hir on Tyrosinase Activity
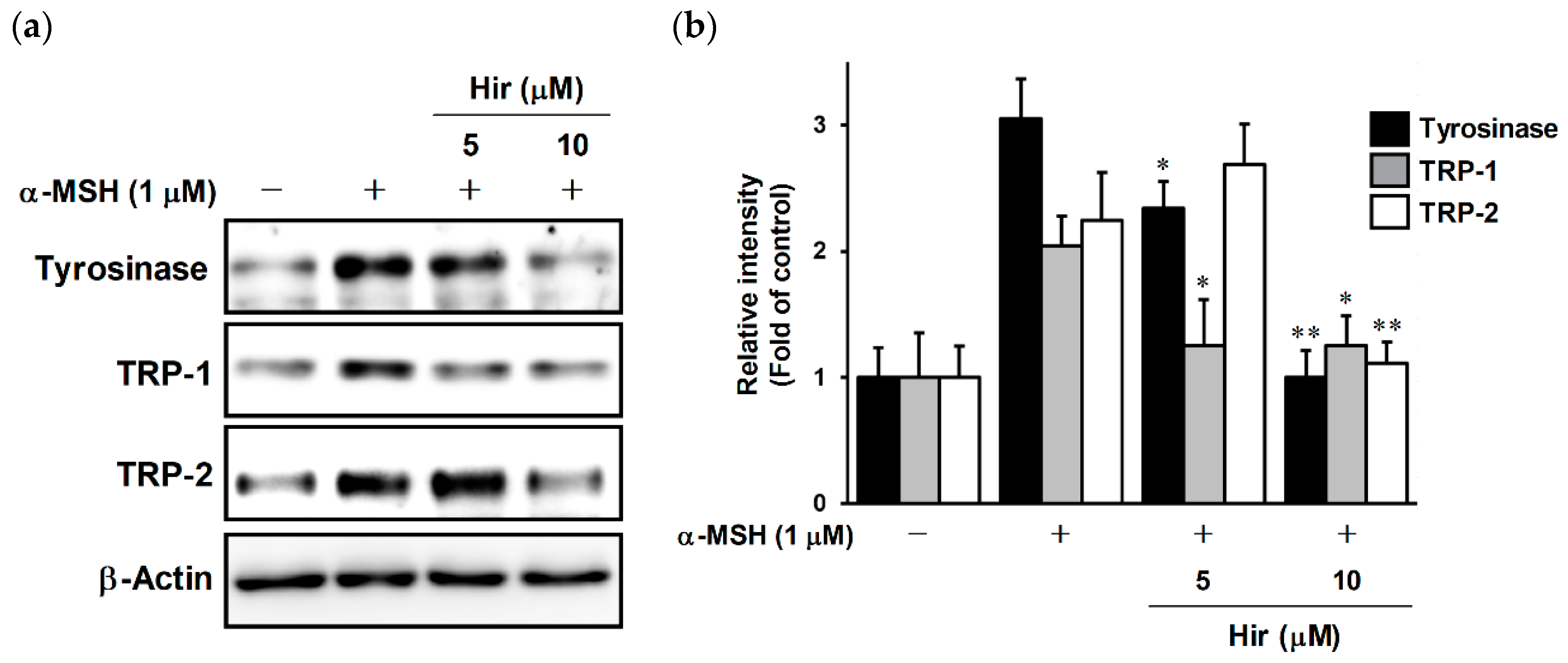
2.4. Effects of Hir on Expression of Tyrosinase, TRP-1, and TRP-2 in B16-F1 Cells
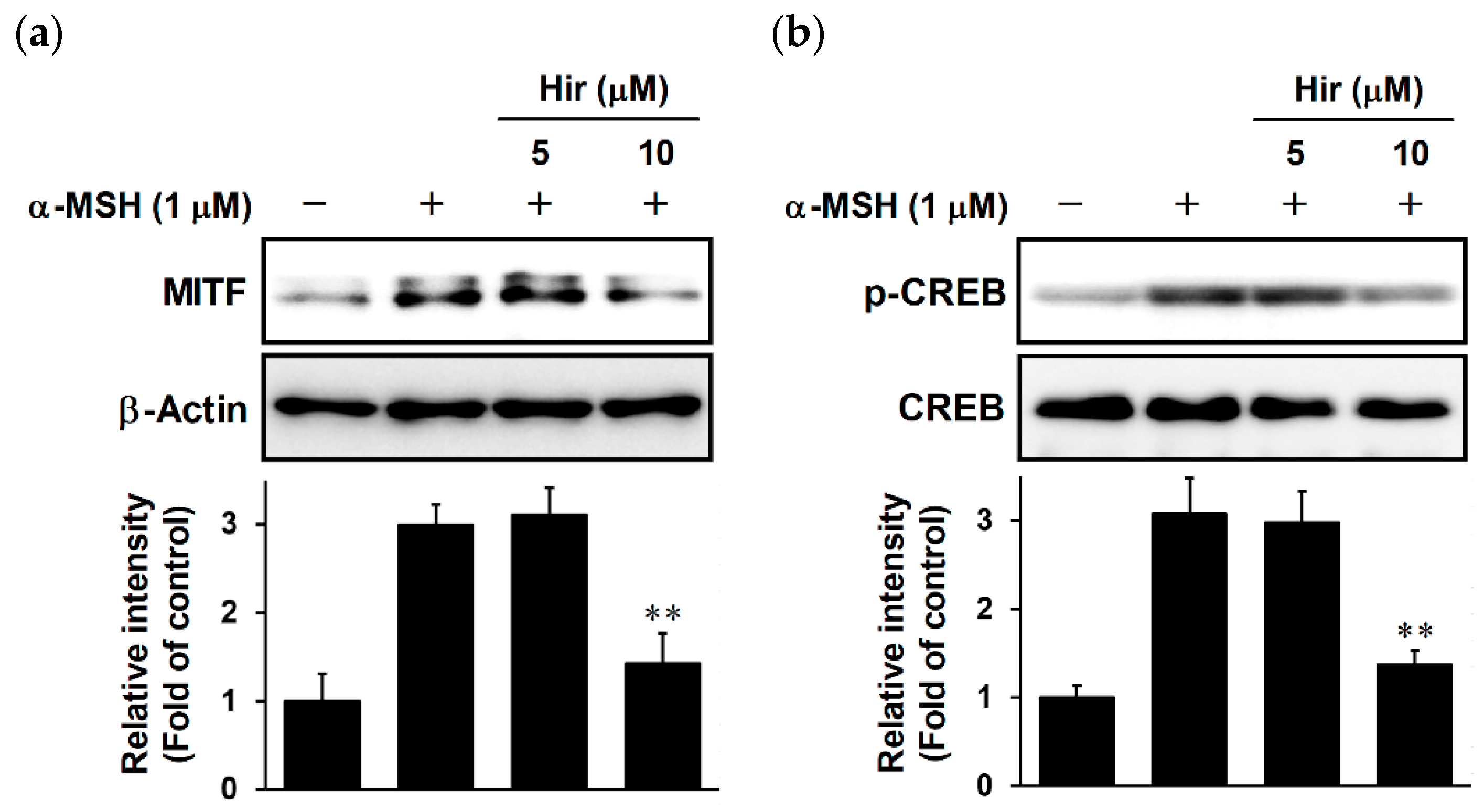
2.5. Effects of Hir on MITF Expression and CREB Phosphorylation in B16-F1 Cells
3. Materials and Methods
3.1. Materials
3.2. Plant Material and Isolation of Hir and Ore
3.3. Cell Culture and Treatment
3.4. Measurement of Melanin Content and Microscopy
3.5. Cell Viability Assay
3.6. Measurement of Cell-Free Mushroom Tyrosinase Activity
3.7. Assay of Intracellular Tyrosinase Activity
3.8. Western Blot Analysis
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lambert, M.W.; Maddukuri, S.; Karanfilian, K.M.; Elias, M.L.; Lambert, W.C. The physiology of melanin deposition in health and disease. Clin. Dermatol. 2019, 37, 402–417. [Google Scholar] [CrossRef] [PubMed]
- Le, L.; Sirés-Campos, J.; Raposo, G.; Delevoye, C.; Marks, M.S. Melanosome biogenesis in the pigmentation of mammalian skin. Integr. Comp. Biol. 2021, 61, 1517–1545. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y. Skin pigmentation abnormalities and their possible relationship with skin aging. Int. J. Mol. Sci. 2021, 22, 3727. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Jung, S.H. Recent development of signaling pathways inhibitors of melanogenesis. Cell Signal 2017, 40, 99–115. [Google Scholar] [CrossRef]
- Jimbow, K.; Obata, H.; Pathak, M.A.; Fitzpatrick, T.B. Mechanism of depigmentation by hydroquinone. J. Investig. Dermatol. 1974, 62, 436–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brtko, J. Biological functions of kojic acid and its derivatives in medicine, cosmetics, and food industry: Insights into health aspects. Arch. Pharm. 2022, e2200215. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, M.; Eslamifar, M.; Khezri, K. Kojic acid applications in cosmetic and pharmaceutical preparations. Biomed. Pharmacother. 2019, 110, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.X.; Xue, M.G.; Li, Z.; Ye, B.C.; Zhang, B. Recent progress on feasible strategies for arbutin production. Front. Bioeng. Biotechnol. 2022, 10, 914280. [Google Scholar] [CrossRef] [PubMed]
- Boo, Y.C. Arbutin as a skin depigmenting agent with antimelanogenic and antioxidant properties. Antioxidants 2021, 10, 1129. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Wichers, H.J.; Soler-Lopez, M.; Dijkstra, B.W. Structure and function of human tyrosinase and tyrosinase-related proteins. Chemistry 2018, 24, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Zhan, Q.; Du, S.; Ding, Y.; Fang, B.; Du, W.; Wu, Q.; Yu, H.; Li, L.; Huang, W. Catalysis-based specific detection and inhibition of tyrosinase and their application. J. Pharm. Anal. 2020, 10, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Urabe, K.; Winder, A.; Jemenez-Cervantes, C.; Imokawa, G.; Brewington, T.; Solano, F.; Garcia-Borron, J.C.; Hearing, V.J. Tyrosinase related protein 1 (TRP-1) functions as a DHICA oxidase in melanin biosynthesis. EMBO J. 1994, 13, 5818–5825. [Google Scholar] [CrossRef]
- Goding, C.R.; Arnheiter, H. MITF-the first 25 years. Genes Dev. 2019, 33, 983–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibahara, S.; Takeda, K.; Yasumoto, K.; Udono, T.; Watanabe, K.; Saito, H.; Takahashi, K. Microphthalmia-associated transcription factor (MITF): Multiplicity in structure, function, and regulation. J. Investig. Dermatol. Symp. Proc. 2001, 6, 99–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sati, S.C.; Sati, N.; Sati, O.P. Bioactive constituents and medicinal importance of genus Alnus. Pharmacogn. Rev. 2011, 5, 174–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, T.; Tokoroyama, T.; Kubota, T. Biarylheptanoids and other constituents from wood of Alnus japonica. Phytochemistry 1981, 20, 1097–1104. [Google Scholar] [CrossRef]
- Wada, H.; Tachibana, H.; Fuchino, H.; Tanaka, N. The new diarylheptanoid glycosides from Alnus japonica. Chem. Pharm. Bull. 1998, 46, 1054–1055. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.S.; Jang, E.R.; Kim, Y.J.; Myung, S.C.; Kim, W.; Lee, M.W. Diarylheptanoid hirsutenone enhances apoptotic effect of TRAIL on epithelial ovarian carcinoma cell lines via activation of death receptor and mitochondrial pathway. Investig. New Drugs 2012, 30, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Kuroyanagi, M.; Shimomae, M.; Nagashima, Y.; Muto, N.; Okuda, T.; Kawahara, N.; Nakane, T.; Sano, T. New diarylheptanoids from Alnus japonica and their antioxidative activity. Chem. Pharm. Bull. 2005, 53, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Tung, N.H.; Kim, S.K.; Ra, J.C.; Zhao, Y.Z.; Sohn, D.H.; Kim, Y.H. Antioxidative and hepatoprotective diarylheptanoids from the bark of Alnus japonica. Planta Med. 2010, 76, 626–629. [Google Scholar] [CrossRef] [Green Version]
- Tung, N.H.; Kwon, H.J.; Kim, J.H.; Ra, J.C.; Kim, J.A.; Kim, Y.H. An anti-influenza component of the bark of Alnus japonica. Arch. Pharm. Res. 2010, 33, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Tung, N.H.; Suzuki, M.; Uto, T.; Morinaga, O.; Kwofie, K.D.; Ammah, N.; Koram, K.A.; Aboagye, F.; Edoh, D.; Yamashita, T.; et al. Anti-trypanosomal activity of diarylheptanoids isolated from the bark of Alnus japonica. Am. J. Chin. Med. 2014, 42, 1245–1260. [Google Scholar] [CrossRef]
- Tung, N.H.; Sun, K.; Fan, J.Y.; Shoyama, Y.; Han, J.Y. Oregonin from the bark of Alnus japonica restrained ischemia-reperfusion-induced mesentery oxidative stress by inhibiting NADPH oxidase activation. Microcirculation 2014, 21, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.S.; Choi, S.E.; Kim, J.Y.; Kim, J.S.; Kim, E.J.; Park, K.H.; Lee, D.I.; Joo, S.S.; Lee, C.S.; Bang, H.; et al. Atopic dermatitis-like skin lesions reduced by topical application and intraperitoneal injection of hirsutenone in NC/Nga mice. Clin. Dev. Immunol. 2010, 2010, 618517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.Y.; Jeong, H.J.; Kim, J.H.; Kim, Y.M.; Park, S.J.; Kim, D.; Park, K.H.; Lee, W.S.; Ryu, Y.B. Diarylheptanoids from Alnus japonica inhibit papain-like protease of severe acute respiratory syndrome coronavirus. Biol. Pharm. Bull. 2012, 35, 2036–2042. [Google Scholar] [CrossRef] [Green Version]
- León-Gonzalez, A.J.; Acero, N.; Muñoz-Mingarro, D.; López-Lázaro, M.; Martín-Cordero, C. Cytotoxic activity of hirsutanone, a diarylheptanoid isolated from Alnus glutinosa leaves. Phytomedicine 2014, 21, 866–870. [Google Scholar] [CrossRef]
- Kang, S.; Kim, J.E.; Li, Y.; Jung, S.K.; Song, N.R.; Thimmegowda, N.R.; Kim, B.Y.; Lee, H.J.; Bode, A.M.; Dong, Z.; et al. Hirsutenone in Alnus extract inhibits akt activity and suppresses prostate cancer cell proliferation. Mol. Carcinog. 2015, 54, 1354–1362. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Tung, N.H.; Appiah-Opong, R.; Aning, A.; Morinaga, O.; Edoh, D.; Nyarko, A.K.; Shoyama, Y. Antiproliferative and pro-apoptotic activity of diarylheptanoids isolated from the bark of Alnus japonica in human leukemia cell lines. Am. J. Chin. Med. 2015, 43, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Uto, T.; Ohta, T.; Yamashita, A.; Fujii, S.; Shoyama, Y. Liquiritin and liquiritigenin induce melanogenesis via enhancement of p38 and PKA signaling pathways. Medicines 2019, 6, 68. [Google Scholar] [CrossRef] [Green Version]
- Uto, T.; Tung, N.H.; Ohta, T.; Shoyama, Y. (+)-Magnolin enhances melanogenesis in melanoma cells and three-dimensional human skin equivalent; involvement of PKA and p38 MAPK signaling pathways. Planta Med. 2022, in press. [Google Scholar] [CrossRef]
- Uto, T.; Ohta, T.; Katayama, K.; Shoyama, Y. Silibinin promotes melanogenesis through the PKA and p38 MAPK signaling pathways in melanoma cells. Biomed. Res. 2022, 43, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Ohta, T.; Nakayama, E.; Nakagawa, M.; Hatada, M.; Shoyama, Y. Bioassay-guided fractionation of clove buds extract identifies eugenol as potent melanogenic inducer in melanoma cells. J. Oleo Sci. 2022, in press. [Google Scholar]
- Song, H.; Hwang, Y.J.; Ha, J.W.; Boo, Y.C. Screening of an epigenetic drug library identifies 4-((hydroxyamino)carbonyl)-N-(2-hydroxyethyl)-N-phenyl-benzeneacetamide that reduces melanin synthesis by inhibiting tyrosinase activity independently of epigenetic mechanisms. Int. J. Mol. Sci. 2020, 21, 4589. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.B.; Lim, Y.Y. Antioxidant and tyrosinase inhibition activity of the fertile fronds and rhizomes of three different Drynaria species. BMC Res. Notes 2015, 8, 468. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uto, T.; Tung, N.H.; Shoyama, Y. Hirsutanone Isolated from the Bark of Alnus japonica Attenuates Melanogenesis via Dual Inhibition of Tyrosinase Activity and Expression of Melanogenic Proteins. Plants 2022, 11, 1875. https://doi.org/10.3390/plants11141875
Uto T, Tung NH, Shoyama Y. Hirsutanone Isolated from the Bark of Alnus japonica Attenuates Melanogenesis via Dual Inhibition of Tyrosinase Activity and Expression of Melanogenic Proteins. Plants. 2022; 11(14):1875. https://doi.org/10.3390/plants11141875
Chicago/Turabian StyleUto, Takuhiro, Nguyen Huu Tung, and Yukihiro Shoyama. 2022. "Hirsutanone Isolated from the Bark of Alnus japonica Attenuates Melanogenesis via Dual Inhibition of Tyrosinase Activity and Expression of Melanogenic Proteins" Plants 11, no. 14: 1875. https://doi.org/10.3390/plants11141875